Parasitism is always costly to the host
2021-04-15NanLyu,WeiLiang
DEAR EDITOR,
Recent studies have examined the cost of raising parasitic cuckoos and highlighted the importance of “no extra cost” in explaining the low levels of defense in hosts.To clarify the reasons for parasitization in typical hosts, we present a simple model to explore the immediate and future costs of parasitism in shaping the evolution of defense behavior in hosts.Our results suggest that any cost of parasitization is maladaptive to the host and learned egg recognition is always favored to overcome these costs.Furthermore, although facing a potential cost of mis-imprinting, learned nestling recognition may still evolve when there is a non-zero immediate cost from raising a parasitic nestling.Therefore, we contend that “no extra cost” does not provide sufficient evidence to explain the low levels of defense behavior in hostsper se.
Caring for an offspring is energetically expensive (Lukas,2013).Thus, theories predict that individuals will not expend energy if they do not gain fitness (Sheldon, 2002; Trivers,1972), as is the case for hosts of avian brood parasites (e.g.,cuckoos and cowbirds), who lay their eggs in the nests of other species to hatch and raise (Davies, 2000).The evolution of brood parasitism has been a long-lasting puzzle in biology,as rearing a parasite should always result in negative net fitness to the host (Davies, 2000; Soler, 2017).
Two recent published papers (Samaš et al., 2018, 2019a)explored the immediate and future costs of raising a common cuckoo (Cuculus canorus) on typical hosts.In the first case of the common redstart (Phoenicurus phoenicurus), a cavitynesting host, raising a parasitic cuckoo was not associated with overall higher physiological or physical costs to the hosts above natural levels imposed by efforts to rear their own progeny (Samaš et al., 2018).In the second case of the reed warbler (Acrocephalus scirpaceus) host, raising a cuckoo nestling did not entail any additional immediate or future costs for hosts above the natural costs incurred during the raising of their own offspring (Samaš et al., 2019a).These findings were suggested to explain, at least in part, the low levels of host defense seen in the above species (Samaš et al., 2018,2019a, 2019b; but see Yang et al., 2019) and the maintenance of long-term host-parasite systems (Krüger,2011).
However, host individuals that are successfully parasitized by cuckoos (Figure 1) may lose their reproductive output and fail to pass their genes onto the next generation.Thus, natural selection should always favor those individuals that possess greater defense against brood parasitism than others,because they can have more offspring and superior transmission of genes (Darwin, 1859; Dawkins, 1976).As such, parasitization should always be a huge cost for any host, which was not especially discussed in Samaš et al.(2018, 2019a).
In nature, to defend against cuckoo parasitism (i.e.,overcome the costs of being parasitized), various antiparasitism strategies have been reported in hosts, such as nest defense, egg recognition and rejection, and nestling discrimination (Feeney et al., 2014; Kilner & Langmore, 2011;Soler, 2014, 2017).Even in those host species that accept highly dissimilar cuckoo eggs, other mechanisms are applied to overcome the fitness costs of parasitism.For example,Cape bulbuls (Pycnonotus capensis) always accept the extremely dissimilar Jacobin cuckoo (Clamator jacobinus)eggs as they still have a relatively high probability of obtaining non-zero reproductive success, and nest desertion can increase predation and parasitism risks later in the season(Krüger, 2011).Sheehan & Reeve (2020) presented a general evolutionary game model to explore the evolution of recognition systems and found that recognition failure is more likely to arise when brood parasites do not kill host chicks within the same nests.Therefore, we contend that the no immediate or future extra cost concept proposed by Samaš et al.(2018, 2019a) does not provide sufficient evidence to explain the low level of host defense behavior or support their claims.
In this study, to clarify the reasons for the occurrence of parasitization in typical hosts, we present a simple model toexplore the immediate and future costs of parasitism in shaping the evolution of defense behavior in hosts (Lotem,1993).We highlight that any immediate cost of raising a cuckoo chick should promote the evolution of egg and nestling recognition in general.Furthermore, unlike the conclusion of Lotem (1993), we suggest that learned nestling recognition can evolve, although under relatively restricted conditions.

Figure 1 A common cuckoo nestling fed by its Oriental reed warbler host parent (Photo by Jian Yang)
Models
(1) Cost to parasitized host without defense strategies We established a relevant model following Lotem (1993).We assumed that each host individual will reproduce in two consecutive breeding seasons, and then will die (Figure 2A).The host individual is parasitized by a common cuckoo at a probability ofP.As either raising its own offspring or that of a cuckoo should be costly to the host (Dawkins, 1976; Trivers,1972), we assumed that if the host has not been parasitized(with a probability of 1-P), it pays a cost ofcto raise its own offspring and then acquires reproductive successx(x>0) on average.In contrast, if the host has been parasitized, its reproductive success will be reduced to zero, but it will still pay a cost ofc" to rear the cuckoo chick (representing an“immediate cost”, as given by Samaš et al., 2019a).
In the second breeding season, those host individuals that have not been parasitized will pay an additional cost of Δc(representing a “future cost”, as given by Samaš et al.,2019a), while those host individuals that have been parasitized will pay an additional cost of Δc".The payoff for a host individual can therefore be calculated as follows:

Figure 2 Schematics of host fitness models

which reduces to:

Any positive values ofc" and Δc" would reduce the fitness of the host individual (whenP>0), i.e., any cost of being parasitized is maladaptive to the host.
(2) Cost to parasitized host with defense strategies
We next included the host defense strategies of egg and nestling recognition into the model, respectively.In the firstcase of egg recognition (Figure 2B), the host learns to recognize and reject the cuckoo egg in the second breeding season if it has not been parasitized during the first breeding season, i.e., it will gain a benefit ofb(Lotem, 1993) and no longer needs to pay the cost ofc" as the cuckoo egg will have been rejected.In contrast, if the host has been parasitized during the first breeding season, egg recognition does not develop as both the cuckoo egg and host’s own eggs co-occur in the nest.The host will therefore obtain the same fitness in the second breeding season as in the first model (see Figure 2A).The payoff for the host individual thus becomes:

Learned egg recognition can evolve whenfe>fis satisfied,which reduces to
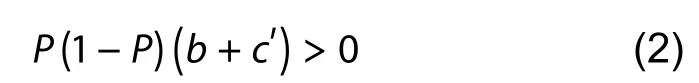
Therefore, even if there is no benefit gained by rejection(i.e.,b=0), learned egg recognition is still favored as long as raising a cuckoo chick is costly (i.e.,c" >0).
In the case of learned nestling recognition (Figure 2C), the host may develop a mistaken imprint of the cuckoo nestling as its own (Lotem, 1993), and, if so, will reject its own nestlings in the future.The payoff for the host individual thus becomes:

Nestling recognition can evolve whenfn>fis satisfied, which reduces to:

This shows that whenb>x-c"-cis satisfied, learned nestling recognition is favored (whenP>0).Additionally, as the benefit of rejection (b) should be lower than the net fitness value of an unparasitized individual (i.e.,x-c) in general, we therefore obtain the conditionx-c"-c<b<x-cfor the evolution of learned nestling recognition.Theoretically, any positive value for the immediate cost of raising a cuckoo chick (i.e.,c">0) is essential for driving the evolution of learned nestling recognition.
Samaš et al.(2018, 2019a) suggested thatc" is equal tocand Δc" is equal to Δc.However, positing that this can promote the evolution of parasitism and explain the low levels of host counter-defenses is unfounded as an improved defense capability should always be more beneficial for a host to reduce the costs ofc" and Δc" (see equation 1), e.g., through learned egg or nestling recognition.Therefore, to unravel the adaptive evolution of recognition failures in hosts, other apparent benefits are still required to overcome the fitness costs of parasitism (Davies, 2000).
In addition, the results given by Samaš et al.(2018, 2019a)are insufficient to support the conclusion that rearing a cuckoo chick would not bring immediate or future extra costs.
Firstly, most of the data in Samaš’s study were collected by transferring cuckoo hatchlings from deserted or multiparasitized great reed warbler (Acrocephalus arundinaceus)nests (natural parasitism rate >50%) into reed warbler nests(rate of 5% to 20%) (Samaš et al., 2019a).This implies that changes in body mass, heterophil/lymphocyte ratio (i.e.,stress-level indicator), and feeding frequencies represent responses of those reed warbler individuals that have not been parasitized.The authors assumed that there would be no differences in adult body mass on nest hatching day between individuals naturally parasitized by cuckoos or those not; however, they cannot exclude the possibility that parasitized reed warblers may differ from those that have not been parasitized in condition or even behavioral patterns.
Secondly, future costs were assessed by return rates from only 16 re-trapped individuals the following year (Samaš et al.,2019a).It remains unclear whether the non-parasitized individuals would have a longer lifespan and more reproductive success than those that have been parasitized or would have a lower rate of parasitization and produce more offspring in the future.
Additionally, as the authors discuss in their paper, host parents need to provide post-fledging care (e.g., 17 days for cuckoo chicks), which may further aggregate the host burden.
Therefore, we suggest that the existence of immediate or future extra costs remains to be explored, and the claim that“no extra cost” helps to explain the evolution and maintenance of host defense in avian brood parasitism does not hold upon further examination and requires stronger evidence (also see Yang et al., 2019).
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS’ CONTRIBUTIONS
W.L.designed the study; N.L.developed the model and wrote the draft manuscript; W.L.revised and improved the manuscript.All authors read and approved the final version of the manuscript.
ACKNOWLEDGEMENTS
We would like to thank the two anonymous reviewers for their constructive comments on this manuscript.
杂志排行

Zoological Research的其它文章
- A new species of Kurixalus (Anura, Rhacophoridae)from Guizhou, China
- Comparative transcriptomics highlights convergent evolution of energy metabolic pathways in group-living spiders
- Long-term trends in a forest ungulate community: park establishment increases numbers, but poaching is a constant threat
- Molecular phylogeny of the genus Muntiacus with special emphasis on the phylogenetic position of Muntiacus gongshanensis
- Flexible breeding performance under unstable climatic conditions in a tropical passerine in Southwest China
- Yunnanilus chuanheensis, a new loach species(Cypriniformes: Nemacheilidae) from the upper Lixianjiang River in Yunnan, China