氮添加对黄河三角洲高潮滩芦苇生长和土壤理化性质的短期效应
2021-04-13赵连军张俪文衣华鹏兰思群陈琳路峰韩广轩
赵连军, 张俪文, 衣华鹏, 兰思群, 陈琳, 路峰, 韩广轩
氮添加对黄河三角洲高潮滩芦苇生长和土壤理化性质的短期效应
赵连军1,2, 张俪文2,*, 衣华鹏1, 兰思群1,2, 陈琳2,3, 路峰4, 韩广轩2
1. 鲁东大学, 资源与环境工程学院, 山东烟台 264025 2. 中国科学院烟台海岸带研究所, 中国科学院海岸带环境过程与生态修复重点实验室, 山东烟台 264003 3. 聊城大学, 环境与规划学院, 山东聊城 252000 4. 山东省黄河三角洲国家级自然保护区管理局, 山东东营 257091
氮富集会影响到全球生态系统的植物生长繁殖和土壤理化性质。然而, 目前关于氮富集对潮汐湿地生态系统植物生长和土壤理化性质的影响研究相对较少。通过氮添加野外控制实验, 研究了4个氮添加水平(CK: 0g·m-2·a-1、N1: 5 g·m-2·a-1、N2: 20 g·m-2·a-1、N3: 50 g·m-2·a-1)对黄河三角洲高潮滩芦苇植被区的芦苇生长和土壤理化性质短期效应。结果表明: N1添加水平显著增加芦苇高度, 增长节间长和叶片宽度, 但减短穗长; N2添加水平显著增加芦苇高度, 增长节间长和叶片长度, 增大叶片宽度, 但减少叶片数; N3添加水平则对芦苇生长繁殖无显著影响。但氮添加对芦苇密度、叶片全碳、全氮和碳氮比的影响不显著。而除了N3添加水平增加了NO3--N (0—10 cm)含量外, 其他土壤理化性质均不受氮添加的影响。因此, 在短期内适量氮输入可促进黄河三角洲高潮滩芦苇的生长, 过量的氮则对其生长没有作用, 保护区应控制对高潮滩芦苇植被过量的氮输入。
芦苇; 氮添加; 生长; 繁殖; 土壤理化性质; 黄河三角洲; 潮汐湿地
0 前言
植物在生长过程中需要从土壤中吸收大量的营养元素, 氮素是其中最重要的一种, 同时也是限制土壤生产力高低的主要因子之一。然而在人类活动的影响下, 氮沉降和农业施肥量日益增加, 进入生态系统的氮素越来越多[1], 土壤中的氮素含量也日益增高。氮富集可能会引起植物生理特性、土壤微生物活性及土壤理化性质的改变。例如, 氮素的持续输入会改变森林生态系统的植被组成结构, 并且有可能导致土壤酸化[2]; 氮素输入会改变草地生态系统的群落物种组成及优势种, 增加植被高度、盖度, 降低植被透光率, 使得物种丰富度降低[3]。氮输入增加是当前全世界共同面对的重要的环境问题, 也深刻影响着湿地生态系统的各种生态过程[4]。许多学者对氮输入可能带来的影响做了研究, 其中大多集中于森林生态系统[5-7]、草地生态系统[8-10]和农田生态系统[11-13], 对滨海潮汐湿地生态系统的研究相对较少, 且多集中于微生物和土壤呼吸方面的研究[14-15]。
氮富集会对湿地生态系统产生明显影响[16]。在湿地生态系统中氮是一种限制性资源, 对植物生长至关重要[17]。不同植物对氮素的吸收利用效率有所不同, 所以氮富集对不同植物的影响程度会有所差异。例如, 在氮素不足的情况下, 芦苇的叶、茎形态会发生改变, 低氮会抑制芦苇生长, 降低生物量, 增加根冠比[18]; 氮输入量的增加会显著增加互花米草的分株数、叶片数、叶片长度、叶片宽度和叶面积等生长繁殖性状[19]。因此每一种植物有其营养耐受范围, 低于或超出这个范围会不利于植物生长。施氮的影响与添加浓度有关, 一般情况下氮素缺乏会不利于植物生长, 过量的氮素则会抑制植物生长, 适量增加氮的供应能促进植物生长[5,20]。
黄河三角洲滨海湿地位于黄河入海口, 是中国暖温带地区最完整、最广阔、最年轻的新生湿地, 增长迅速且面积广阔, 具有净化污染物、固沙促淤、调节气候、为野生动植物提供适宜生境等多种重要的生态功能, 具有重要的生态保护价值。宁凯等[21]研究表明, 大气氮沉降已成为该地区土壤氮素的主要来源之一。同时, 人类活动排放的生活污水和工业废水日益增加, 黄河径流带来的无机氮、无机磷等污染物的不断输入, 该区域的养分状况发生了显著改变[22]。
芦苇()属于多年生根茎型禾本科植物, 抗逆性强, 生长迅速, 繁殖能力强, 尤其有着极好的耐盐特性, 因而有着广泛生境适应能力[23-24], 成为盐碱湿地的主要建群种和湿地生态恢复的工具种, 对维持湿地生态系统功能和生态平衡具有非常重要的作用。芦苇是黄河三角洲滨海湿地重要的植被类型, 大面积分布于黄河三角洲河口湿地、非潮汐湿地和盐沼。生长在盐沼高潮滩区域的潮水芦苇, 其种群面积在黄河三角洲湿地较少, 但是其遗传多样性却比其他生境的芦苇种群高[25]。
目前, 关于野外不同氮输入水平下芦苇的生长状况的研究较少, 特别是黄河三角洲滨海湿地生态环境脆弱, 探讨氮输入与芦苇生长的关系有助于对这一区域的芦苇植被的保护与恢复提出合理措施。因此, 本文提出以下科学问题: (1)黄河三角洲高潮滩芦苇个体性状对短期氮添加如何响应?(2)黄河三角洲高潮滩芦苇植被区土壤理化性质对短期氮添加如何响应?通过氮添加野外控制实验回答这两个科学问题, 旨在为黄河三角洲湿地生态保护提供理论依据和实践参考。
1 研究地区与研究方法
1.1 研究区概况
研究区位于山东省东营市境内的黄河三角洲国家级湿地自然保护区(37°40′ N—38°10′ N, 118°41′ E—119°16′ E)内, 是以保护黄河口原生湿地生态系统和珍稀濒危鸟类为主体的自然保护区。黄河三角洲是黄河携带大量泥沙在渤海凹陷处沉积形成的冲积平原。气候属暖温带大陆性季风气候, 年平均气温11.7—12.8 ℃, 年平均降雨量530—630 mm, 其中70%在夏季。年蒸发量为1900—2400 mm, 潮汐为非规则半月潮和半日潮。该地区地势平坦, 主要优势植物为: 盐地碱蓬()、芦苇、柽柳()和互花米草(), 土壤类型为隐域性潮土和盐土。
1.2 研究方法
1.2.1 样地选择和样方设置
本研究于2017年5月中旬在黄河三角洲国家级自然保护区大汶流管理站的高潮滩芦苇植被区(37°44'5.46"N, 119°12'56.11"E)设置样方(图1), 该植被区为单一芦苇种群, 受不规则半日潮影响, 春末夏初潮汐频率低、淹水时间短, 夏末和秋冬季潮汐频率高、淹水时间长, 潮汐受风向影响大。选择地势相近、芦苇生长状况均一(2017年5月, 各样方芦苇高度均值为(48.08±0.58) cm, 平均密度为(11.72±0.69) 株·m-2; 不同氮处理样方间芦苇高度不存在显著差异, Kruskal-Wallis χ²为3.55,=0.31; 密度在不同处理间不存在显著差异, F=0.625,=0.607。)的地块作为实验样地。设置4个氮添加水平: CK: 0 g·m-2·a-1、N1: 5 g·m-2·a-1、N2: 20 g·m-2·a-1、N3: 50 g·m-2·a-1, 每个水平设置6个重复共建立24个2 m × 2 m长期实验样方, 样方之间间距10 m以上。每年5月中旬、7月中旬添加尿素[CO(NH2)2, 含氮46%], 将尿素均匀地撒在样方内。
1.2.2 芦苇生长和繁殖性状测定
芦苇生长性状测定于2017年9月中旬(芦苇生长季末期)进行, 测量每一个样方内所有芦苇的植株高度、节间长、叶片数、叶片宽度、叶片长度和穗长。芦苇叶片全碳(TC)、全氮(TN)的测定, 在各样方中采集10片左右完整叶片, 置于80 ℃烘箱中烘干至恒质量, 磨碎后过100目, 将样品送至中国科学院烟台海岸带研究所分析测试中心使用元素分析仪测定。
1.2.3 土壤理化性质测定
土壤样品采集于2017年9月中旬进行, 在每个样方内使用内径为2.5 cm的土钻分0—10 cm和10—20 cm两个土层进行取样, 共采集了48个土壤样品。将采集好的土壤样品带回实验室, 取一部分新鲜土样放入冰柜冷冻保存待测; 另一部分经自然风干后待测。
图1 样地所在位置示意图
Figure 1 Location of the study site
采用新鲜土样测定土壤含水率、铵态氮(NH4+-N)和硝态氮(NO3--N)。土壤含水率采用烘干法(105 ℃烘干)测定。土壤铵态氮(NH4+-N)和硝态氮(NO3--N)采用2 mol·L-1KCl浸提, 浸提液利用流动分析仪(AutoAnalyzer III, Bran+Luebbe GmbH Germany)测定。
自然风干后的土样将根和石块挑出, 磨碎后过100目土壤筛用于测定其他土壤理化性质。土壤酸碱度(pH)、电导率(EC)采用pH计(QT-PH220S)和电导率仪(DDBJ-350)测定, 水土质量比为5: 1。土壤全碳(TC)、全氮(TN)、全硫(TS)的测定, 将样品送至中国科学院烟台海岸带研究所分析测试中心使用元素分析仪测定。
1.3 数据分析
数据使用R软件(R i386 3.5.3 version)进行分析。对于平衡、满足残差正态分布和方差齐性的数据, 采用单因素方差分析法(One-way ANOVA)进行方差分析, 并采用Tukey HSD法进行多重比较。对于平衡但不满足正态性检验和方差齐性的数据(如芦苇密度、叶片TC、叶片TN、叶片C: N)和不平衡数据(芦苇高度、节间长、叶片数、叶片宽度、叶片长度、穗长)则采用非参数统计方法Kruskal-Wallis检验, 非参数多重比较(Multiple comparison test after Kruskal-Wallis)。采用Origin 9.0软件绘图, 文中数据为平均值±标准误。
2 结果
2.1 氮添加对高潮滩芦苇生长的短期效应
芦苇平均高度为(77.98±0.61) cm, 平均节间长为(5.15±0.05) cm, 平均叶片数为(9.06±0.09)片, 平均叶片宽度为(1.86±0.01) cm, 平均叶片长度为(16.79±0.15) cm, 平均穗长为(1.72±0.11) cm。芦苇叶片TC含量均值为(461.57±2.50) g·kg-1, TN含量均值为(25.52±0.43) g·kg-1, C: N均值为18.18±0.28。
结果表明, 氮添加水平对芦苇生长性状: 高度、叶片数、节间长、叶片宽度、叶片长度、穗长均有显著影响(表1;<0.05), 对芦苇密度、叶片养分(TC、TN、C: N)无显著影响(表2;>0.05)。
2.1.1 氮添加对高潮滩的芦苇密度的短期效应
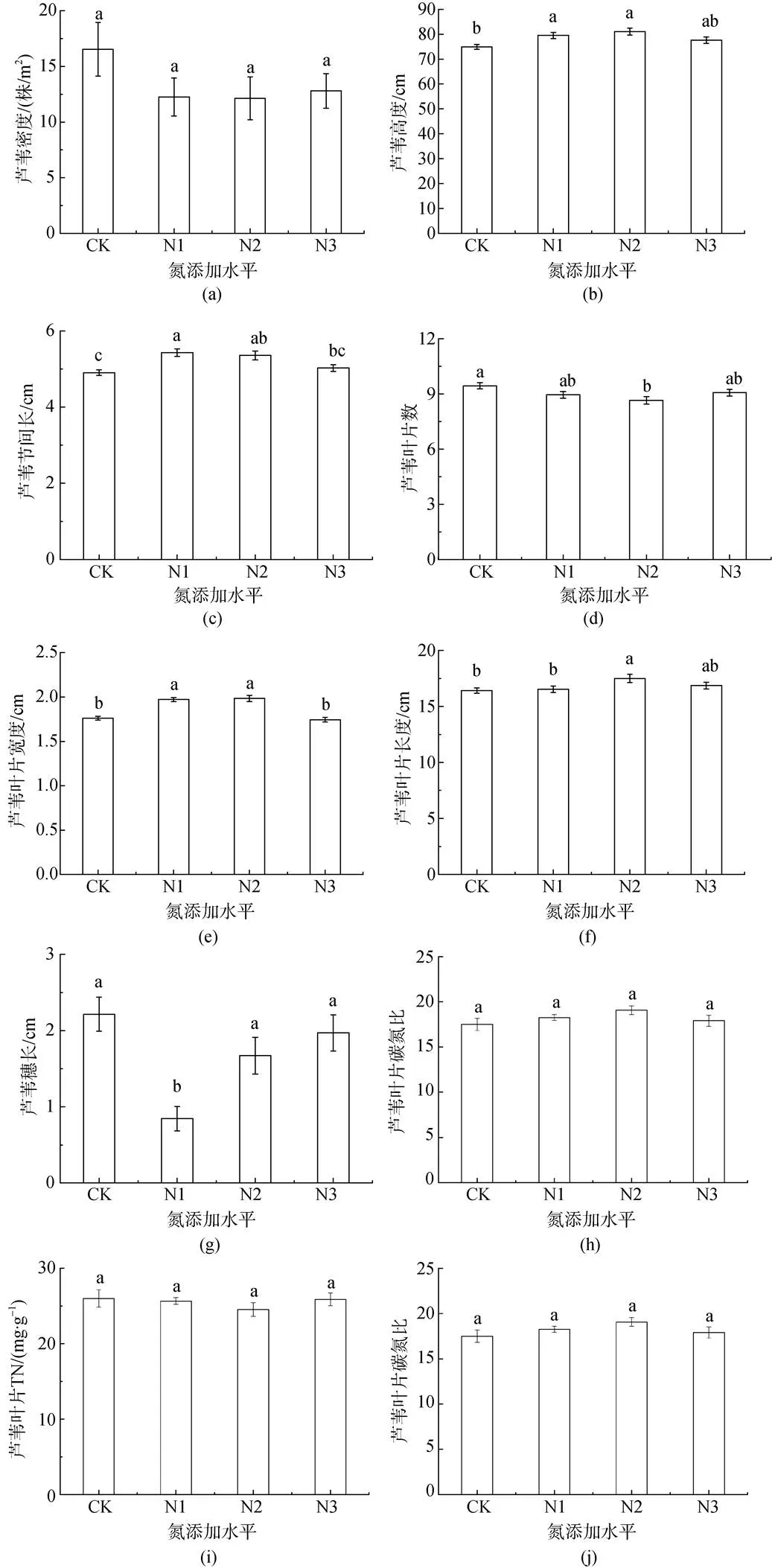
结果表明, 芦苇平均密度为13.4 株·m-2, 各氮添加水平下芦苇的密度无显著差异(>0.05) (图2a)。生长季结束时共有芦苇1289株。其中CK水平为397株(16.5 株·m-2), 占比30.8%; N1水平为294株(12.3株·m-2), 占比22.9%; N2水平为291株(12.1 株·m-2), 占比22.6%; N3水平为307株(12.8 株·m-2), 占比23.8%, 各氮添加水平下芦苇的密度无显著差异(>0.05)。24个样方中, 芦苇株数为21—93株, 平均53.7株(13.4 株·m-2)。
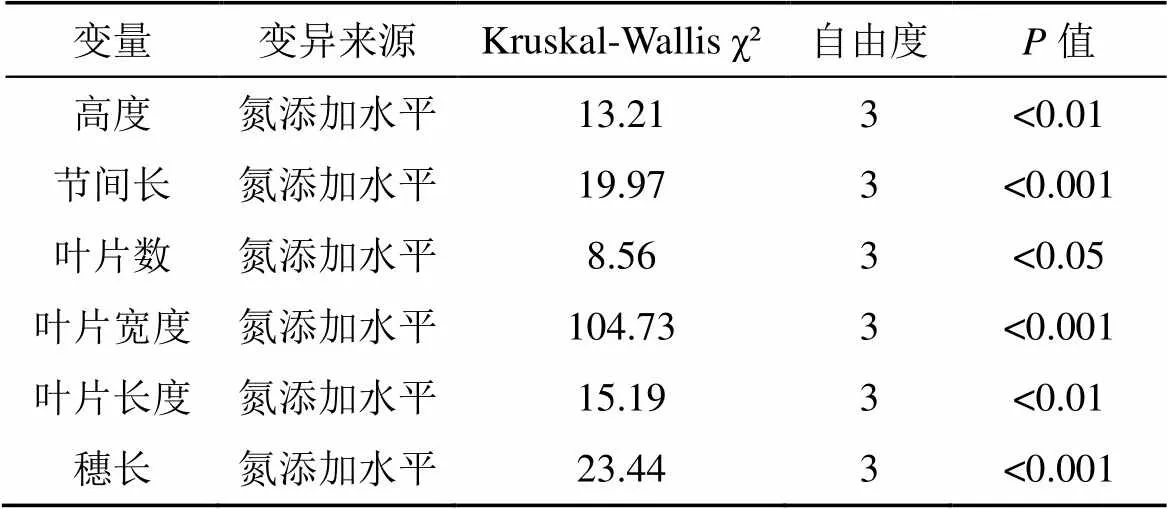
表1 氮添加对芦苇生长和繁殖性状短期效应的非参数统计检验结果
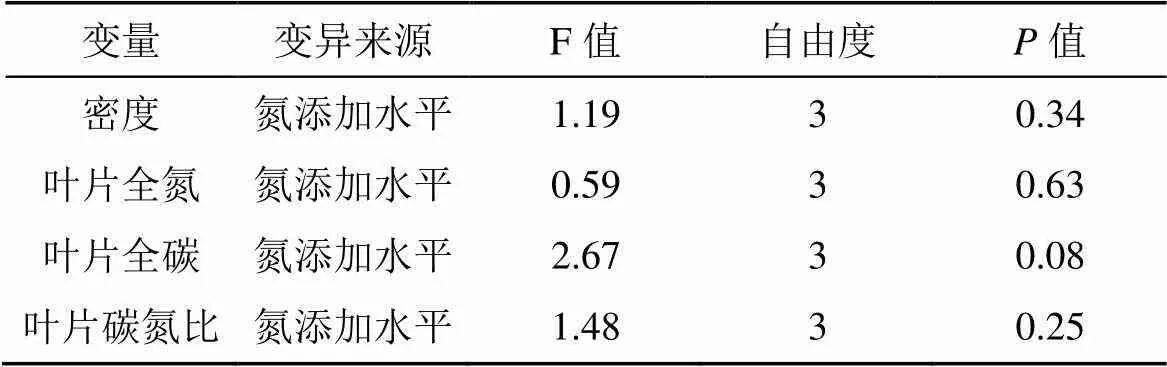
表2 氮添加对芦苇生长性状的短期效应的方差分析结果
2.1.2 氮添加对芦苇生长和繁殖性状的短期效应
各氮添加水平间芦苇高度、节间长、叶片数、叶片宽度、叶片长度、穗长均表现出显著差异(图2)。本实验各氮添加水平下芦苇高度的变化状况如图2所示。实验结果表明, 不同的氮添加水平下芦苇高度呈现出先升高后下降的状况(图2b)。芦苇高度在CK水平下表现为最低[(74.89±0.97) cm]; 在N1[(79.50±1.24) cm]、N2[(81.04±1.42) cm]两个水平下芦苇高度表现较为接近, 较之CK[(74.89±0.97) cm]分别显著提高了6%和8%(<0.05); N3[(77.61± 1.29) cm]较之N1、N2有所下降, 但差异并不显著(>0.05)。
节间长在N1[(5.43±0.09) cm]水平下表现最佳, 与N2[(5.35±0.11) cm]无明显差异(>0.05), 却明显高于较CK[(4.90±0.07) cm]和N3[(5.02±0.09) cm]分别显著提高了11%和8%(<0.05)(图2c)。
不同氮添加水平下芦苇叶片数呈现出先下降后上升的趋势, 在CK[(9.44±0.17) cm]水平下芦苇叶片数表现最优, 较N2[(8.66±0.20) cm]水平提高了9% (<0.05); 但CK、N2与N1[(8.96±0.19) cm]和N3[(9.07±0.19) cm]均无显著性差异(>0.05)(图2d)。
注: 相同的字母(a、b、c)表示在置信水平为0.05时, 多重比较没有显著差异, 不同的字母则表示多重比较有显著差异。误差线表示±SE。
Figure 2 The sh[1] ort-term effects of nitrogen addition ongrowth and reproductive traits
芦苇叶片宽度在CK[(1.76±0.02) cm]和N3[(1.74± 0.03) cm]两个水平下表现相近; N2[(1.98±0.03) cm]水平相比于CK和N3分别显著提高了12.5%和14% (<0.05); N2[(1.98±0.03) cm]与N1 [(1.97±0.02) cm]无明显性差异(>0.05)。叶片长度在N2[(17.50±0.36) cm]水平下芦苇叶片长度表现最高, 较CK[(16.42± 0.24) cm]和N1 [(16.53±0.30) cm]两个处理分别显著提高了7%和6%(<0.05); N2[(1.98±0.03) cm]与N3[(16.86± 0.29) cm]无显著性差异(>0.05)(图2f)。
在N1[(0.84±0.16) cm]水平下芦苇穗长表现最低, 较CK[(2.21±0.22) cm]、N2[(1.67±0.24) cm]和N3[(1.97± 0.24) cm]水平分别下降了62%、50%、57%, 呈现出显著性差异(<0.05)(图2g)。
2.1.3 氮添加对芦苇叶片养分(TC、TN、C: N)的短期效应
各个氮添加水平间芦苇叶片TC含量、TN含量以及C : N均无显著差异(图2h、i、j)。芦苇叶片TC含量较高, 为(434.79—495.06) g·kg-1, 均值为(461.57±2.50) g·kg-1; TN含量较低, 为(21.51—30.88) g·kg-1, 均值为(25.52±0.43) g·kg-1; C : N较大, 为14.92—20.87, 均值为18.18±0.28。
2.2 氮添加对土壤理化性质的短期效应
不同氮添加水平下, 研究区土壤基本理化性质见表3。研究区域内的土壤均为碱性土壤(pH>7)。由表3可知, 黄河三角洲高潮滩土壤0—10 cm和10—20 cm土层的pH值、电导率、含水率、全碳(TC)、全氮(TN)、碳氮比(C:N)、全硫(TS)、铵态氮(NH4+-N)、硝态氮(NO3--N(10-20 cm))在4个氮添加水平间均无显著差异(>0.05); NO3--N (0-10 cm)在CK水平下最低(0.45±0.09) mg·kg-1, 在N1[(0.55±0.06) mg·kg-1]和N2[(0.58±0.03) mg·kg-1]下几乎一致且与CK水平差异不显著(>0.05), 但在N3水平下表现最高[(0.77± 0.06) mg·kg-1], 比CK水平显著提高了71%(<0.05)。
3 讨论
高度、节间长是衡量芦苇长势的重要性状, 叶片的数量、宽度、长度更是直接关系到芦苇的光合作用, 穗长是芦苇繁殖能力的间接体现。光合作用的强弱决定着植物生长发育和生产力, 是研究环境因子对植物影响的重要指标[26], 高大的植株、宽阔的叶片更有利于芦苇进行光合作用促进生长发育, 因此高度、节间长、叶片数、叶片长度和叶片宽度都是决定芦苇生长发育的重要性状。在本研究中, 不同氮添加水平间芦苇生长和繁殖性状存在明显差异(<0.05)。N1添加水平显著增加芦苇高度, 增长节间长和叶片宽度; N2添加水平显著增加芦苇高度, 增长节间长和叶片长度, 增大叶片宽度; N3添加水平则对芦苇生长繁殖无显著影响。表明芦苇对施氮量有一定的耐受范围, 适当的氮添加量会促进芦苇的生长, 过高的施氮量对芦苇生长不起作用。
施氮所带来的影响与氮添加量有关, 一般情况下氮素缺乏不利于植物生长, 超出植物需求的氮素则会降低植物的生产力, 适量的氮添加会促进植物生产力的提高[5]。黄河三角洲高潮滩潮涨潮落频繁, 营养元素积累量较低, 加之土壤形成年龄短, 因而与内陆湿地生态系统相比, 土壤氮素含量明显偏低, 例如高原湿地纳帕海沼泽化草甸(云南)0—20 cm土层中, 全氮含量为(4.93±0.02) g·kg-1[27]; 三江平原湿地土壤中全氮含量高于17 g·kg-1[28], 远高于黄河三角洲高潮滩芦苇区域。研究区土壤中氮素含量偏低[0—10 cm和10—20 cm土层全氮含量分别为(0.45± 0.04) g·kg-1, (0.46±0.05) g·kg-1], 并未达到适宜芦苇生长所需的浓度。因此, 在黄河三角洲高潮滩湿地生态系统中氮是一种限制性资源, 适当提高土壤中的氮素含量, 可以促进该区域芦苇的生长。其他研究也表明, 随着氮添加的增多, 芦苇幼苗的高度、叶面积、地上生物量、地下生物量和总生物量都显著增加, 而比叶面积和根冠比降低[29]; 随着氮素输入的增加, 互花米草的分株数、叶片数、叶片长度、叶片宽度和叶面积等生长性状随之显著增加[19]。
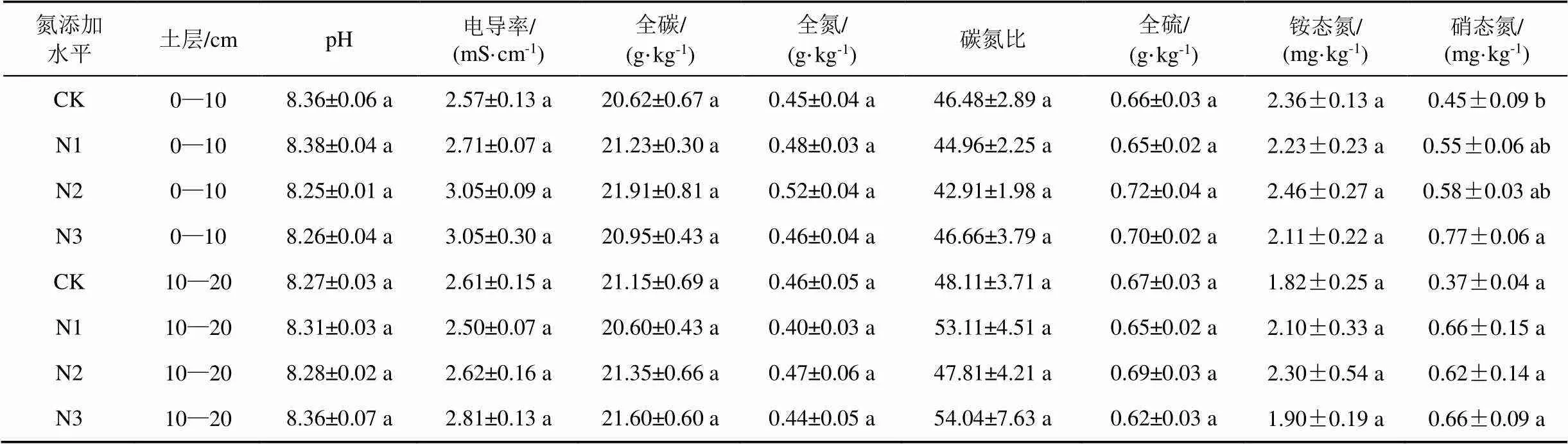
表3 氮添加对土壤理化性质的短期效应
注: 表中数据为Mean±SE; 同列数据不同字母表示多重比较显著差异(<0.05)
另外, N1添加水平显著增加芦苇高度, 增长节间长和叶片宽度, 但减短穗长。这表明, 在低氮素添加条件下芦苇的生长和繁殖可能表现出权衡关系。Silvertown和Lovett[30]指出: 植物在生长过程中, 会根据需求的不同调整资源分配, 不同的需求产生冲突导致植物在不同生长活动之间的权衡(trade-off)。有研究表明植物生长所投入的资源与投入到繁殖资源呈现出相反关系[31], 资源的可调节导致植物各器官权衡生长。例如, 不同的氮处理水平会显著影响燕麦的生长和繁殖资源分配, 燕麦的资源分配在三者间(地下生物量分配、茎叶生物量分配和繁殖生物量分配)存在权衡关系[32]。
叶片中TC、TN和C: N是反映植物生理代谢和生长状况的重要性状。研究发现, 不同氮添加水平间芦苇叶片中TC、TN和C: N无显著差异, 植物将营养用于生长, 可能是叶片养分无显著变化的原因。本研究还发现从1年实验尺度来看不同氮添加水平间芦苇密度差异不显著。多数湿地植物依靠无性繁殖进行种群更新、扩大生态位空间[33], 芦苇也不例外。芦苇天然种群主要依靠无性营养繁殖补充更新, 根状茎是芦苇进行无性繁殖的器官, 每年生长季芦苇都会在根状茎中存储大量的营养物质用于来年芦苇幼苗的繁殖[26,34]。上一年生长季结束时芦苇根状茎生物量的大小可能决定了第二年新生幼苗的数量, 因此短时间内氮添加水平可能不会对芦苇的密度产生明显影响。前人的研究表明, 氮添加会对湿地植物的无性繁殖能力产生影响[33]。短时间看, 不同氮添加水平间芦苇密度差异不显著(>0.05), 可能长期的氮素添加会对芦苇的地下根状茎生物量产生显著影响, 从而影响到芦苇的生长密度, 这一点有待进一步研究。
本研究发现氮添加对黄河三角洲高潮滩土壤中0—10 cm和10—20 cm土层的pH、EC、TC、TN、C: N、TS、NH4+-N、NO3--N (10—20) cm均无显著影响(>0.05)(表3)。氮添加对土壤理化性质的影响涉及到多种因素。可能氮添加本身不会对这些土壤理化性质性状产生显著影响, 也可能是被芦苇吸收用于植株生长。另外, 在潮滩频繁的潮汐作用也会带走一部分氮使得氮添加的量尚不足以对这些性质产生显著影响。土壤中的NH4+-N和NO3--N以不同的方式损失, NH4+-N容易以氨的形式挥发, 而NO3--N则容易随着降水淋失[35]。本研究中, 氮添加水平对0—10 cm和10—20 cm土层的NH4+-N、NO3--N (10—20) cm无显著影响(>0.05), 但不同氮添加水平下NO3--N(0—10) cm存在显著差异(<0.05)(表3)。NO3--N(0-10) cm在CK水平下最低[(0.45±0.09) mg·kg-1], 在N1[(0.55±0.06) mg·kg-1]和N2[(0.58± 0.03) mg·kg-1]下几乎一致且与CK水平差异不显著(>0.05), 但在N3水平下表现最高[(0.77±0.06) mg·kg-1],比CK水平显著提高了71%(<0.05)。表明N3水平显著增加了黄河三角洲高潮滩土壤表层中NO3--N的含量, 过量的氮添加可能导致黄河三角洲高潮滩土壤表层NO3--N过度残留。彭正萍等(2015)的研究结果也表明, 随着施氮量的增加, 氮素的损失量也会相应升高, 施用过多的氮肥非但不会明显提高作物产量, 反而会引起土壤中硝态氮大量残留[36]。
总之, 在黄河三角洲高潮滩湿地适当地添加养分, 可以促进该区域芦苇的生长。从一年实验尺度来看, 氮添加条件下, 芦苇高度、叶片宽度、叶片长度、节间长、穗长等性状均产生一定程度的改变。本实验将长期监测氮素输入对实验区域的芦苇生长和繁殖的长期效应, 并在未来实验中, 加强氮添加对根系生长、种子繁殖能力、土壤微生物多样性等的影响研究。
[1] PERRING M P, HEDIN L O, LEVIN S A, et al. Increased plant growth from nitrogen addition should conserve phosphorus in terrestrial ecosystems[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(6): 1971–1976.
[2] 吕超群, 田汉勤, 黄耀. 陆地生态系统氮沉降增加的生态效应[J]. 植物生态学报, 2007(2): 205–218.
[3] 李禄军, 曾德慧, 于占源, 等. 氮素添加对科尔沁沙质草地物种多样性和生产力的影响[J]. 应用生态学报, 2009, 20(08): 1838–1844.
[4] VITOUSEK P M, ABER J D, HOWARTH R W, et al. Human alteration of the global nitrogen cycle: sources and consequences [J]. Ecological Applications, 1997, 7(3): 737–750.
[5] 李德军, 莫江明, 方运霆, 等. 模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响[J]. 植物生态学报, 2005, 29(4): 543–549.
[6] 樊后保, 刘文飞, 裘秀群, 等. 杉木人工林凋落物量对氮沉降增加的初期响应[J]. 生态学杂志, 2007, 26(9): 1335–1338.
[7] 周学雅, 陈志杰, 耿世聪, 等. 氮沉降对长白山森林土壤团聚体内碳、氮含量的影响[J]. 应用生态学报, 2019, 30(5): 1543–1552.
[8] 刘红梅. 氮沉降对贝加尔针茅草原土壤碳氮转化及微生物学特性的影响[D]. 北京: 中国农业科学院, 2019.
[9] 潘庆民, 白永飞, 韩兴国, 等. 氮素对内蒙古典型草原羊草种群的影响[J]. 植物生态学报, 2005, 29(2): 311–317.
[10] 王伟, 刘学军. 青藏高原氮沉降研究现状及草地生态系统响应研究进展[J]. 中国农业大学学报, 2018, 23(5): 151–158.
[11] 崔键, 周静, 杨浩, 等. 我国红壤区大气氮沉降及其农田生态环境效应[J]. 土壤, 2015, 47(2): 245–251.
[12] 骆晓声, 石伟琦, 鲁丽, 等. 我国雷州半岛典型农田大气氮沉降[J]. 生态学报, 2014, 34(19): 5541–5548.
[13] 郝卓, 高扬, 张进忠, 等. 南方红壤区氮湿沉降特征及其对流域氮输出的影响[J]. 环境科学, 2015, 36(5): 1630– 1638.
[14] 吴松芹, 汪成忠, 李梦莎. 模拟氮沉降对滨海湿地土壤微生物功能多样性的影响[J]. 土壤, 2017, 49(6): 1153–1158.
[15] 朱敏, 张振华, 于君宝, 等. 氮沉降对黄河三角洲芦苇湿地土壤呼吸的影响[J]. 植物生态学报, 2013, 37(6): 517– 529.
[16] 胡星云, 孙志高, 孙文广, 等. 黄河口新生湿地碱蓬生物量及氮累积与分配对外源氮输入的响应[J]. 生态学报, 2017, 37(1): 226–237.
[17] CHEN Jianyao, TANIGUCHI M, LIU Guanqun, et al. Nitrate pollution of groundwater in the Yellow River delta, China[J]. Hydrogeology Journal, 2007, 15(8): 1605–1614.
[18] 初晓丹. 松嫩平原两种禾本科牧草对低氮胁迫的响应机制[D]. 长春: 东北师范大学, 2018.
[19] 张耀鸿, 张富存, 李映雪, 等. 外源氮输入对互花米草生长及叶特征的影响[J]. 生态环境学报, 2010, 19(10): 2297– 2301.
[20] 魏红旭, 徐程扬, 马履一, 等. 不同指数施肥方法下长白落叶松播种苗的需肥规律[J]. 生态学报, 2010, 30(3): 685–690.
[21] 宁凯, 于君宝, 屈凡柱, 等. 黄河三角洲滨海地区植物生长季大气氮沉降动态[J]. 地理科学, 2015, 35(2): 218– 223.
[22] YU Junbao, ZHAN Chao, LI Yunzhao, et al. Distribution of carbon, nitrogen and phosphorus in coastal wetland soil related land use in the Modern Yellow River Delta[J]. Scientific Reports, 2016, 6(1): 37940.
[23] 庄瑶, 孙一香, 王中生, 杨琳璐, 等. 芦苇生态型研究进展[J]. 生态学报, 2010, 30(8): 2173–2181.
[24] RUIZ M, VELASCO J. Nutrient Bioaccumulation in: Management Tool for Reduction of Pollution in the Mar Menor[J]. Water Air and Soil Pollution, 2010, 205(1/4): 173–185.
[25] 张俪文, 王安东, 赵亚杰, 等. 天津厚蟹植食对黄河三角洲高潮滩芦苇幼苗生长的影响[J]. 生态学杂志, 2018, 37(4): 1065–1070.
[26] 仲启铖, 王江涛, 周剑虹, 等. 水位调控对崇明东滩围垦区滩涂湿地芦苇和白茅光合、形态及生长的影响[J].应用生态学报, 2014, 25(2): 408–418.
[27] 任玉连, 陆梅, 范方喜, 等. 高原湿地沼泽化草甸土壤真菌与理化性质的关系[J]. 生态科学, 2019, 38(1): 42–49.
[28] YU Junbao, SUN Weidong, LIU Jingshuang, et al. Enhanced net formations of nitrous oxide and methane underneath the frozen soil in Sanjiang wetland, northeastern China[J]. Journal of Geophysical Research, Atmospheres, 2007, 112: doi: 10. 1029/2006JD008025.
[29] 潘艳文, 古勇波, 唐占辉, 等.盐度和氮添加对盐碱湿地芦苇幼苗生长及生物量分配的影响[J]. 土壤与作物, 2018, 7(2): 257–265.
[30] SILVERTOWN J W, LOVETT D J. Introduction to Plant Population Biology. Oxford: Blackwell Scientific Publications, 1993, 141–157.
[31] 赵志刚, 杜国祯, 任青吉. 5种毛茛科植物个体大小依赖的繁殖分配和性分配[J]. 植物生态学报, 2004, 28(1): 9–16.
[32] 赵宏魁, 马真, 张春辉, 等. 种植密度和施氮水平对燕麦生物量分配的影响[J]. 草业科学, 2016, 33(2): 249–258.
[33] 潘艳文. 盐碱湿地芦苇根茎出芽和幼苗生长的影响因子研究[D]. 北京: 中国科学院大学, 2018.
[34] 张永涛, 崔保山, 蓝艳, 等. 白洋淀不同年龄芦苇根状茎中氮和磷含量[J]. 湿地科学, 2013, 11(2): 286–291.
[35] 张亚欣, 范志平, 闫加亮, 等. 氮添加对沙质草地氨挥发及硝态氮淋溶的影响[J]. 生态学杂志, 2011, 30(9): 1969– 1974.
[36] 彭正萍, 刘亚男, 李迎春, 等. 持续氮素调控对小麦/玉米轮作系统氮素利用和表观损失的影响[J]. 水土保持学报, 2015, 29(6): 74–79.
第一行的纵坐标单位格式不对
The short-term effects of nitrogen addition on the growth ofand soil physical and chemical properties in a high marsh of the Yellow River Delta
ZHAO Lianjun1,2, ZHANG Liwen2,*, Yi Huapeng1, Lan Siqun1,2, Chen Lin2,3, Lu Feng4, HAN Guangxuan2
1. School of Resources and Environmental Engineering, Ludong University, Yantai Shandong 264025, China 2. CAS Key Laboratory of Coastal Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences (CAS), Yantai Shandong 264003, China 3. College of Environment and Planning, Liaocheng University, Liaocheng Shandong 252000, China 4. Administration Bureau of the Yellow River Delta National Nature Reserve, Dongying Shandong 257091, China
Nitrogen enrichment can affect plant growth and reproduction and soil nutrient cycling in ecosystems around the world. However, there are few studies on the effects of nitrogen enrichment on plant growth and soil physical and chemical properties in tidal wetland ecosystem. In this study, the short-term effects of four nitrogen addition levels ( CK:0 g·m-2·a-1, N1:5 g·m-2·a-1, N2: 20 g·m-2·a-1, N3:50 g·m-2·a-1) ongrowth and soil physical and chemical properties in thehigh marsh of the Yellow River Delta were studied through the field manipulative experiment of nitrogen addition. The experiment results showed that N1 addition level significantly increasedheight, internode length and leaf width, but decreased spike length. N2 addition level significantly increasedheight, internode length, leaf length, leaf width, but decreased leaf number. N3 addition level had no significant effect on the growth and reproduction of. However, the effects of nitrogen addition ondensity, leaf TC, TN and C:N were not significant. Except that N3 level increased the content of NO3—N(0-10) cm, other soil physical and chemical properties were not affected by nitrogen addition. Therefore, appropriate nitrogen addition can enhance the growth ofin the short term, but overload of nitrogen had no effect on the its growth. The nature reserve should control the overloading of nitrogen flowing into the high marshvegetation.
; nitrogen addition; growth traits; reproductive traits; soil physical and chemical properties; the Yellow River Delta; tidal wetland
10.14108/j.cnki.1008-8873.2021.02.003
Q945.79
A
1008-8873(2021)02-018-08
2019-10-20;
2019-12-03基金项目:国家自然科学基金项目(31670533); 中国科学院青年创新促进会(2018247); 山东省林业科技创新项目(LYCX07-2018-39)
赵连军(1993—), 男, 山东枣庄人, 硕士生, 主要从事滨海湿地生态学研究, E-mail: lianjunzhao@yic.ac.cn
张俪文, 女, 博士, 副研究员, 主要从事滨海湿地生态学研究, E-mail: lwzhang@yic.ac.cn
赵连军, 张俪文, 衣华鹏, 等. 氮添加对黄河三角洲高潮滩芦苇生长和土壤理化性质的短期效应[J]. 生态科学, 2021, 40(2): 18–25.
ZHAO Lianjun, ZHANG Liwen, Yi Huapeng, et al. The short-term effects of nitrogen addition on the growth ofand soil physical and chemical properties in a high marsh of the Yellow River Delta[J]. Ecological Science, 2021, 40(2): 18–25.
