单向明渠流与波浪作用下植被对水沙运动影响研究综述
2021-04-12张英豪赖锡军唐彩红
张英豪,赖锡军,唐彩红
(1. 聊城大学环境与规划学院,山东 聊城 252000;2. 中国科学院南京地理与湖泊研究所中国科学院流域地理学重点实验室,江苏 南京 210008;3. 华北电力大学水利与水电工程学院,北京 102206)
含植被水沙运动研究缘起于水生植被对洪水的影响。由于水生植被的存在增加了水流阻力,降低了河道行洪能力,水生植物被大量移除[1]。然而,水生植被具有重要的生态服务价值,使其成为淡水以及滨海生态系统中的重要组成部分。首先,水生植物为生态系统中其他生物提供食物来源,且通过吸收二氧化碳并释放氧气,是湿地生态系统中固碳的主要方式[2];其次,通过营造弱水动力环境,水生植物为一些鱼类以及大型底栖生物提供了多变的栖息环境[3];再次,水生植物可以吸收水体中过剩的营养盐,能够改善水质[4];最后,通过减流削浪,水生植物还能够起到保护岸坡以及减少侵蚀的作用[5]。因此,如何在发挥水生植被生态服务价值的基础上保障河湖的行洪安全与输水能力,使得含植被水沙运动研究受到广泛关注[6]。含植被水沙运动研究不仅能够为河道行洪能力设计、河湖水质净化以及生态修复、洲滩湿地演化等提供科学依据,具有很高的实际应用价值,还对水生态学、水文学、水力学以及紊流力学等具有重要的基础理论意义。
含植被水沙运动研究可追溯于20世纪30年代,最初主要探讨单向明渠水流条件下植生河道的水流阻力问题;20世纪70年代起,随着先进测试手段的应用,如声学多普勒流速仪(ADV)、激光多普勒测速仪(LDV)和粒子成像速度测量仪(PIV)等,获取水体高频三维脉动流速成为可能,含植被水流结构(尤其是水体紊动结构)的相关研究大量涌现。21世纪以来,单向明渠流条件下植被对水流结构的影响研究取得显著成就;受水动力条件控制,植被内的泥沙运动规律也被大量野外以及实验室研究所揭示。此外,波浪作为滨海以及内陆湖泊表层水体的又一种运动形式,波浪条件下植被对水沙运动影响的机理研究也逐渐受到广泛关注。含植被水沙运动研究已有的综述文章主要聚焦于单向明渠流条件下水生植被对水流结构的影响[7- 9],缺少单向明渠流条件下植被对泥沙运动影响以及波浪条件下植被对水沙运动影响规律的总结与概述。本文综述了单向明渠流以及波浪条件下水生植被对水沙运动影响的主要规律,并对今后的研究趋势进行展望。
1 水生植被对水动力的影响研究
1.1 单向明渠水流条件
1.1.1 水流阻力
水生植被对水流阻力的影响是含植物水沙运动研究中起步最早且历史最为悠久的部分。植被对水流阻力的影响通常采用曼宁糙率系数(n)进行表征,且受植被参数(形态、高度、柔韧度、分布密度、分布方式等)以及水流条件(流速、水深、相对淹没度等)的影响,不同植被对n值影响差异较大。
根据柔韧度,可将水生植物分为刚性植物(在水流条件下处于相对静止状态)与柔性植物(在水流条件下植株随水流会发生摆动)。刚性植物主要包括木本植物,如生长于河川泛洪区的高大乔木和分布于滨海湿地的红树林,以及禾本植物,如广泛分布于淡水河湖与盐碱湿地的芦苇。这类植物由于其高刚度的特质,一旦受水流冲击,将受到比柔性植物更为强大的冲击力,提供更大的水流阻力。实验室水槽研究中,通常将刚性植被简化为圆柱体,进而对其阻力进行估算[10- 11]。柔性植物在水流条件下能够发生摆动,一般指一些草本植物,如生长于淡水中的金鱼藻、苦草,以及生长于滨海湿地的大叶藻与日本鳗草等。单向明渠流作用下,柔性植被发生弯曲而呈现流线形,使得植被对水流的阻力较刚性植被相比明显减小,且随着植被柔性增加,植被高度(hv)与水深(h)之比逐渐减小,植被面积占整个断面的比例逐渐减小,n值随植被柔性的增加而减小[12- 13]。
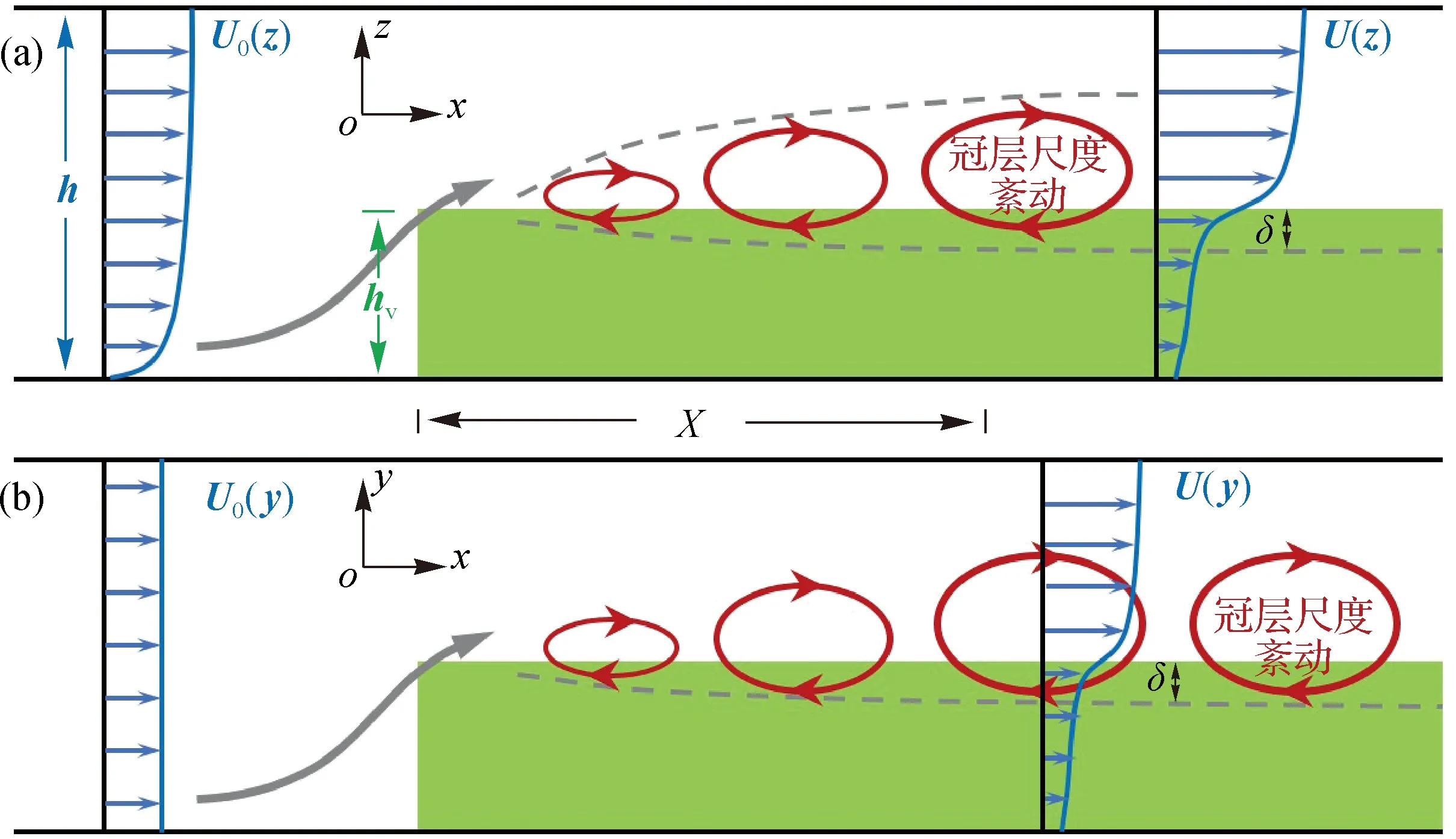
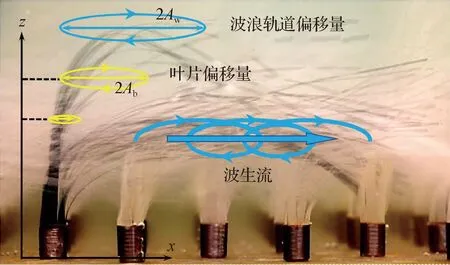
相对淹没度是影响水流阻力的另一重要因素。水生植被根据其淹没度可分为非淹没植被与淹没植被:前者指hv>h的植被,如位于潮侵浅滩地的盐藻植被以及生长于低潮位的海草冠层;后者hv 此外,Green[17]提出,植被对水流的阻力主要取决于河道中植被的空间分布。通过建立3种植被阻水因子(河道横断面植被面积占比、水面上植被面积占比以及植被体积占比),Green[17]发现植被阻水因子与n值间具有很好的线性关系。近些年,学者们还基于含植物明渠流体微元方法提出了含植物河道总曼宁系数[18],考虑局部植物分布导致的植物区与无植物区二次流影响的等效曼宁系数[19],以及漫滩植物复式河道的等效曼宁系数[20],这些均可用于描述不同植被分布形态下的植物阻力特征。 1.1.2 水流结构 紊流作为自然水体物质输运的基本动力,其结构特征直接影响着水体中物质的循环过程。水生植物的存在能够将水流的平均动能转化为紊动能,改变水体的紊动结构。含植物水流紊动结构研究对阐明水生植物生态环境效应的物理机制具有重要意义。 水流流经冠层时,水体在植株茎秆周围发生绕流,在茎秆后方可导致尾涡产生,形成尾流紊动(图1(a))。冠层内尾流紊动的产生取决于茎秆雷诺数(Red=Uν/d,U为冠层内时均流速,ν为水动力黏性系数,d为茎秆直径):当Red>120时,茎秆周围有尾流紊动产生;当Red<120时,无尾流紊动产生[21]。非淹没条件下,全水深范围内水体紊动以尾流紊动为主,且冠层内水体紊动能(turbulent kinetic energy,TKE)为时均流速、植被密度(m)以及植株形态参数的函数[22]。冠层内尾流紊动的尺度(lt)取决于植株茎秆直径(d)与相邻茎秆间距(S)[22],被称为茎秆尺度紊动。尾流紊动的存在可增加水体紊动能,但冠层的存在减小了时均流速。因此,冠层内紊动能水平随植被密度增加的变化主要取决于冠层内由时均流速减小引起的动能减小与由尾流紊动产生而引起的紊动能增加两者之间的相对大小,即随着植被密度的持续增加,冠层内紊动能水平较无植被条件相比,出现先增加后减小的变化特征[16,23- 24]。 淹没条件下,水体的水流结构与植被密度密切相关。当植被密度较小时,时均流速分布遵循紊动边界层剖面(对数剖面,图1(a)),且由于尾流紊动的产生,冠层内水体紊动能较无植被条件下增加[25]。随着植被密度增加,当冠层粗糙密度λf>0.1时(λf=ahv,a为冠层阻流面积)[26- 27],植被对水流的阻力使得时均流速剖面在冠顶附近(z≈hv)出现拐点(图1(b)和图1(c)),流速剖面与自由剪切层类似[28- 30]。时均流速剖面中拐点的出现,在植被顶部附近产生剪切紊动,且剪切紊动的紊动尺度与植被高度相当,被称为冠层尺度紊动(图1(b))。这种形式的紊动可影响自冠层顶部向下一定深度范围内(穿透深度,用δ表示)水体的紊动,并控制着植被冠层内部与其上覆水体的物质交换。Nepf和Vivoni[31]根据δ将冠层内部分为两部分:① 在不受剪切紊动影响的冠层底部(z 图1 不同植被密度条件下的水流时均以及紊动结构[9] 天然条件下,植被形态存在较大差异,不同形态的植被对水流结构的影响不同。通常,植被形态通过冠层阻流面积(a=md)以及体积分数(φ=mπd2/4)进行表征。非淹没条件下以及淹没条件下的冠层底部(水平交换区),由于水体雷诺切应力较小,水体时均流速与a呈反比[32- 33]。因此,对于叶基生的水生植物(如苦草等),由于植物叶分支位置以下a逐渐减小,使得时均流速在冠层内近床底处出现局部最大值[34- 35];相反,有些植物呈藤蔓状,叶片在全水深范围内分布均匀(a(z)=C,C为一常数),冠层内时均流速在全水深范围内也呈均匀分布[36- 37]。此外,天然状态下植被分布面积有限,往往呈斑块状[38]。水流流经植被斑块后,水流结构在斑块上游边缘以及两侧发生调整(图2),斑块边缘与内部的水流结构存在差异。淹没状态下,受植被阻力的影响,水流在进入植被斑块的开始阶段流速逐渐减小,剪切紊动在冠顶处逐渐发展(如图2(a)中红色圆圈所示),并在斑块内距上游边界一定距离处(X)达到稳定[39];在植被斑块两侧,植被区与无植被区的流速差异也可导致剪切紊动的产生(如图2(b)中红色圆圈所示),并影响斑块侧向物质的交换过程[40- 41]。Nepf等[27]指出,剪切紊动的垂向与侧向穿透深度(δ)以及植被斑块边缘处水流结构的调整长度(X)与冠层阻流面积成反比,即δ∝(CDa)-1与X∝(CDa)-1(CD为阻力系数)。植被斑块的存在,不仅影响斑块内部的水流结构,还改变着斑块周边水体的时均以及紊动结构。如图3所示,当水流遇到植被斑块后,受植被阻力影响,斑块内水体流速减小,斑块两侧流速增加;而在紧靠斑块的下游区域,水体时均流速与紊动能均减小,形成“遮蔽区”[42- 48]。 图2 水流结构在植被斑块上游以及侧向边缘的调整[39,41] 图3 植被斑块周围的水流结构[43] 波浪是区别于单向明渠流的又一种常见的水体运动形式,在滨海地区、内陆湖泊以及水库的表层水体,通常受到波浪的影响。目前,国内外关于植被与波浪相互作用的研究主要关注植被引起的波能衰减和植被对水流结构的影响。 1.2.1 波能衰减 水生植被对波能的减弱作用主要包括植被对波高以及波浪流速的影响。大量研究结果均表明,植被的存在能够有效减小波高,且水体与植物间的相对运动决定了波浪在冠层内的衰减程度[49- 51]。因此,植物对波高减弱作用的定量研究,关键在于如何确定植株与波浪间的相对运动。对于此,不同学者给出了不同的方法。例如,Mendez等[49]采用叶片偏移量(Ab,叶片轨道运动半径,图4)来刻画植物在波浪中的运动幅度,并认为叶片偏移量随叶片高度呈线性增加,从而得出植株与水体间的相对运动速度;Bradley和Houser[51]通过记录海草叶片顶部的运动,并采用悬臂模型将叶片顶部的偏移量演变为整个叶片的偏移量,从而有效刻画植株在波浪中的运动;Mullarney和Henderson[52]利用线性梁理论,建立了波浪作用下单株盐沼植物的动力模型;Luhar等[53]则采用有效叶片高度(产生相同波能衰减量条件下的刚性叶片高度)定量评价了柔性叶片对波高的减弱作用。植被对波浪流速的减弱作用首次被Lowe等[54]研究。采用刚性模型植被,Lowe等[54]建立了植被影响下冠层内部波浪轨道流速模型,并指出冠层内波浪流速的减弱程度与植被形态参数(如株高、株间距等)以及波浪参数(如波浪轨道偏移量(Aw,水质点轨道运动半径,图4))有关,且当Aw/S>1时,水生植被能够有效减小波浪流速。 图4 波浪作用下柔性植物叶片随水体摆动轨迹[54] 由于许多环境下水体运动受到波浪与水流的共同作用,一些学者对波浪- 水流共存场条件下水生植被对波高的减弱作用也开展了初步探究。Li和Yan[55]以及Hu等[56]以刚性植被为研究对象,分别采用三维RANS模型以及实验室水槽实验,发现与波浪条件相比,波- 流共存场条件下水生植物对波高的减弱作用加强。Paul等[57]以柔性模型植被为研究对象进行实验室水槽实验,却得出相反的实验结果,即水流与波浪的叠加减弱了植物的消波作用。此后,Losada等[58]采用真实植被开展水槽实验,发现当水流方向与波浪传播方向一致时,与波浪条件相比,水流的叠加使得植被的消波作用加强;相反,当水流方向与波浪传播方向相反时,植被的消波作用减弱。同时,考虑到水流能够导致植被发生倾伏,Losada等[58]以冠层倾伏高度代替冠层高度,为波- 流共存场条件下水生植被对波高削减作用的定量计算提供了一种新的方法。 1.2.2 水流结构 目前,波浪条件下植被对水流结构的影响研究受到忽视,研究成果较少。已有研究主要集中于冠层内波生流的出现以及冠层内的紊动特性。 Luhar等[59]以柔性模型植被为研究对象进行实验室水槽实验,发现冠层内部存在与波浪传播方向一致的定向水流(波生流,图4),并建立模型对冠层内水体的水深平均流速进行预测。Luhar等[60]又通过野外测量对其实验室结果进行了验证。此外,Abdolahpour等[61]从拉格朗日坐标系的视角出发,得出冠层内定向水流最大值的经验模型。van Rooijen等[62]通过实验室水槽实验与数值模拟研究了淹没条件下冠层内时均流速的垂线分布,发现波浪雷诺应力梯度是冠层内波生流出现的主要驱动力,且雷诺应力梯度的出现与植被顶部波浪轨道运动所产生的漩涡有关。 波浪作用下,植物茎秆周围也有尾流紊动产生。Sumer[63]指出,植株茎秆周围尾流紊动的出现受KC数(Keulegan-Carpenternumber,且KC=UmaxT/d,其中Umax为水质点轨道运动速度最大值,T为波浪周期)控制:当KC<6时,茎秆周围无尾流紊动产生;当KC>6时,有尾流紊动产生,且紊动影响范围随KC数增加而增加。Pujol等[64]利用在静水中振动栅格的方法产生波浪,发现刚性植被与柔性植被的存在均能够减小冠层内水体紊动能,且减弱程度随植株茎秆直径的减小以及植株密度的增加而逐渐增大。Pujol和Nepf[65]研究了破波条件下植物对紊动能的影响,指出由于植物可使波高降低,与无植物条件下相比,冠层在破波后对紊动能的减弱较强,但冠层内近床底处的紊动能却为无植物条件下的2倍,冠层内紊动能的增加主要是由于植株产生的尾流紊动。Pujol等[66]以及Ros等[67]研究了小波高条件下刚性和柔性植物对紊动的影响,发现刚性植物可增加水体紊动能,而柔性植物却降低了水体紊动能。Zhang等[68]认为,植被的存在能否增加冠层内部水体紊动能主要取决于Aw/S:当Aw/S<0.5时,冠层内部紊动能水平与无植物条件相当,冠层的存在没有显著增加水体紊动能;当Aw/S>0.5时,近床底紊动能显著增加,且植被内紊动能的大小可通过波浪流速、植株密度以及植被形态参数进行预测。 通过改变水动力结构(尤其是水体紊动结构),水生植被对泥沙运动产生影响。含植被水体的泥沙运动特性研究主要聚焦于水生植被对泥沙侵蚀以及沉降的影响。 单向明渠流条件下,水生植被的存在虽然减小了冠层内的时均流速,但植株存在可能导致尾流紊动产生,进而影响泥沙侵蚀/沉降过程。Liu和Nepf[21]指出:当冠层内尾流紊动存在时(Red>120),冠层内泥沙沉积量减少,床底泥沙发生侵蚀;相反,当冠层内无尾流紊动产生(Red<120)时,冠层内泥沙沉积量增加。Follett和Nepf[42]发现冠层内尾流紊动的大小与泥沙净侵蚀量成正比。由于尾流紊动的存在而导致冠层内泥沙发生侵蚀的现象也为其他实验室研究[43- 44,69]以及野外研究[45]所发现。 传统而言,无植被河道中泥沙起动的临界流速可通过床面剪切应力(τ)进行表征[70]。然而,植生河道中植被的存在导致尾流紊动的产生,基于床面剪切应力对冠层内泥沙的起动与输移进行预测往往产生较大偏差[71- 74]。近些年,一些研究指出,与τ相比,近床底水体紊动能(TKE)能够对床底泥沙运动进行更好的预测[72- 74]。例如,Yang等[72]以及Yang和Nepf[73]指出,随着植被密度的增加,冠层内尾流紊动增加,导致冠层内泥沙起动的临界流速逐渐减小,床面输沙率逐渐增大。通过综合考虑床面以及植被产生的紊动,Yang等[72]以及Yang和Nepf[73]提出近床面水体总紊动能的计算方法,且新的紊动能能够对无植被以及有植被条件下床底泥沙起动的临界流速以及推移质输沙率进行较好的预测。Shan等[75]研究了河道中植被的斑块分布对推移质输沙率的影响,指出河道平均输沙率随河道中植株总数以及斑块分布直径的减小而减小,且通过建立河道床底平均紊动能模型,对床面平均输沙率进行了预测。 植被的存在不仅改变了冠层内部泥沙的沉降与再悬浮格局,也影响着斑块周边区域泥沙的运动状况。正如1.1.2节所述,当水生植被成斑块状分布时,斑块两侧水体流速增加,局部泥沙发生侵蚀[45- 48];而在紧靠斑块下游区域的“遮蔽区”(图3),水体时均流速与紊动能均减小,泥沙沉积量增加[42- 43]。由于泥沙中含有丰富的营养物质,十分有利于植物生长,植被斑块后方泥沙沉积量的增加可促进植被斑块沿水流方向扩展。实际上,一些野外研究[76- 79]也发现了这种特征,即天然条件下植被主要沿水流方向呈流线形向下游延伸。 波浪作用下,植物茎秆周围的尾流紊动也影响着冠层内泥沙的再悬浮过程。Tinoco和Coco[80]采用刚性模型植被进行水槽实验,指出植物与波浪相互作用下产生的尾流紊动是冠层内泥沙再悬浮的主要驱动因素,与无植被条件下相比,尾流紊动的产生可增加冠层内泥沙再悬浮量。Tang等[81]指出,由于尾流紊动的存在,植被冠层内悬浮泥沙起动的临界流速随着植株密度的增加而逐渐减小。然而,大量野外实验却得出相反的结果,即水生植被的存在可有效降低床底泥沙再悬浮量。例如,Zhu等[82]以太湖为研究区,发现与无植被区相比,水生植物可使床底泥沙再悬浮量降低84%;Gacia和Duarte[83]以及Granata等[84]也指出,与无植被条件下相比,海草冠层的存在可使泥沙再悬浮量降低24%~70%。这2种相反结果反映了植被在2种尺度上的影响:① 冠层尺度,植被与波浪的相互作用可减弱波能[50,53];② 植株尺度,植被与波浪的相互作用可产生尾流紊动[68,80]。野外条件下,植被区通常长达几十米至几百米,在这种尺度下,植被可大大降低波能,从而减小冠层内泥沙再悬浮量;然而,在相同波浪条件下的实验室水槽中,植被区长度通常为几米,这种尺度下,植被对波能的减弱作用较小,使得冠层内尾流紊动居主导地位,从而增加植被内泥沙再悬浮量。此外,Zhang和Nepf[85]通过水槽实验发现,水生植被可通过影响床底沙波的形成对悬浮泥沙运动产生影响:当Aw 含植被水流研究已取得巨大进展,尤其是近20年,国内外大量研究揭示了单向明渠流条件下植被内水体的紊动特征,为今后研究植被内部的物质传输奠定了基础;随着紊流研究的开展,水生植被对泥沙运动的影响机制也逐渐被揭示。通过对国内外已有研究进行综述,今后含植被水沙运动研究应关注以下问题: (1) 波浪条件下植被对水体紊动结构的影响。水生植被与波浪的相互作用使得冠层内部出现波生流,进而影响冠层内的物质输移。然而,冠层内波生流大小的定量预测(其与波浪参数、植被密度、形态以及柔性等的关系)以及波生流对冠层内紊动尺度与强度的影响,目前国内外研究较少。 (2) 水生植被对泥沙运动影响的定量研究。床底泥沙运动不仅与近床面的水体紊动有关,还受泥沙粒径影响。因此,含植被水流中泥沙运动(如起动的临界流速、推移质输沙率、泥沙沉降等)的预测,还应纳入泥沙粒径因素,以提高预测的适用性。此外,波浪作用下床底泥沙再悬浮量可尝试通过近床底紊动能进行表征,并结合现有的紊动能模型,建立泥沙再悬浮量与植被参数、水流参数以及泥沙粒径间的定量关系,研究可为浅水湖泊底泥营养盐释放的定量预测奠定基础。 (3) 冠层尺度下植被对水沙运动影响的定量研究。目前,含植被水沙运动研究大多从小尺度水动力学角度出发,研究植被- 水流- 泥沙相互作用机制。今后应多从冠层尺度出发,充分考虑植被区水流结构的空间差异,定量研究植被区对水沙运动的影响,如植被覆盖条件下行洪流量的计算、洪水过程中洲滩湿地的冲淤格局以及泥沙淤积厚度的预测等。

1.2 波浪条件
2 水生植被对泥沙运动的影响研究
2.1 单向明渠流条件
2.2 波浪条件
3 结论及展望
