黄河三角洲湿地不同景观类型影响下植被与土壤因子的空间分布特征
2021-04-13路景钫石洪华孙辉李捷黄建涛刘译蔓
路景钫, 石洪华, 孙辉, 李捷, 黄建涛, 刘译蔓
黄河三角洲湿地不同景观类型影响下植被与土壤因子的空间分布特征
路景钫1, 3, 石洪华2, 3, 孙辉4, 李捷1,*, 黄建涛1, 刘译蔓1
1. 青岛理工大学环境与市政工程学院, 山东, 青岛 266033 2. 青岛海洋科学与技术国家实验室海洋地质过程与环境功能实验室, 山东, 青岛 266061 3. 自然资源部第一海洋研究所, 山东, 青岛 266061 4. 山东省调水工程运行维护中心青岛分中心, 山东, 青岛 266300
以黄河三角洲湿地为研究区, 结合遥感影像和实地调查数据, 构建湿地景观类型和主要影响因子的空间分布格局, 解析不同景观类型下的植被及土壤因子的空间分布特征以及耦合关系。结果表明: 农田、湿地植被区是研究区内面积最大的景观类型, 分别占有46.87%、20.6%, 而植被覆盖贡献率59.07%、37.62%, 生物量贡献率59.08%、36.04%, 是湿地蓝碳资源的重要补充。另外, 黄河三角洲湿地年均有效磷(AP)以Ⅳ级土壤为主, 土壤肥力标准处于缺乏状态, 景观类型中农田区年均AP最高, 湿地植被区最低; 年均有效钾(AK)以Ⅲ级土壤为主, 处于中等状态; 光滩、盐田区年均AK高、湿地植被区最低。最后,相关性分析显示: 春秋两季, 有效钾与Shannon-wiener指数、植被覆盖度呈显著负相关, 成为植物生长、凋零过程中主控因子。
黄河三角洲; 湿地; 景观类型; 土壤因子; 植被
0 前言
滨海湿地因其独特且重要的生态地位, 一直是环境生态学研究的热点[1-3], 它具有多种生态功能, 包括改善当地气候环境、生物多样性的维持、养护水资源等。通过遥感技术既能及时、综合的对湿地景观格局变化进行研究[4-6], 又可以为植被生物量反演、生态健康评价以及环境污染遥感检测等研究提供方便[7-8]。滨海湿地研究目前主要涉及景观格局和地貌演变、海岸侵蚀、生物群落结构、滨海湿地退化和恢复等方面[9]。
黄河三角洲, 是我国暖温带最完整、面积最大的滨海湿地[10], 其生态系统具有丰富完整的湿地植被, 而土壤理化性质与植物生长过程密切相关[11-15], 土壤中的来源相同的各种元素之间又会相互影响[19-20], 同时湿地土壤性质与地表植被又受景观类型演变的影响[16-18]。近年来, 众多学者围绕黄河三角洲生态系统同样开展了一系列卓有成效的工作, 其中不乏生态系统碳过程、土壤碳含量、NPP 空间分布等相关研究[21-24], 但目前缺乏小尺度景观下的植被因子与土壤因子耦合关系研究[25]。本文通过遥感影像解译得到2017年黄河三角洲滨海湿地景观格局, 结合现场调查的91组实地数据分析了土壤因子、植被因子间的耦合关系, 并模拟两种因子受到区域、季节、土地利用类型的不同呈现空间分布规律。不仅可以对连续时间尺度上的黄河三角洲滨海湿地变化后生态系统的在空间变化上进行分析, 也能对未来黄河三角洲滨海湿地资源利用的变化趋势做出更具时效性的预测。
现代黄河三角洲(118°32′—119°15′E, 37°31′—38°91′N)是指以垦利县鱼洼为顶点、以挑河口和宋春荣沟口为端点构成的区域。本文结合地理完整性和实际情况, 将研究区南侧端点南延至垦利县滨海大道南端红光渔港处。行政区范围以东营市垦利区、河口区和利津县等多个行政区围成的扇形淤积面积(图1), 并全部位于东营市境内, 分别属于河口区(51%)、垦利区(40%)和利津县(9%)。黄河三角洲地处北温带, 属暖温带大陆性季风气候, 年均气温为12.3—12.8℃, 年降水量为530—630 mm。多集中于夏季, 常伴有旱、涝、暴雨等自然灾害发生。植被结构方面, 受气候、降水、含盐量和地貌类型及人类活动多重因素的影响, 类型少, 以草甸景观为主。自然植被以盐生草甸为主, 典型植物群落优势种有芦苇()、柽柳()、盐地碱蓬()等。
1 材料与方法
1.1 分区布点及采样
根据孙志高, 朱博勤, 叶思源[26-28]等人对黄河三角洲的区域划分的研究成果以及研究区内生态保护区开发、管理现状, 将滨海湿地划分为保护区与非保护区两大类, 其中保护区包括一千二管理区(A)和大汶流管理区(C); 非保护区包括油田区(B)、黄河农用区(D)与黄河故道农用区(E)[26-28], 进一步根据所在区的主要景观类型面积占有情况划分为植被区、植被农田区、光滩盐田区、农田1区以及农田2区。具体站位布点信息如表1。
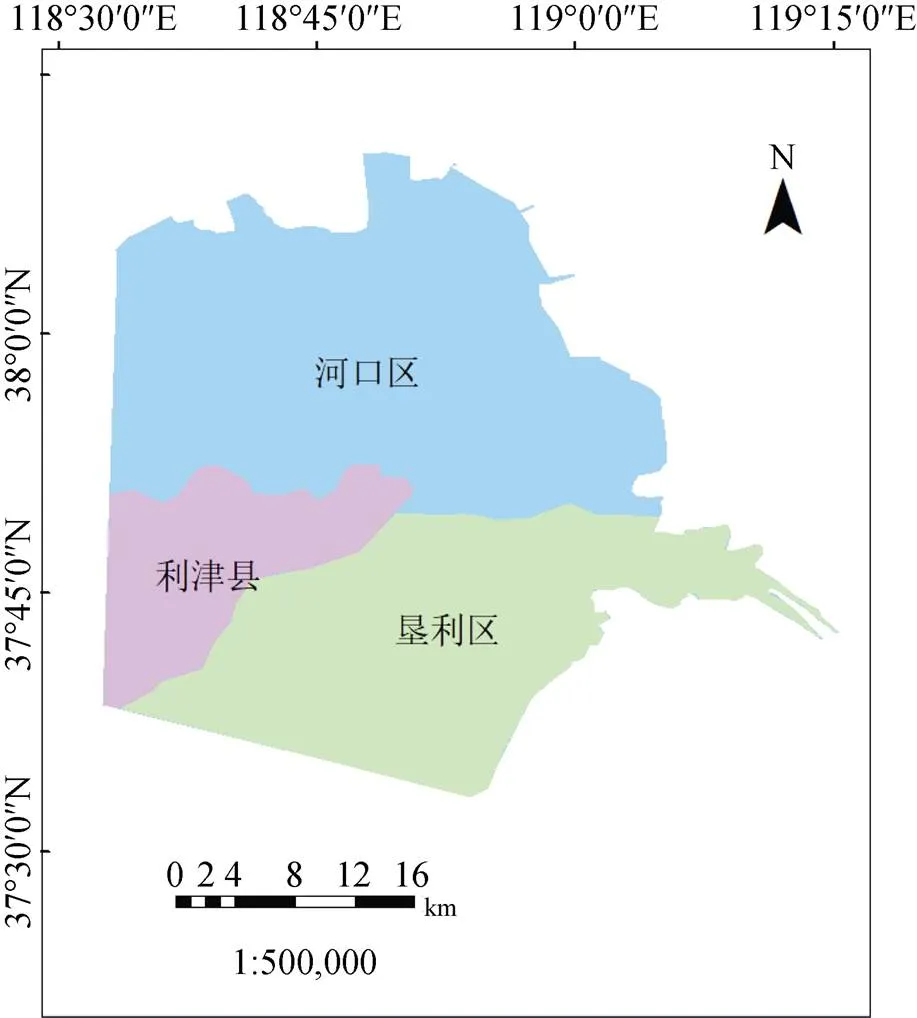
图1 研究区范围
Figure 1 Scope of study area
选取的91个野外调查站位采用网格布点法 (图2)。样地设1 mÍ1 m样方, 当场鉴定植物种类、覆盖率并取样方内全部植物地上部分带回实验室, 土壤样品均为表层土壤(0—20 cm深度)。
1.2 数据检测
测生物量数据, 取内所有植物地上部分, 在100℃下恒温烘干至恒重并称量; 测土壤数据需前处理, 在实验室风干, 取出石块和植物根系等杂物, 研磨过筛。土壤pH值测定采用电位法; 土壤全氮(TN)测定采用开氏法; 土壤有效磷(AP)测定采用碳酸氢钠浸提-钼锑抗分光光度法; 土壤有效钾(AK)测定采用醋酸铵提取, 火燃光度法[28]。以上所有指标均测定3个平行, 取平均值。
1.3 遥感影像处理
在地理空间数据云网站(http://www.gscloud.cn/)选择Landsat 8 OLI_TIRS 卫星数字产品。并选取2017年10月黄河三角洲滨海湿地区域(经纬度范围: 118.1055°E—120.1444°E、36.3972°N—38.5361°N)的Landsat 8遥感影像, 行列号121/34, 云量4.62%。
在ENVI 5.0和ArcGIS10.0中对遥感影像进行图像配准以及波段融和, 获得非标准假彩色图像。通过目视解译法并建立景观分类体系(表2), 将研究区地表覆盖类型划分为植被、农田、水域、盐田、养殖池、裸地、建筑类用地、光滩8类, 通过得到2017年黄河三角洲滨海湿地景观类型分布图(图3)。
1.4 植被因子、土壤肥力划分标准
按照全国养分含量等级标准将黄河三角洲滨海湿地TN、AK、AP含量划分为丰富(I级)、较丰富(II级)、中等(III级)、较缺乏(IV级)、缺乏(V级)和极缺乏(VI级)6个等级(表3)。
参考任继周、李博[29-30]等人对植被、草地退化程度划分的方法以及研究区内的植物物种组成、地上生物量与盖度、土壤状况, 依据植被盖度划分成未退化、轻度退化、中度退化、重度退化、极度退化(盖度分别为>90%、70%—90%、50%—70%、30%—50%、<30%)。参考池源[31][32]的研究成果, 将研究区划分为高生产力区、中高生产力区、中生产力区、中低生产力区以及低生产力区(生物量分别为>180g·m–2、120g·m–2—180g·m–2、80g·m–2—120g·m–2、50g·m–2—80g·m–2、<50g·m–2)。
1.5 数据分析
用ORIGIN 2018版进行基础数据和图型的整理; 运用AICGIS中的Kriging插值法表现不同土壤因子在不同季节、不同地域的空间梯度特征, 利用重分类法计算不同肥力指数类型的面积情况; 运用SPSS10中Spearman相关性分析对黄河三角洲地区的土壤因子之间进行相关性分析。
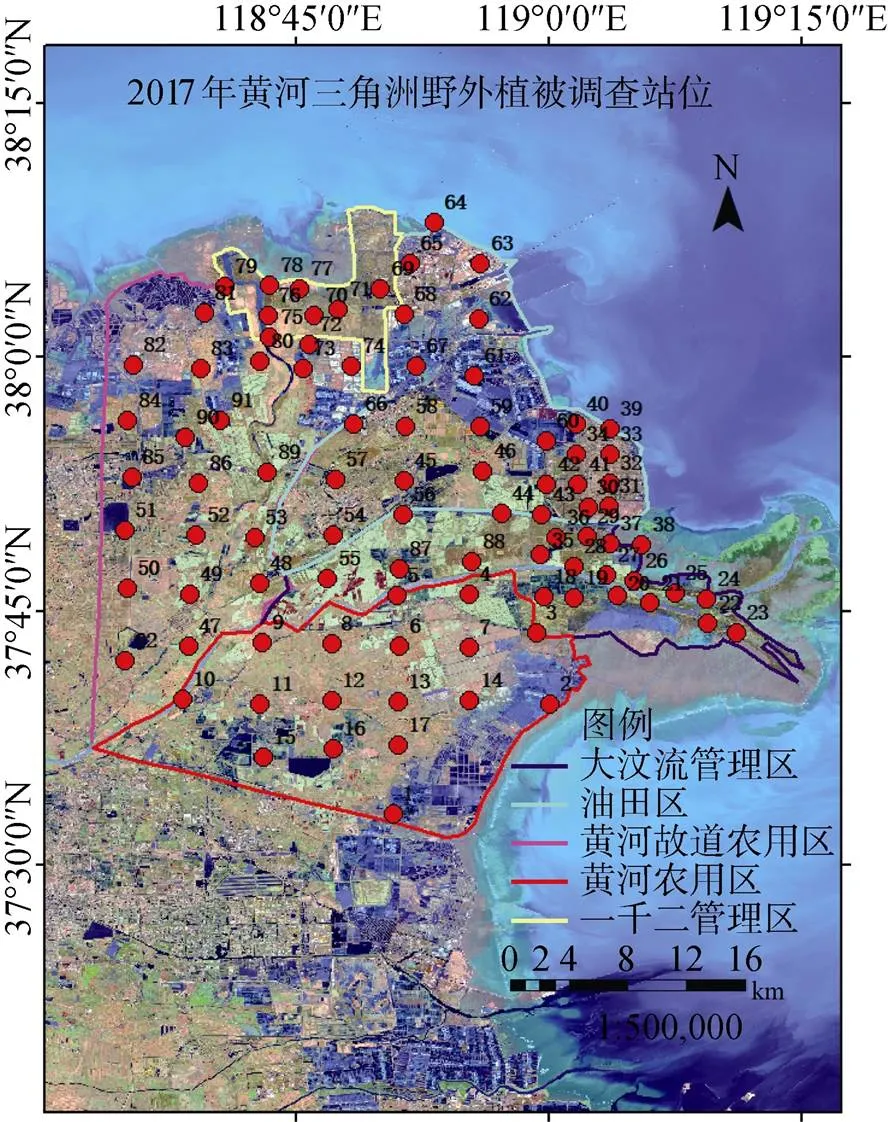
图2 黄河三角洲滨海湿地野外调查站位
Figure 2 Field survey stations of YRDW
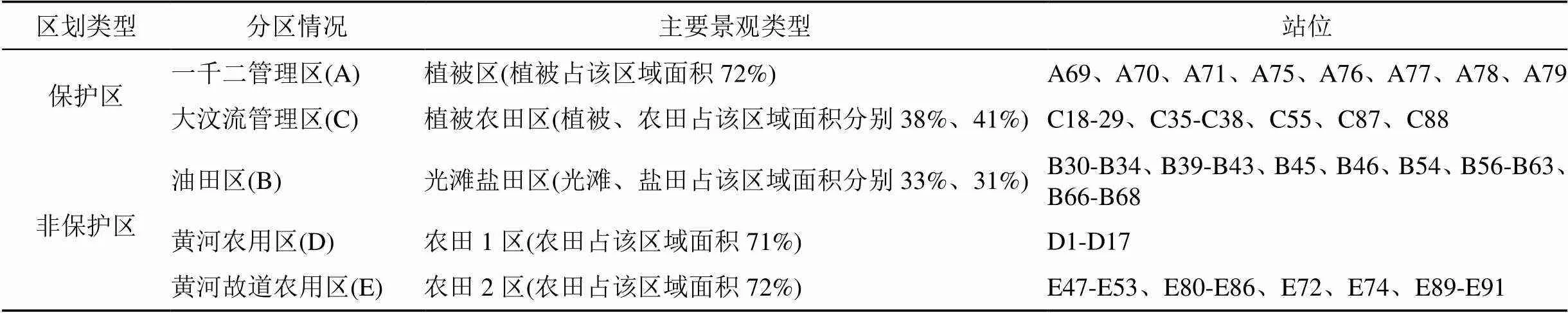
表1 分区站位布点信息

表2 黄河三角洲湿地景观分类体系

表3 土壤肥力指标等级划分
2 结果与分析
2.1 黄河三角洲湿地景观类型
截止到2017年10月, 黄河三角洲滨海湿地研究区面积共计2496.2 km2, 其中农田以1198.53 km2成为第一大景观类型, 植被区次之为528.53 km2, 两种景观类型面积占比分别为46.87%、20.6% (表4)。
2.2 不同景观类型下植被因子空间分布分析
研究区的植被覆盖度与生物量不同季节表现出极大的空间相似度, 而不同季节表现出了明显的空间分异性(图4—图5)。夏、秋两季的整体着色度明显要高, 主要体现在内陆的黄河农用区南部以及黄河故道农用区中部, 其生物量划为高生产力区、植被覆盖度均划为轻度退化区。
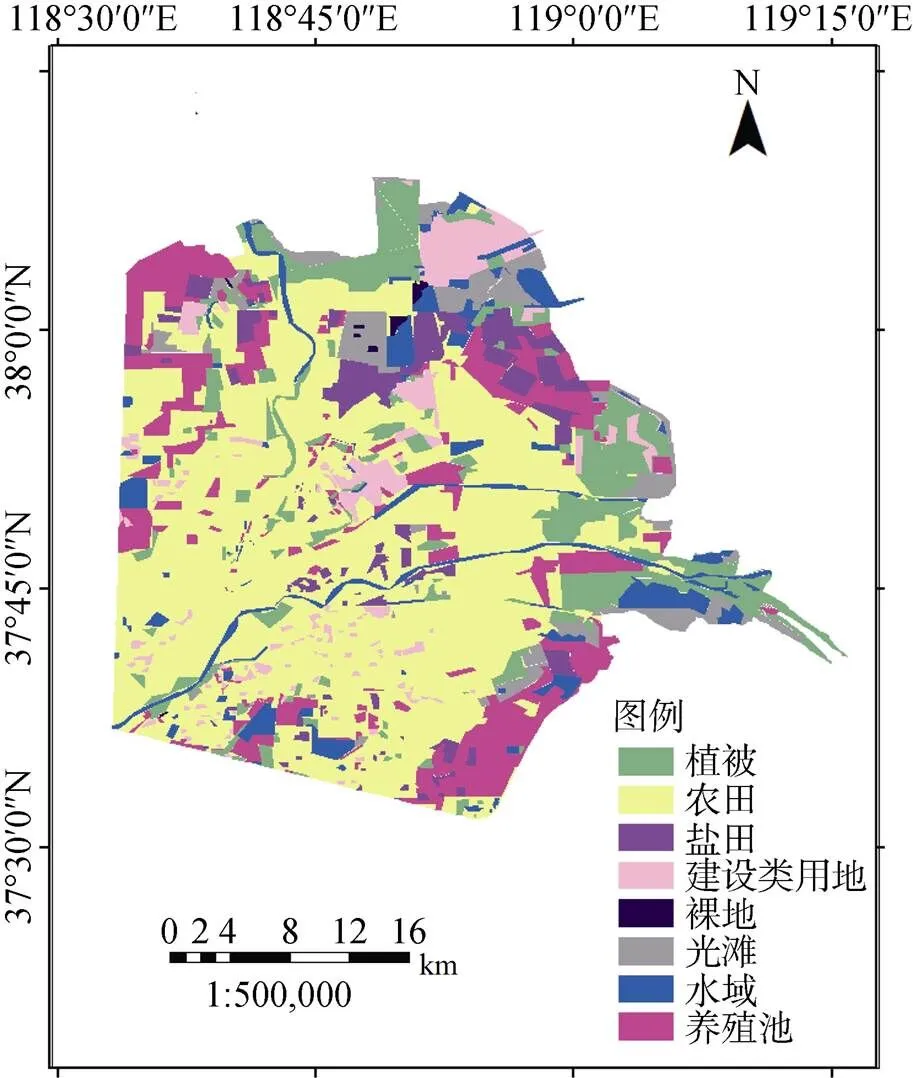
图3 2017年黄河三角洲滨海湿地景观类型分布
Figure 3 Landscape types distribution of the YRDW in 2017

表4 不同景观类型的面积统计情况
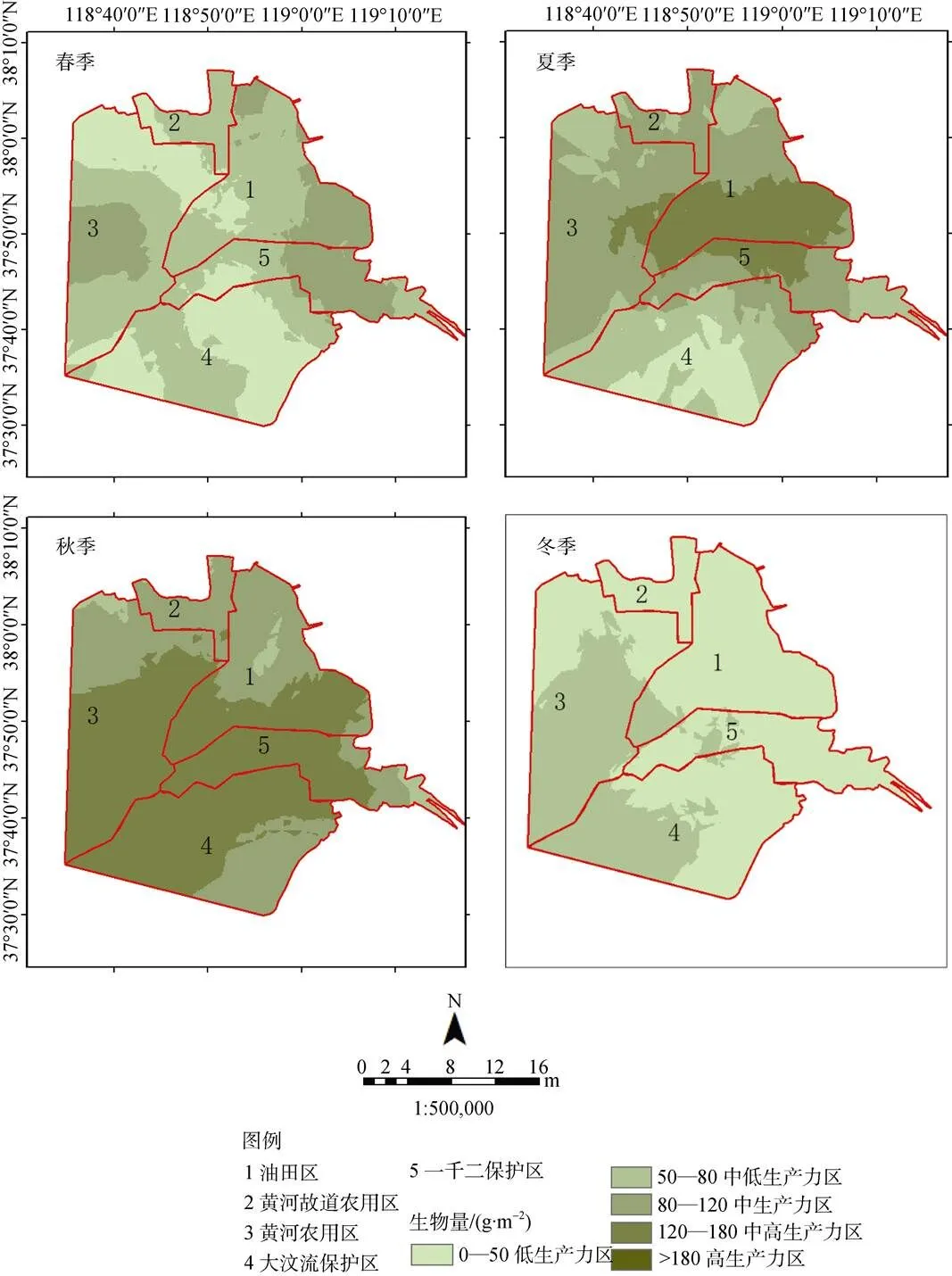
图4 2017年4个季度黄河三角洲生物量空间梯度变化
Figure 4 Spatial gradient changes of biomass in the fourth quarter
不同景观类型同样表现出了明显的空间分异性, 农田相比其他景观类型表现出更大的波动性, 其下降和增长都十分明显, 秋季生产力184.6 g·m–2, 植被覆盖率90.9%, 达到未退化、高生产力区; 而春季刚播种, 生物量34.5 g·m–2, 植被覆盖率24.3%, 为退化、低生产力区。湿地植被、光滩四季植被覆盖度以及生物量波动小, 年均值达到轻度退化、中生产力区; 而光滩区植被覆盖度、生物量低, 全年为重度退化、低生产力区(表5)。
根据表5统计结果, 计算四类景观的年平均生物量、植被覆盖度, 得到植被区、农田植被覆盖度贡献率37.62%、59.07%、生物量贡献率36.04%、59.08%, 都远高于盐田以及光滩, 若用植被覆盖度和生物量两个指标来反映不同景观类型下植被的生长状态, 则植被区优于农田区优于盐田区, 光滩区最差(图6)。
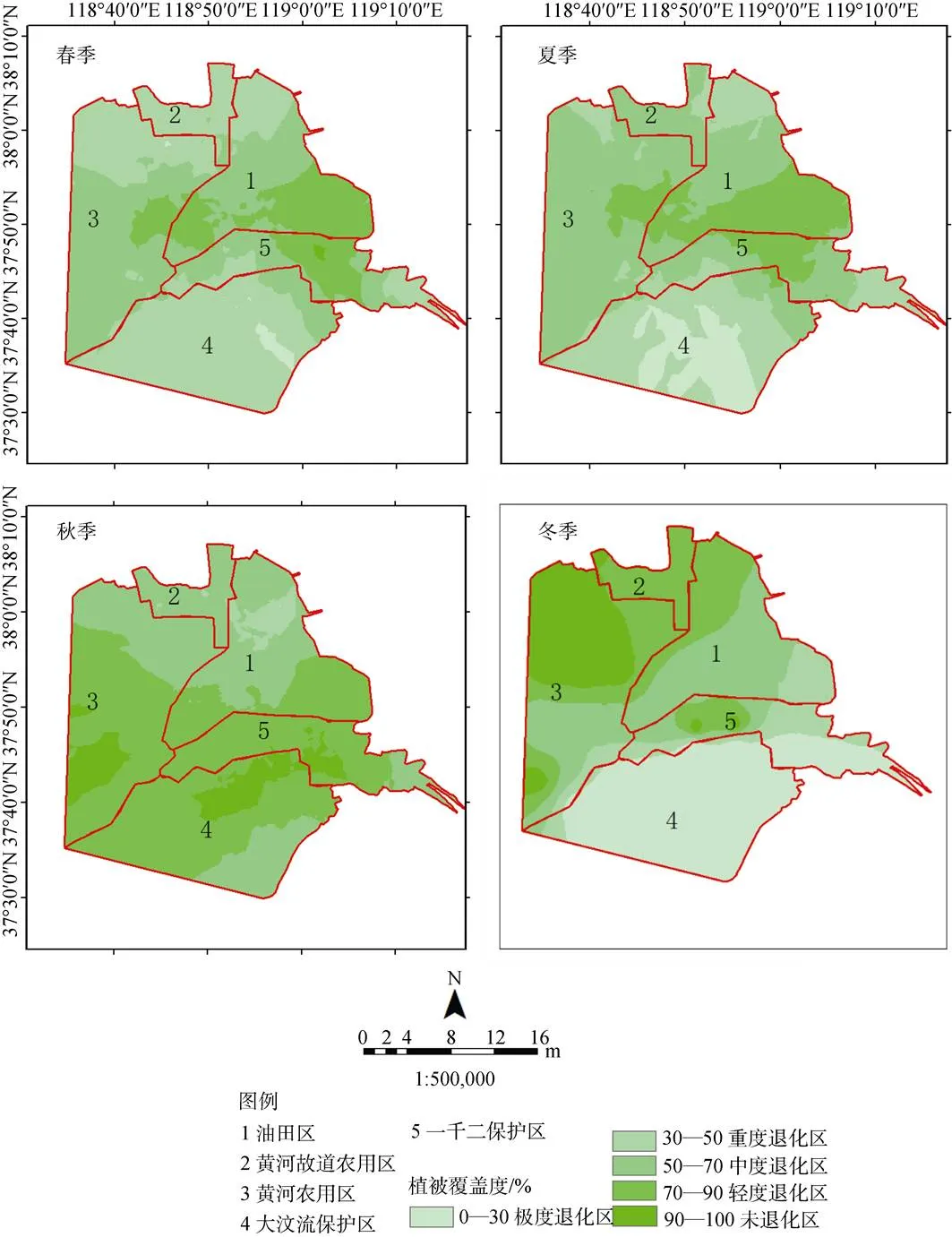
图5 2017年4个季度黄河三角洲植被覆盖度空间梯度变化
Figure 5 Spatial gradient change of vegetation coverage in YRWD in the four quarters
2.3 不同景观类型下土壤因子季节空间分布特征
2.3.1 土壤含水量(SW)
研究区不同季节的SW呈现出相同的空间梯度变化规律, 整体而言中部区域的SW高, 四周低; 东部沿海高, 内陆低。从不同景观类型分析: 光滩盐田区和植被区SW差别更为明显。8月(秋季)、11月(冬季)相对于2月(春季)、5月(夏季)在整体颜色波动明显, 有明显的下降, 农田区SW季节波度小(图7)。
不同景观类型同样表现出了明显的空间分异性: 黄河三角洲西部的湿地植被区呈现的深蓝色均值在32%左右, 其次是光滩盐田区均值在24.6%, 上述两种景观类型含水量最高; 农田2区及农田1区呈绿色SW低, 均值在22%以下。
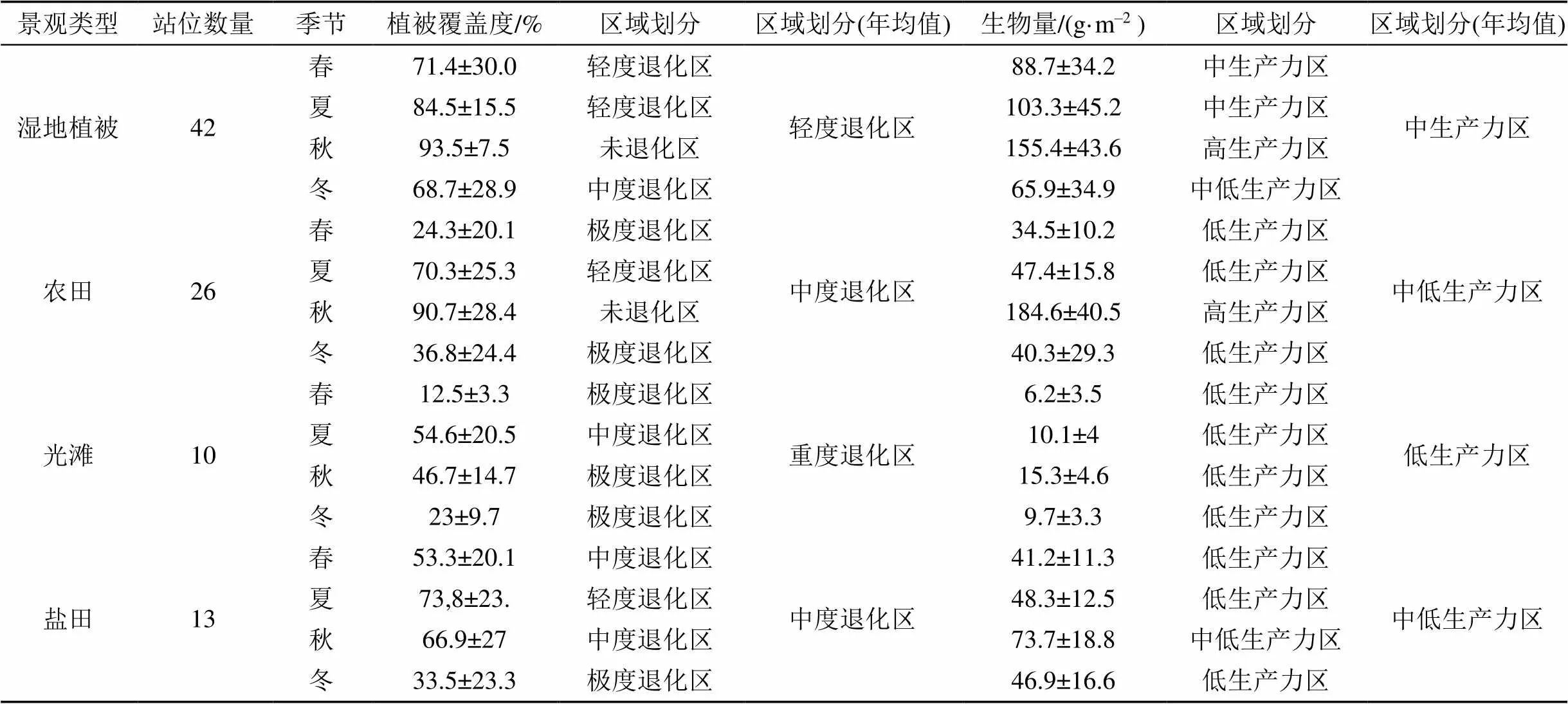
表5 不同景观类型四个季度生物量、植被覆盖度分区情况
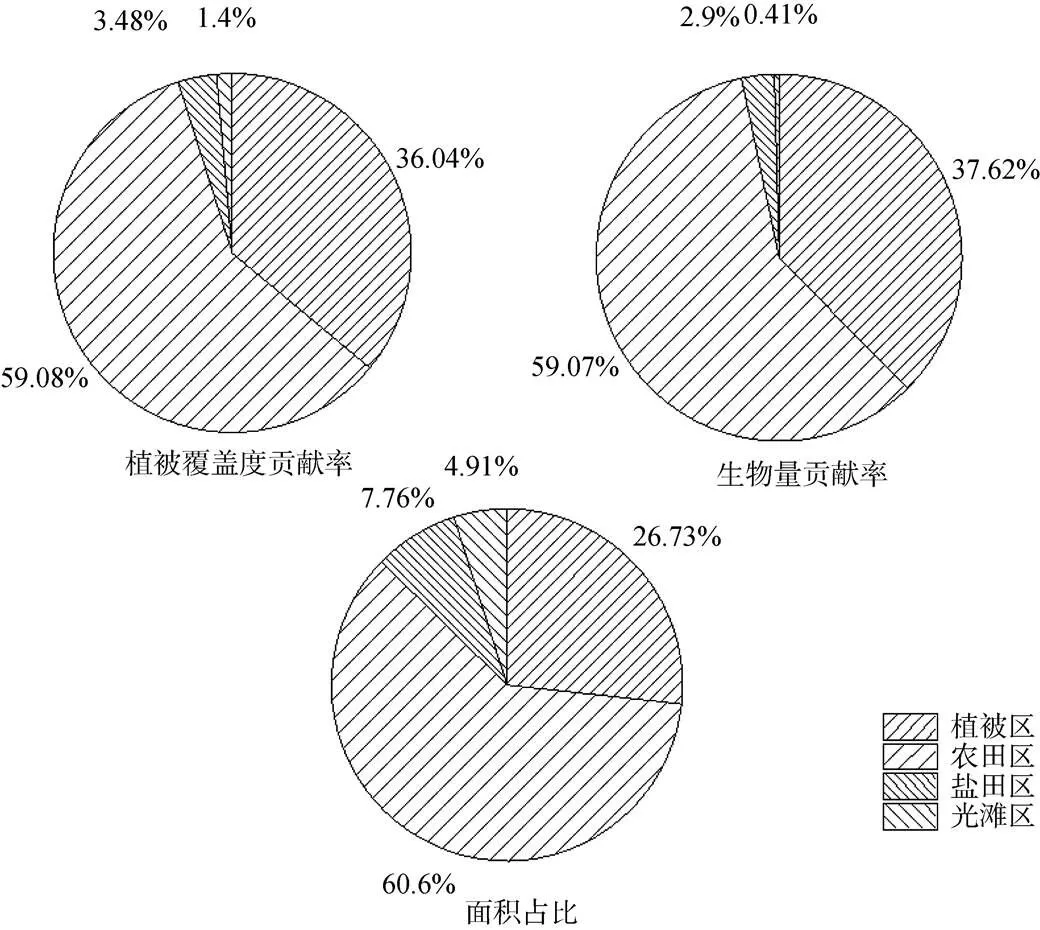
图6 不同景观类型下植被因子的贡献率
Figure 6 Contribution rate of vegetation factors under different landscape types
2.3.2 土壤含盐量(SA)
研究区不同季节的SA呈现出相同的空间梯度变化规律, SA高的区域主要在沿海带, 呈现三角之势即西北角、东北角和东南角多呈现蓝色(三个区域主要的景观类型为盐田)并向中心递减。从不同景观类型分析: 沿海的光滩盐田区、植被区SA均值最高(>8 mg·kg–1), 而内陆的农田1区、2区SA均值在5 mg·kg–1以下, 远低于其他景观类型(图8)。
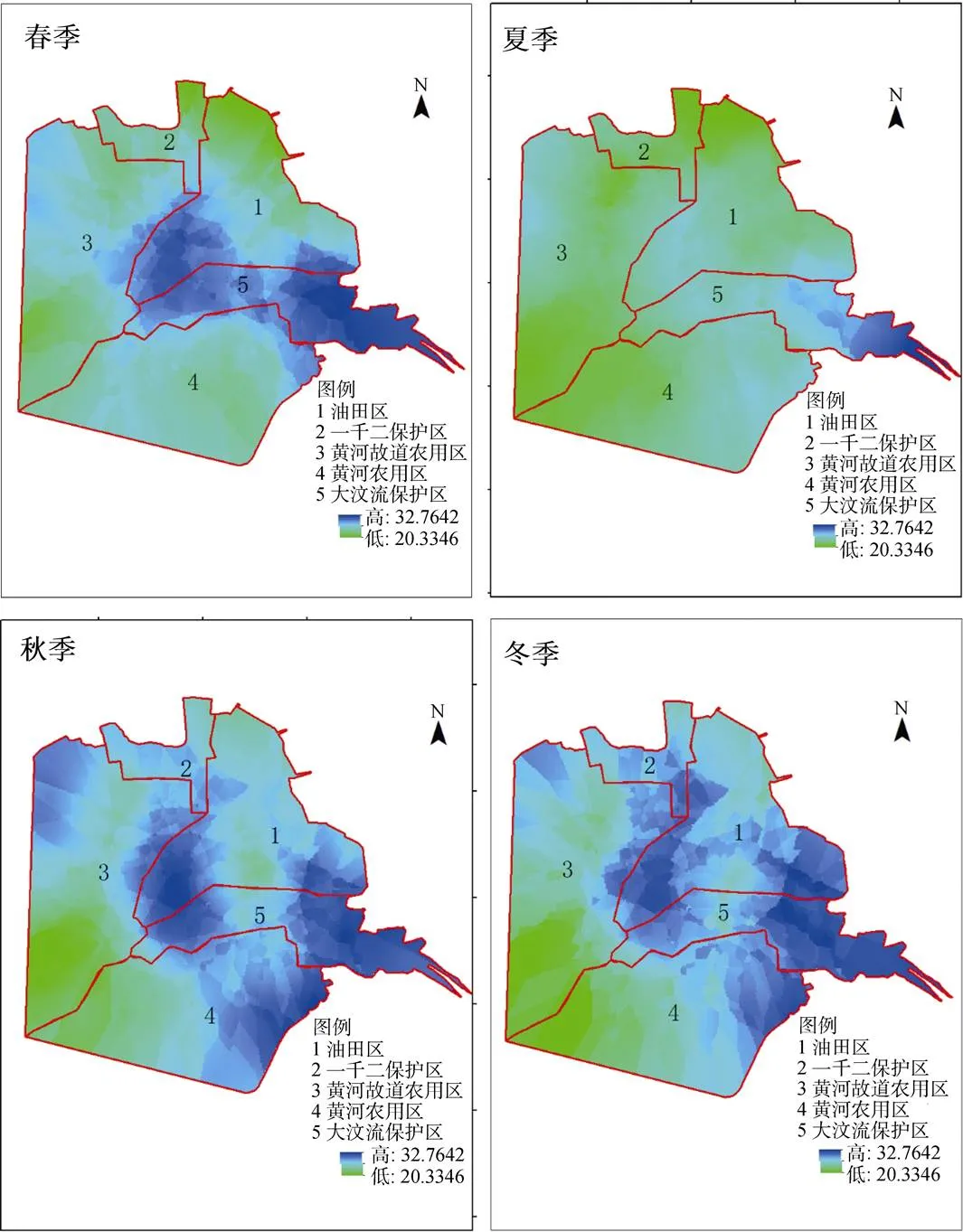
图7 2017年黄河三角洲湿地4个季度SW空间梯度变化
Figure 7 Spatial gradient changes of SW in four quarters
2.3.3 土壤有效磷(AP)
研究区不同季节的AP呈现出不规律空间梯度分布, 区域AP相关度不高。春季呈现沿海地区低于内陆地区的规律, 只有沿海的光滩盐田区区域显示深灰色, 达到IV级标准, AP的含量最高; 夏季研究区整体呈现浅灰色, 属于VI级标准的极缺乏状态; 秋季农田1区、2区80%面积显示为浅灰色(V级标准及以下), AP的含量最十分低。冬季AP含量整体要高, 农田1区要整体达于III标准, 植被区、农田2区达到IV级标准, 整体呈现西高东低(图9)。
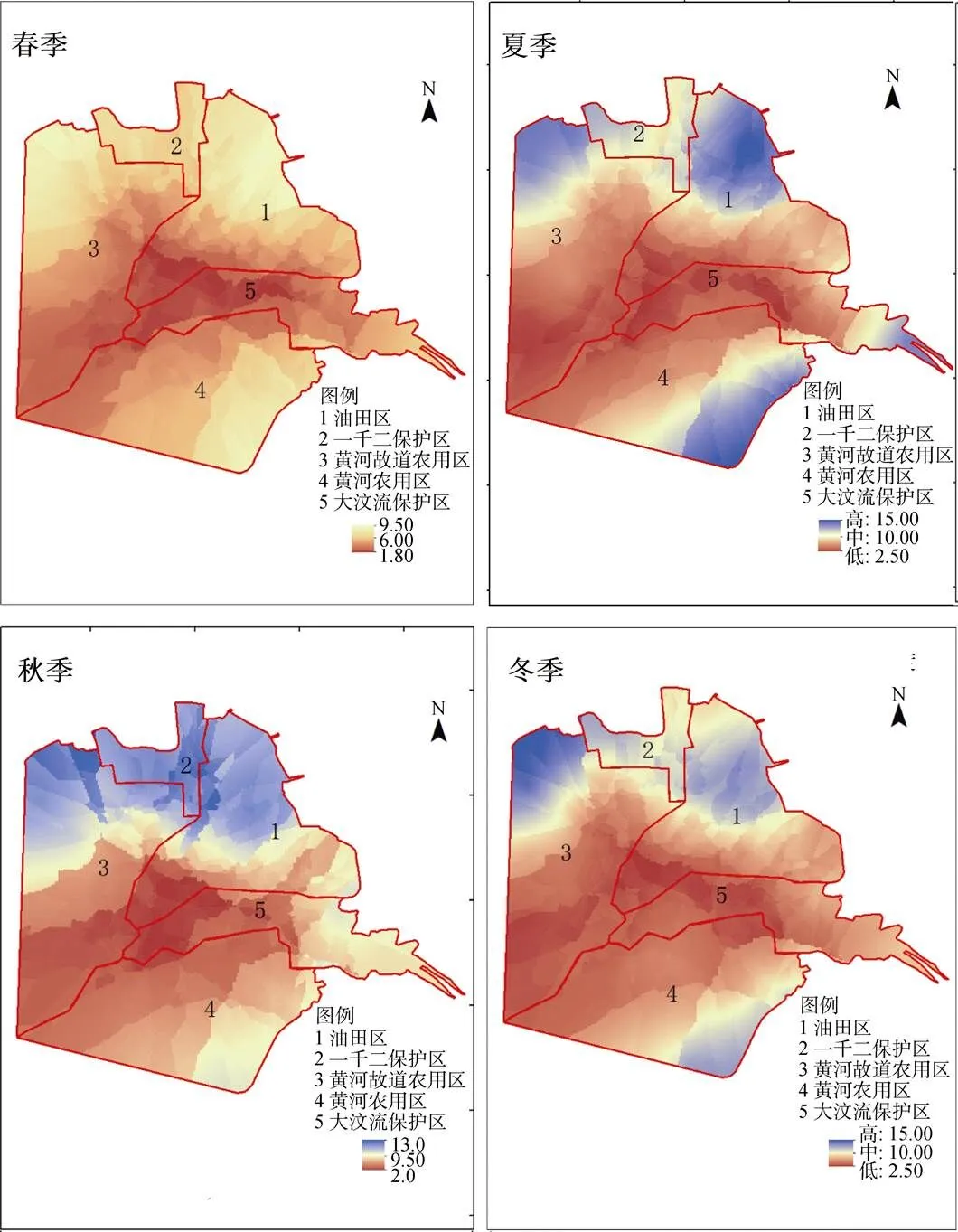
图8 2017年4个季节黄河三角洲SA空间梯度变化
Figure 8 Spatial gradient change of SA in four quarters.
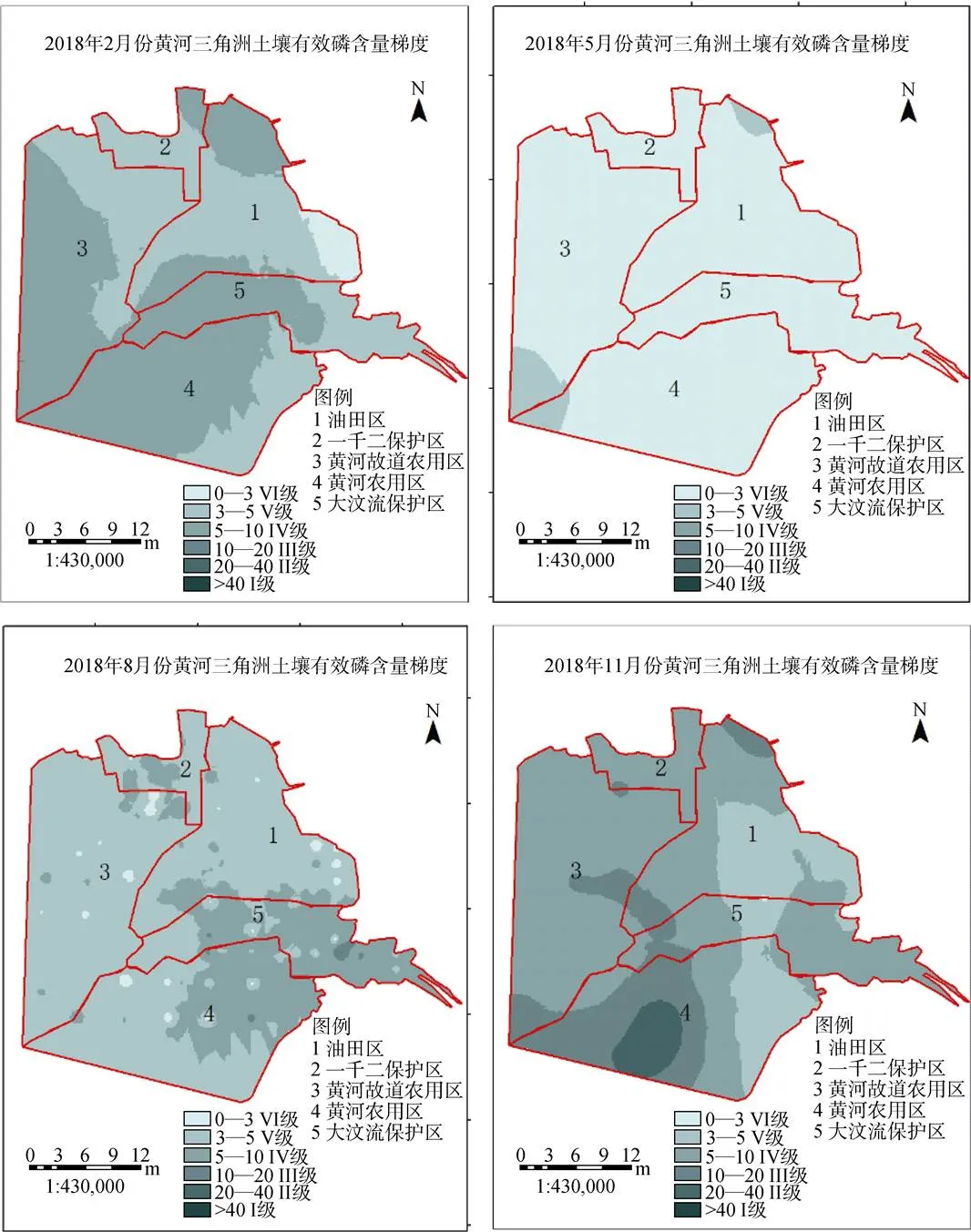
图9 2017年4个季度黄河三角洲有效磷含量梯度变化
Figure 9 Spatial gradient change of AP in four quarters
2.3.4 土壤有效钾(AK)
研究区不同季节的AP呈现出不规律空间梯度分布, 区域AP相关度不高。整体而言, AK色度偏红色, 说明研究区土壤AK含量高。而不同季节的AK呈现出相同的空间梯度变化规律, 沿海AK浓度远远高于内地地区, 呈现沿海向内陆浓度递减的规律。以东南沿海带以及北部沿海带为典例, 沿海区域呈环带状向内陆递减。2017年黄河三角洲滨海湿地年均AK以III级标准土壤为主, 占34.02%, 整体处于中等水平(图10)。
从不同景观类型分析: 农田区AK要低于其他景观类型, 全年未达到III级标准的区域为农田1区, 而全年呈深红色的区域(达到I、II级标准)的区域有光滩盐田区北部、植被农田区东部(黄河口生态保护区)及农田2区西北部。第二: 光滩盐田区年均最高, AK 值为151.22 mg·kg–1; 植被农田区年均AK为88.77 mg·kg–1为最低, 但植被农田区内部区域梯度差异明显, 西高东低。

图10 2017年4个季度黄河三角洲土壤有效钾梯度变化
Figure 10 Spatial gradient change of AK in four quarters
2.3.5 土壤全氮(TN)
根据2017年春冬两季TN呈现出梯度规律显示: TN含量整体处于较低水平, 西高东低(图11)。通过对比景观类型图土壤肥力标准高的地区在黄河三角洲中部以及西北部的农田区、东北区的盐田色度偏浅, 为IV级土壤, 其余地区均为V标准以上的土壤。面积总和7392.6 km2, 占29.61%; VI级标准面积2496.8 km2, 占8.13%。
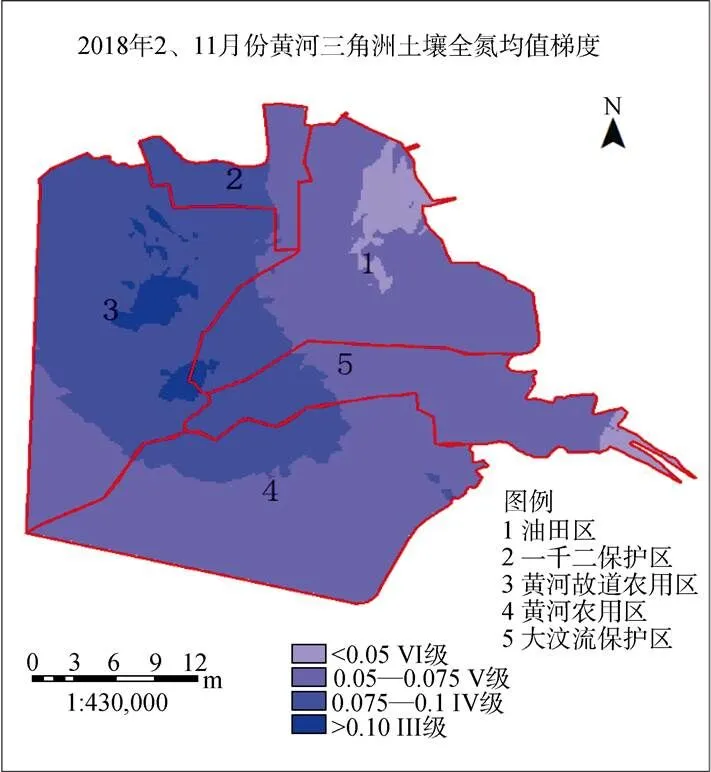
图11 2017年2个季度黄河三角洲TN含量梯度变化
Figure 11 Spatial gradient change of TN in two quarters
2.3.6 土壤pH
研究区不同季节的土壤pH之间呈现不规律分布的特点, 沿海高内陆低。如图12所示: 光滩盐田区以及植被农田区季节pH波动不大且色度偏深色, 年均pH达到8.21、8.11, 高于其他三个区。两个农田区季节pH波动较大、整体色度偏浅色, pH值高。
2.4 土壤因子相关性分析
对91个站位的土壤因子进行相关性分析, 结果显示: 土壤TN与AP在0.01水平上显著负相关(P<0.01); 土壤TN与SA在0.01水平上显著负相关。土壤SA与AP含量在0.05水平上显著负相关(P<0.05); 土壤TN与AK在水平上显著负相关(表6)。可见, 土壤全氮含量存在指向性特征, 对其他土壤因子影响更大, 表明研究区土壤其他因子受土壤全氮含量的限制更明显。
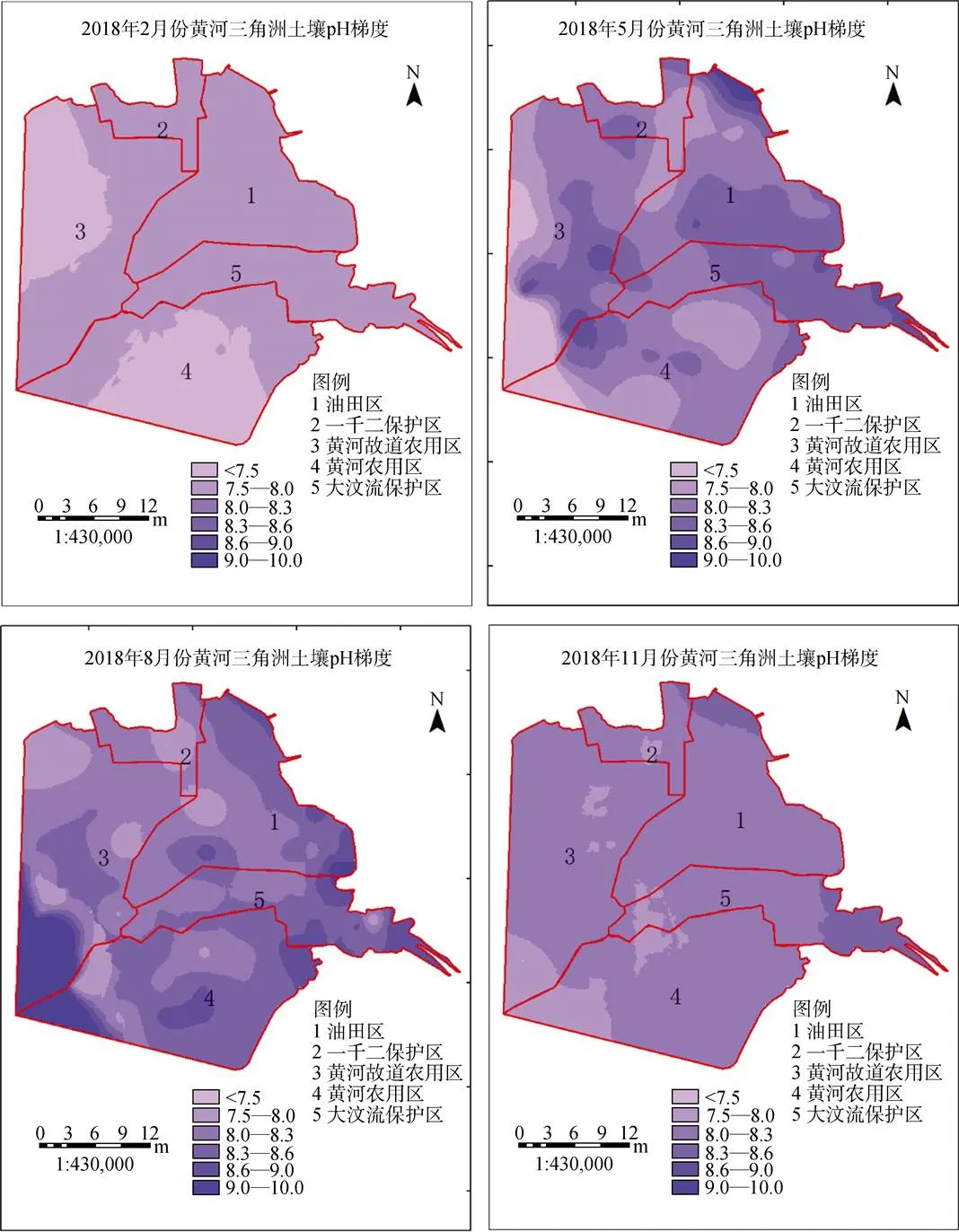
图12 2017年4个季度黄河三角洲土壤pH值梯度变化
Figure 12 Spatial gradient change of AK in four quarters
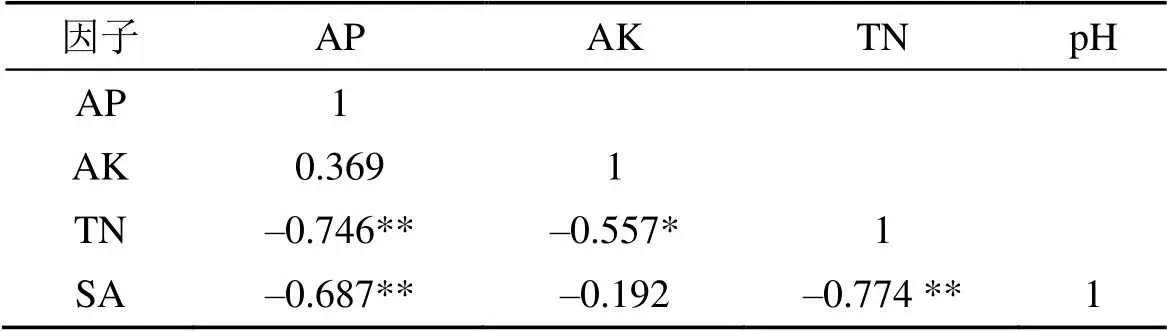
表6 土壤因子之间相关性分析
注:*在 0.05 水平(双侧)上显著相关; **.在 0.01 水平(双侧)上显著相关。
2.5 植被因子与土壤因子相关性分析
对植物生物量、Shannon-wiener指数、植被覆盖度与土壤因子、pH值, 按照季节进行相关性分析, 结果显示: 春季植物AK与Shannon-wiener指数在0.01水平上显著负相关, 与植被覆盖度在0.05水平上显著负相关; 秋季植物AK与Shannon-wiener指数在0.05水平上显著负相关; 春秋两季SA与植被覆盖度在0.05水平上显著负相关。说明在土壤AK成为植物的多样性以及植物覆盖率的主要控制因子(表7)。
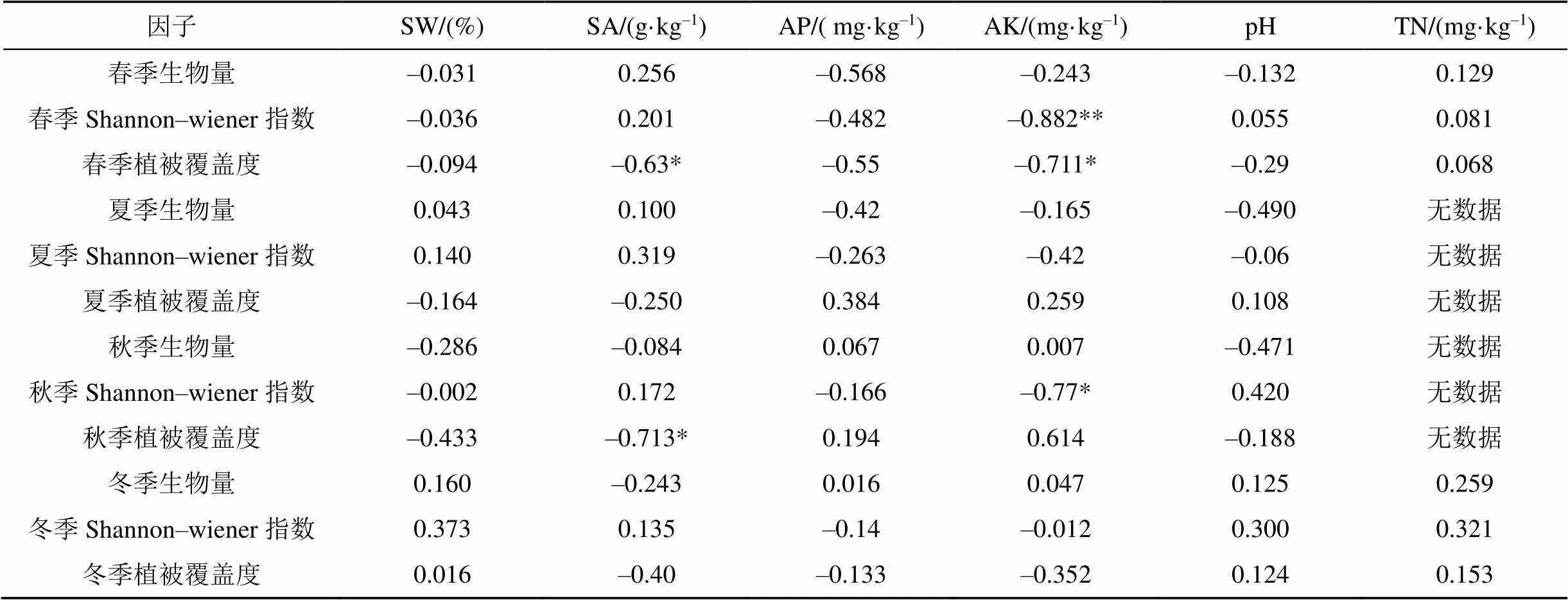
表7 土壤因子与生物量、生物多样性指数、植被覆盖度相关性分析
注:*在 0.05 水平(双侧)上显著相关; **.在 0.01 水平(双侧)上显著相关。
3 讨论
土壤肥力流失已成为湿地最严重的生态问题。按照国家养分含量标准, 湿地土壤全氮含量、有效磷含量总体严重偏低。湿地土壤为贫营养状态, 土壤盐度是滨海地区土壤养分的限制因子[33], 本文通过相关性分析证实了该观点。相比之下, 在空间分析中, 土壤盐渍化主要受景观类型的影响, 光滩盐田区的盐度十分高, 对离岸距离不敏感, 说明盐水入侵不明显。另外, 土壤肥力和土壤盐渍化是土地利用变化的主要原因, 也是自然环境长期积累的结果, 严重威胁着湿地的生态稳定[34-35]。
人类活动对自然生态系统的负面影响[36]。研究区天然湿地植被面积少, 盐田和养殖池等人工湿地面积大, 影响了湿地植被生长及土壤涵养, 例如, 养殖池的污水会污染土壤、卤水的大量开采会引发湿地土壤盐渍化, 导致植物生长受阻, 进而导致土壤质量下降。这是造成研究区养分匮乏, 流失严重的主要原因。
针对湿地现状, 提出以下建议。一是, 土壤肥力等级、含盐量是景观形成差异或是景观变迁的主要原因, 也是自然环境长期累积的结果, 并严重威胁研究区内的生态稳定性, 难以通过系统自身的调节恢复到良好的状态, 因此, 涵养水源, 修复湿地, 土壤污染修复, 通过自然物质以及能量的流动使湿地生态系统恢复到健康的“本底”。二是, 人类活动不仅干扰自然生态系统, 还包括生态建设和管理, 如自然保护区的建立和规范, 土壤盐渍化问题的持续治理, 它们在维护自然生态系统方面发挥了重要作用[36]。
4 结论
以黄河三角洲湿地为研究区, 构建了湿地景观类型和主要影响因子的空间分布格局, 解析不同景观类型下的植被及土壤因子的空间分布特征以及耦合关系。本研究可得到以下几个结论:
(1)根据全国养分含量等级标准, 2018年黄河三角洲滨海湿地AP以IV级标准土壤为主, 占39.96%, 整体处于缺乏状态; TN以IV级标准土壤为主, 占54.70%, 处于轻缺乏状态。
(2)植被和农田是研究区分布最广的景观类型, 也是生物量和植被覆盖度的主要贡献者。总量占比均在90%以上。农田相比其他景观显示出更大的季节波动性和空间异质性, 其土壤因子含量的季节性下降和增长都十分明显, 主要受农作物生长构成的限制。
(3)植被与土壤因子相关性分析结果: 春秋季AK与Shannon-wiener指数、植被覆盖度呈显著负相关(<0.01); SA与植被覆盖度相关系数呈显著负相关。AK成为植物生长、凋零过程中主控因子。
[1] 孙伟富. 1978—2009 年莱州湾海岸线变迁研究[D]. 青岛: 国家海洋局第一海洋研究所, 2010.
[2] 牛振国, 张海英, 王显威, 等. 1978—2008 年中国湿地类型变化[J]. 科学通报, 2012, 57(16): 1400–1411.
[3] 吴文挺, 田波, 周云轩, 等. 中国海岸带围垦遥感分析 [J]. 生态学报, 2016, 36(16): 5007–5016.
[4] 潘翔, 孙元敏, 吴剑, 等. 厦门湾海岸带景观格局时空动态演变研究[J]. 生态科学, 2016, 35(1): 117–123.
[5] 陈建, 王世岩, 毛战坡. 1976—2008年黄河三角洲湿地变化的遥感监测[J]. 地理科学进展, 2011, 30(5): 585–592.
[6] 许宁. 中国大陆海岸线及海岸工程时空变化研究[D]. 烟台:中国科学院烟台海岸带研究所, 2016.
[7] 谷东起, 付军, 闫文文, 等. 盐城滨海湿地退化评估及分区诊断[J]. 湿地科学, 2012, 10(1): 1–7.
[8] 吴涛, 赵冬至, 康建成, 等. 辽东湾双台子河口湿地翅碱蓬()生物量遥感反演研究[J]. 生态环境学报, 2011, 20(1): 24–29.
[9] SONG Wei, DENG Xiangzheng, YUAN Yongwei, et al. Impacts of land-use change on valued ecosystem service in rapidly urbanized North China Plain[J]. Ecological Modelling, 2015, 318 (24): 245–253.
[10] 蒋蕊竹, 李秀启, 朱永安, 等.基于MODIS 黄河三角洲湿地 NPP 与 NDVI 相关性的时空变化特征.生态学报, 2011, 31(22): 6708–6716.
[11] ANNAHEIMK E. Use of enzyme additions to characterize the nature and hydrolysability of soil organic phosphorus[J]. Tidsskrift for Den NorskeLgeforeningTidsskrift for Praktisk Medicin Ny Rkke, 2013, 91 (19): 279–285.
[12] BENBID K, BRAR S P S. Influence of soil organic carbon on the interpretation of soil test P for wheat grown on alkaline soils[J]. Fertilizer Research, 1994, 37(1): 35–41.
[13] SONG C, HAN X Z, TANG C. Changes in phosphorus fractions, sorption and release in Udic Mollisols under different ecosystems[J]. Biology & FertilityofSoils, 2007, 44(1): 37–47.
[14] 贺强, 崔保山, 赵欣胜, 等. 水盐梯度下黄河三角洲湿地植被空间分异规律的定量分析[J]. 湿地科学, 2007, 5(3): 208–214.
[15] SHARMA K N, SINGH H, VIG A C. Influence of continuous cropping and fertilization on adsorption and desorption of soil phosphorus[J]. Fertilizer Research, 1994, 40(2): 121–128.
[16] 朱新玉. 黄河故道湿地土壤质量因子与景观类型的耦合关系[J]. 资源科学, 2015, 37(1): 85–93.
[17] ZHANG Yili, WANG Chunlian, BAI Wanqi, et al. Alpine wetlands in the Lhasa River Basin, China[J]. Journal of Geographical Sciences, 2010, 20(3): 375–388.
[18] TORMA A, CSÁSZÁR P. Species richness and composition patterns across trophic levels of true bugs (Heteroptera) in the agricultural landscape of the lower reach of the Tisza River Basin[J]. Journal of Insect Conservation, 2013, 17(1): 35–51.
[19] SUN Haimin, MENG Wei, MU Chunsheng. The influence of plant diversity and aboveground biomass on soil microbial diversity in alkali–saline grassland [J]. Fresenius Environmental Bulletin, 2014, 23(3): 763–768.
[20] CHEN Dima, CHENG Junhui, CHU Pengfei, et al. Regional–scale patterns of soil microbes and nematodes across grasslands on the Mongolian plateau: relationships with climate, soil, and plants [J]. Ecography, 2015, 38(6): 622–631.
[21] 苏德纯. 从土壤磷的空间分布特征探讨提高磷肥及土壤磷有效性的新途径[J]. 中国农学通报, 1995, 11(1):38–40.
[22] 李远, 章海波, 陈小兵, 等. 黄河三角洲内陆到潮滩土壤中碳、氮元素的梯度分布规律.地球化学, 2014, 43(4): 338–345.
[23] 李玉, 康晓明, 郝彦宾, 等. 黄河三角洲芦苇湿地生态系统碳、水热通量特征.生态学报, 2014, 34(15): 4400–4411.
[24] 初小静, 韩广轩, 朱书玉, 等. 环境和生物因子对黄河三角洲滨海湿地净生态系统 CO2交换的影响. 应用生态学报, 2016, 27(7): 2091–2100.
[25] 莱州湾滨海湿地人类活动强度及其生态效应[D]. 青岛理工大学, 2018.
[26] 张绪良, 叶思源, 印萍, 等. 黄河三角洲自然湿地植被的特征及演化[J]. 生态环境学报, 2009, 18(1):292–298.
[27] 陈爱莲, 朱博勤, 陈利顶, 等. 双台河口湿地景观及生态干扰度的动态变化[J]. 应用生态学报, 2010, 21(5): 1120–1128.
[28] 刘峰. 黄河三角洲湿地水生态系统污染、退化与湿地修复的初步研究[D]. 青岛: 中国海洋大学, 2015.
[29] 任继周.草业科学研究方法[M].北京: 中国农业出版社, 1998: 95.
[30] 乌仁其其格. 草甸草原不同退化程度下植被与土壤特征与指示度研究[D]. 呼和浩特: 内蒙古农业大学, 2009.
[31] 池源, 石洪华, 孙景宽, 等. 近30年来黄河三角洲植被净初级生产力时空特征及主要影响因素[J]. 生态学报, 2018, 38(8): 2683–2697.
[32] 吴爱琳. 长江三角洲地区土地生态敏感性时空分析[D]. 上海: 东华大学, 2017.
[33] 张光亮, 白军红, 郗敏, 等. 黄河三角洲湿地土壤质量综合评价[J]. 湿地科学, 2015, 13(6): 744–751.
[34] Wang Hui, Wang Renqing, Yu Yue, et al. Soil organic carbon of degraded wetlands treated with freshwater in the Yellow River Delta, China[J]. Journal of Environmental Management, 2011, 92(10), 2628–2633.
[35] Daliakopoulos I, Tsanis I, Koutroulis A, et al. The threat of soil salinity: A European scale review[J]. Science of the Total Environment, 2016(15), 573, 727–739.
[36] Chi Yuan, Shi Honghua, Zheng Wei, et al. Spatiotemporal characteristics and ecological effects of the human interference index of the yellow river delta in the last 30 years[J]. Ecological Indicators, 2018, 89: 880–892.
Spatial characteristics of vegetation and soil factors under different landscape types in the Yellow River Delta Wetland
LU Jingfang1, SHI Honghua2,3, SUN Hui4, LI Jie1,*, HUANG Jiantao1, LIU Yiman1
1. School of Environmental and Municipal Engineering, Qingdao University of Technology, Qingdao 266033, China 2. Qingdao National Laboratory of Marine Science and Technology Marine Geological Process and Environmental Function Laboratory, Qingdao 266061, China 3. First Institute of Oceanography, Ministry of Natural Resources, Qingdao 266061, China 4. Branch Center of Operation and Maintenance Center of Shandong Water transfer Project,Qingdao 266300, China
The study was conducted in the Yellow River Delta Wetland (YRDW). The purpose of this study is to investigate the spatial characteristics of vegetation and soil factors, so that the coupling relationship can be applied in different wetland landscape types. The results indicated that farmland and wetland vegetation were the two largest landscape types in YRDW, occupying about 46.87% and 20.6%. They provided abundant biomass and vegetation coverage, as an important supplement for wetland blue carbon. In addition, spatial distribution characteristics of soil factors was determined by landscape types. The annual average available phosphorus (AP) was in a state of deficiency, at grade IV standard, and the highest content in the farmland; the annual average available potassium (AK) was in a medium state, at grade IV standard and the highest content in the bare area. Finally, there was a significantly negative correlation between effective potassium and Shannon-wiener index, which became the main controlling factor in plant growth and zeroing.
Yellow River Delta; coastal wetland; landscape type; soil factor; vegetation
10.14108/j.cnki.1008-8873.2021.02.020
文献标识码:A 文章编号:1008-8873(2021)02-157-08
2019-08-07;
2019-12-03
山东省专项海洋科技资金重大科技创新工程(2018SDKJ0505-1); 国家自然资源部研究课题(A2018-099); 国家海洋公益性行业科研专项(201505001); 山东省黄河三角洲生态环境重点实验室开放课题(2016KFJJ01)
路景钫(1994—), 男, 山东潍坊人, 硕士研究生, 主要从事环境生态学研究。E-mail: 975477978@qq.com
李捷, 男, 博士, 教授, 主要从事环境生态学研究, E-mail: jli@qut.edu.com
路景钫, 石洪华, 孙辉,等. 黄河三角洲湿地不同景观类型下植被与土壤因子的空间特征[J]. 生态科学, 2021, 40(2): 157–166.
LU Jingfang, SHI Honghua, SUN Hui, et al. Spatial characteristics of vegetation and soil factors under different landscape types in the Yellow River Delta Wetland [J]. Ecological Science, 2021, 40(2): 157–166.
