基于MODIS时序数据北回归线(云南段)地区植被物候时空变化及其对气候响应分析
2021-04-12张艳可王金亮农兰萍程峰张云峰
张艳可 ,王金亮 *,农兰萍 ,程峰 ,张云峰
1. 云南师范大学地理学部,云南 昆明 650500;2. 云南省高校资源与环境遥感重点实验室,云南 昆明 650500;3. 云南省地理空间信息技术工程技术研究中心,云南 昆明 650500;4. 云南师范大学生命科学学院,云南 昆明 650500
植被物候是指植物受气候和其它因子影响而形成的以年为周期重复出现的自然变化现象,包括发芽、抽枝、展叶、开花、结果、落叶和休眠等(竺可桢等,19991-4;郑景云等,2002;王连喜等,2010)。它不仅指示植被的节律变化,也能反映植被对气候变化的响应,是全球变化的“积分仪”和景观生态环境变化的综合“指示器”(Li et al.,2003)。IPCC第五次评估报告指出,1880—2012年全球地表平均温度上升0.85 ℃,全球变暖毋庸置疑(贾文雄等,2016)。气候环境改变会直接影响植被物候变化,植被物候变化又会直接影响着陆地碳、养分动态平衡和生物多样性格局,而后反过来向气候系统反馈(Wang et al.,2017)。
传统物候学通过人力定时定点观察记录植被生长变化信息,精度高,但观测面积小。随着遥感技术发展,遥感数据为地面定点观测提供了数据补充和支持,使得物候观测对象实现了由点到面的空间转换(White et al.,1997;Jan et al.,2010;李晓东等,2017),借助遥感数据进行植被物候监测也成为新的研究方式,但它与传统意义上的物候观测有所不同,它主要获取的是植被物候变化的关键信息,如生长季始期、生长季末期、生长季长度。遥感物候监测确切来说该方法是借助遥感影像数据中的植被指数变化来确定出物候期,目前用到的植被指数主要有NDVI和EVI两种,它们都能反映植被的生长变化情况,EVI指数是在NDVI指数的基础发展而来的增强型植被指数,它解决了NDVI植被指数中存在的易饱和的问题,同时改进了算法和合成方法,减少了异常值的影响(李文梅等,2010)。因此研究选取MODIS EVI植被指数数据进行物候监测研究。
全球变暖背景下已有不同学者利用遥感数据源对不同地区的物候信息进行观测分析(李明等,2011;浮媛媛等,2016;张晓东等,2018)。在全球物候变化研究中,1982—1999年间(Tucker et al.,2001)全球 35°N以上地区的植被生长季有明显的延长趋势;1982—2009年间(丛楠等,2016)北半球欧洲、亚洲、北美地区呈现植被春季萌发期提前趋势,欧亚大陆提前速率为 (0.22±0.04) d·a-1,显著高于北美大陆的变化速率 (0.03±0.02) d·a-1,且该变化主要受气温因子的影响。在我国植被物候变化研究中,东北地区(俎佳星等,2016)1982—2006年不同植被类型春季物候均表现出先提前后推迟的现象,秋季则较为复杂,其中春季物候主要受温度影响,秋季则主要受降水控制。陕西省(韩红珠等,2018)2001—2016年植被 SOS变化趋势为波动提前(-0.79 d·a-1),生长季结束时间变化趋势为波动推迟(0.5 d·a-1),生长季长度变化呈波动延长趋势(1.29 d·a-1)。青藏高原(常清等,2014)高寒草地在2003—2012近10 a来随着春季温度提高,总体呈现生长季开始期提前的趋势(-0.248 d·a-1)。2001—2016年秦岭地区(李登科等,2020)植被物候呈现SOS提前(0.61 d·(10 a)-1)、EOS推迟(3.1 d·(10 a)-1)、LOS 延长(3.7 d·(10 a)-1)的变化特征,3月份气温升高会抑制植被生长期到来、9月气温升高会延迟生长结束期到来。但 2001—2016年新疆地区(何宝忠等,2018)植被物候却出现了SOS推迟趋势(1.9 d·(10 a)-1),EOS呈提前趋势(3.66 d·(10 a)-1),LOS 缩短趋势(5.6 d·(10 a)-1),且 SOS主要受春季气温、前一年冬季降水量和日照时数的影响,EOS主要受夏季和秋季降水量影响。由以上研究可以看出,植被物候变化存在明显的区域性差异,不同地区、不同时间段变化趋势不同,各地区主导变化的气候因子也各不相同。因此深入研究区域尺度植被物候变化及其对气候因子的响应对于理解一地区生态系统内循环和气候—生物圈的相互作用有显著意义(李登科等,2020)。
我国对植被物候变化的研究明显集中在中东部地区以及北部地区,极少对西南部云贵高原做研究。云南省地处中国西南边疆,青藏高原的东部,南部有北回归线穿过,北回归线是北半球热带和温带的分界线,地理位置特殊。研究以北回归线在云南省穿过的县域为研究区,利用长时间序列(2001—2019年)MODIS EVI植被指数提取北回归线(云南段)地区主要物候值信息,包括生长季始期(SOS)、生长季末期(EOS)、生长季长度(LOS),分析研究区整体和不同植被类型的物候值变化,并结合气温、降水资料分析不同植被生长阶段对水热因子的响应,揭示气候变化对该区域植被生长状况的影响,以期为研究区生态环境的保护和持续利用提供科学依据。
1 研究区与数据来源
1.1 研究区概况
北回归线又名夏至线,位于23°26′21″N,是太阳在北半球能够直射到离赤道最远的位置。研究以北回归线在云南省穿过的县市作为研究区(简称北回归线(云南段))(图1),该地区跨越的经纬度为22°48′54″—24°10′44″N,98°48′40″—106°11′39″E,总面积达57129.27 km2,纬度低,经度跨度广。受来自印度洋西南季风的影响,气候主要为高原亚热带季风气候。地处云贵高原,垂直气候明显,从低海拔到高海拔,形成了不同的自然景观。特殊的地理位置和海拔高度形成了独具特色的植被生长情况,对该地区进行物候研究可深入不同自然环境下植被物候值变化状况。
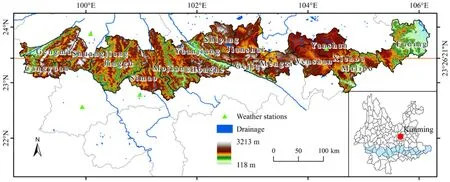
图1 研究区示意图Fig. 1 Sketch map of the research area
1.2 数据来源和处理
MODIS数据:来源于美国国家宇航局(NASA)官网(http://ladsweb.nascom.nasa.gov/data/search.html)。幅号为 H27、V06;(1)2001—2019年增强型植被指数(EVI)——MOD13Q1数据集,19 a间共437期遥感数据,空间分辨率250 m,时间分辨率 16 d;该数据产品已采用最大合成法进行纠正,并已经过了几何纠正和大气校正。同时为保证数据精度,剔除研究区内所有EVI值小于0的区域。(2)2001—2018年MODIS Terra+Aqua3级土地覆盖类型产品——MCD12Q1数据集,空间分辨率为 500 m,为使其和EVI数据分辨率一致,将其重采样为250 m,文中分类方案采用国际地圈生物圈计划(IGBP)的全球植被分类方案。研究区包括的植被类型主要有7种:分别为常绿针叶林、常绿阔叶林、混交林、木本热带稀疏草原、热带稀树草原、草原、耕地。(3)2001—2019年地表温度(LST)数据——MOD11A2数据集,空间分辨率为250 m,时间分辨率为8 d,先利用最大值合成法将每年46期数据合成为12期月均数据。研究区山地地区LST值在每年6、7月存在异常值,其余月份正常,由于6、7月气温差异不大,为提高数据精度,将这两个月数据利用最大值合成法合成为一个月的数据参与相关性分析。
降水数据包括两种,(1)TRMM数据来自于美国国家宇航局(NASA)的Goddard 数据分发中心(https://www.nasa.gov/connect/apps.html)提供,为TRMM 3B43月降雨数据,空间分辨率为0.25°×0.25°,时间范围为 2001—2019 年 1—12月,利用地理加权回归模型建立 TRMM 原始数据和NDVI的关系,并将TRMM 3B43降尺度为1 km,为适用于后续分析,将其重采样为和EVI数据相同的250 m空间分辨率。该数据用于分析19 a间降水和物候参数的空间相关性。(2)气象站点数据:选取耿马、临沧、澜沧、思茅、元江、砚山 6个站点 2001—2019年的月均数据,数据来源于中国气象科学数据共享服务网(http://data.cma.cn//),该数据用于计算气候因子在19 a内的变化状况。
2 研究方法
2.1 时间序列重构方法
MODIS数据受传感器性能、天气等随机因素的干扰会存在一些噪声(刘亚,2017),需要对其进行噪声平滑处理。目前常用的去除时间序列中噪声的方法有S-G(Savitzky-Golay)滤波法、非对称高斯函数 A-G(Asymmetrical-Gaussian)滤波、D-L(Double-Logistic)曲线拟合等。结合前人研究(王乾坤等,2015),并利用3种方法分别进行提取实验进行对比分析,发现S-G滤波法对北回归线(云南段)地区时序数据重构的保真性最好(图2),提取的物候值最符合当地实际物候期。S-G滤波法是1964年由Savitzky和Golay提出的一种最小二乘卷积拟合平滑滤波方法,公式为:
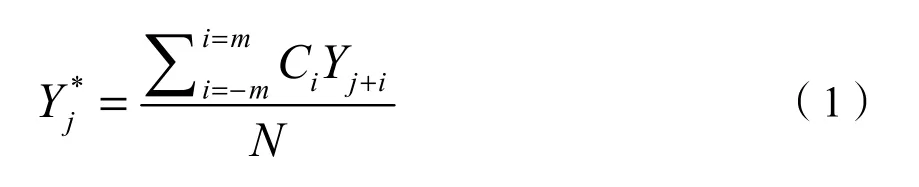
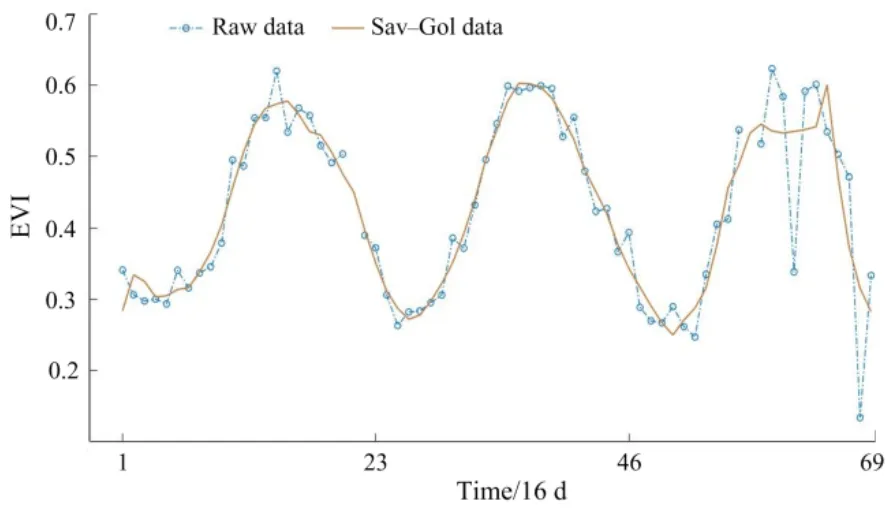
图2 EVI数据平滑前后数据对比Fig. 2 Comparision between before and after smoothing EVI data
式中,Y为原始的EVI值,Y*为滤波拟合后的EVI值,j为数据序列的第j个点,Ci为第i个数据点的滤波系数。N为滑动窗口的宽度即2 m+1(Chen et al.,2004;侯学会等,2013)。
2.2 物候期确定
2.2.1 物候期确定方法
基于植被指数时序曲线提取遥感物候的方法有阈值法、导数法、最大斜率法、模型拟合法等(Chen et al.,2004)。考虑到研究区地形复杂、经度跨度大,采用动态阈值法进行物候参数提取。该方法是Jonsson和Eklundh(2002)提出的物候参数值提取方法,阈值确定采用一种动态的比值形式而不是某一确定值(张延兵等,2015),以植被指数生长曲线左右振幅的 20%为植被生长的起始期和结束期,两点之间的时间长度为生长季长度。研究在宏观上把握植被物候期及时间变化趋势,所以仅考虑一年只有一个生长期的情况(对于有两个生长期的地区,提取该地区植被生长期中EVI值最大的一个生长期)。文中的日期采用儒略日计算法,即将每年的1月1日记为第一天(韩红珠等,2018)。物候值计算公式如下:
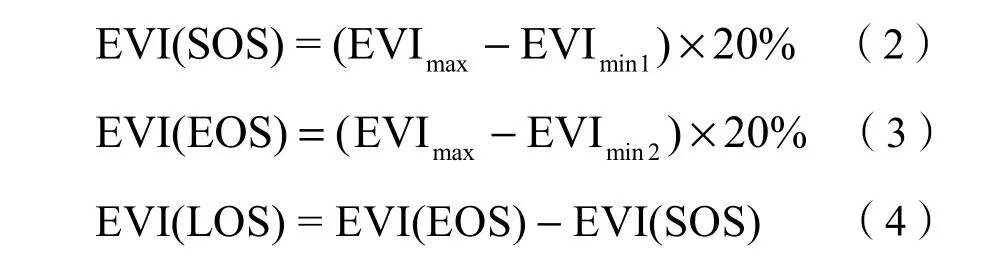
式(2)中,EVI(SOS)表示植被生长季开始期阈值,EVImax表示植被指数最大值,EVImin1表示EVI曲线上升阶段植被指数最小值;式(3)中,EVI(EOS)表示提取EOS的植被指数阈值,EVImin2表示EVI曲线下降阶段植被指数最小值,当动态阈值达到最大与最小EVI差值的20%时,提取出SOS和EOS值。两者在水平方向上的时间差即为EVI(LOS)生长季长度值。
2.2.2 植被物候提取结果精度验证
由于研究区域内没有物候观测点,借助农作物观测点数据进行精度验证。在中国气象数据网获取北回归线(云南段)县区中5个农业气象站点(耿马、临沧、思茅、蒙自、文山)的农作物生长发育数据,选取一季稻的返青时间为对比时间,计算各个站点 2001—2008年一季稻返青期的平均值,以及对应县区栽培植被遥感物候值的平均值,此处栽培植被参考GlobeLand 30数据的划分,将两结果进行比对。耿马、临沧、思茅、蒙自、文山的实测值分别 138、137、113、136、143 d,遥感估算值为 144、144、112、112、144 d,两组数据的相关性指数为0.693(P<0.05)。5个站点的实测值和遥感估算值相差的平均绝对值为2.2 d。考虑到遥感影像的时间分辨率(16 d),这一误差在可接受的范围内。
2.3 一元线性回归分析
一元线性回归分析可以模拟每个栅格的物候要素时间上的变化趋势,文中采用回归方程的斜率来反映北回归线(云南段)地区植被物候值的变化特征,计算公式为:
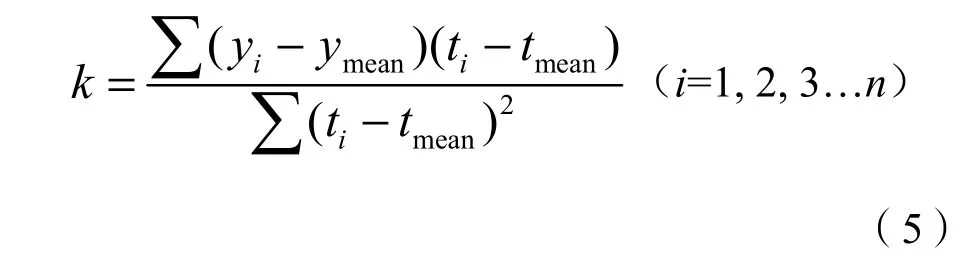
式(5)中,yi为植被物候在第i年对应的生长季始期、生长季末期和生长季长度,ymean为该物候值的多年均值,ti为对应年数,tmean为年数的平均值,k为趋势线的斜率。若k>0,说明物候期的变化趋势是延长或延迟,若k<0,则说明物候期的变化趋势是提前或缩短。
2.4 植被物候期与水热因子关系分析
利用Pearson相关系数来分析北回归线(云南段)地区植被物候的生长季开始期、结束期和生长季长度与气温、降水的相关性,计算公式为:
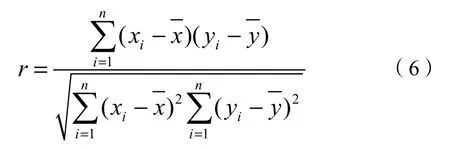
式(6)中,x为气温或降水,y为植被物候值,n为数据组数,i为序列数,相关系数r取值在-1—1之间,当r>0表示x和y为正相关关系,当r<0表示x和y为负相关关系,绝对值越大表示相关性越强。
3 结果与分析
3.1 多年平均物候空间分布
从 2001—2019年植被生长季始期空间分布来看(图3a),SOS大部分集中分布在80—112 d,但区域差异明显。植被生长最早开始地区为东部富宁、麻粟坡、西畴县地区和西部山麓迎风坡(西南坡)地区,约在48—96 d;其次为西部河谷地区,在105—138 d,中东部坝区(砚山、蒙自、建水、石屏)植被生长开始期约在105—150 d。由此可见生长季开始期受海拔和地势起伏制约明显,在地势起伏小且海拔较低的地区,水热充足且蒸发弱,植被SOS最早,反之则较晚。除此之外,研究区SOS还存在一特殊现象,山地地区 SOS开始于秋季(192—240 d),这明显不符合一般情况,原因在于研究区山地地区植被EVI值在夏季存在“逆季”现象,7月EVI值为一年中最低,夏季植被覆盖要低于秋冬季节,这与云南澜沧江流域(李立科,2011)的研究结果和云南西双版纳(赵桔超等,2019)的研究结果一致。
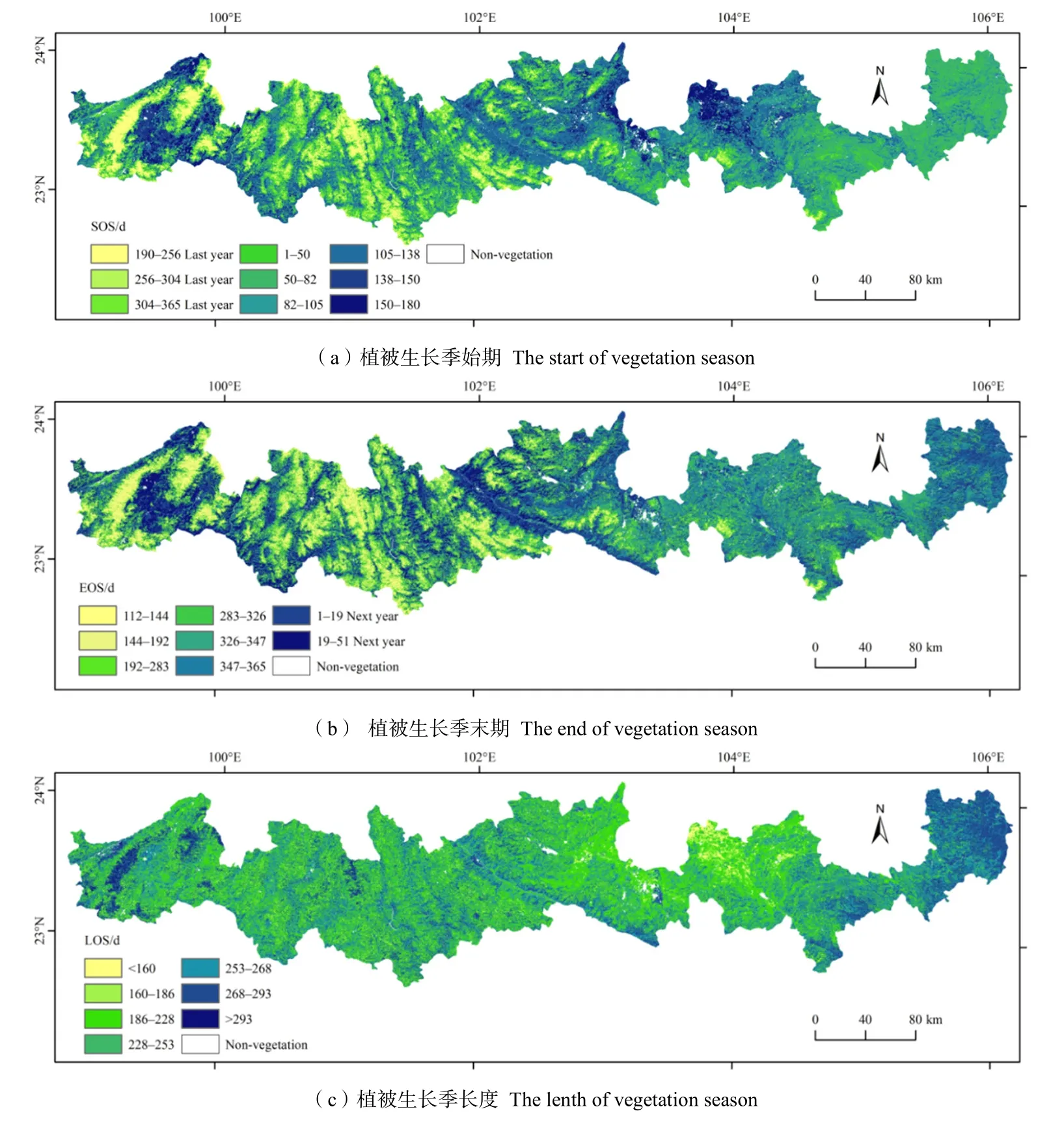
图3 北回归线(云南段)地区植被物候参数分布图Fig. 3 Phenological parameters of the tropic of cancer (Yunnan section)
从 2001—2019年植被生长结束期空间分布来看(图3b),EOS大部分集中在337至次年17 d,结束最晚的西部河谷地区约在次年的17—51 d,其次为东部低海拔地区(富宁、麻粟坡、西畴),约在353至次年17 d,稍早的中东部坝区(砚山、蒙自、建水、石屏)约在320—353 d,由此可见,除山地地区,其余地区 EOS虽存在一定差异,但差异不大,基本都在12月至次年1月。EOS同样也受海拔和地势起伏的制约,河谷地区在冬季存在一定积温作用,结束最晚,东部地区次之,中东部坝区地势平坦,植被类型多为农作物,植被结束生长稍早。山地地区EOS出现在5—6月左右(112—192 d),原因在于在5—6月降水还未到达,山地海拔高,升温快,气温升高加剧干旱,导致植被严重缺水,抑制植被生长。
植被生长季长度大部分集中在224—272 d(图3c)。LOS最长的出现在山地迎风坡(西南坡)地区和东部低海拔区,约为273—305 d,其次为河谷地区,约为257—273 d,山地背风坡地区为200—220 d左右,中东部坝区植被类型主要为耕地作物,生长期最短,约为161—209 d。由此可见植被生长受水热因子影响明显,在降水较多的迎风坡和积温较好的低海拔和河谷地区植被生长时间较长,背风坡和高海拔地区降水量少、蒸发强且积温效果较差,植被生长长度较短。
3.2 不同植被类型物候空间分布特征
从表1来看,在主要的3种植被类型中,森林植被开始生长最早、结束最早、长度最长;草原植被较森林植被开始生长较晚,但结束生长最晚,生长期长度比耕地植被长,比森林植被短;耕地植被开始生长最晚,生长期也最短。不同植被类型生长物候值差异主要和分布地区有关,3种森林类型的植被包括常绿针叶林、阔叶林、混交林分别分布在山顶地区、山腰地区和山麓地区,从山顶到山麓,海拔越高,植被夏季“逆季”现象越明显,跨年生长现象越明显。3种森林植被物候值特点为:植被存在跨年生长,海拔越高,物候值跨年现象越明显,植被开始生长越早,结束越早、生长期越长。3种草原类型包括木本热带稀树草原、热带稀树草原、草原分别分布西部河谷和东部低海拔区(富宁县东部)、中东高原山地和坝子地区、坝子地区。草原类型的植被主要分布在中东高原区,地势相对平坦,3种草原植被类型的特点为:海拔越低,植被开始生长越早、结束越晚、生长期越长。耕地和草原物候值类型相似,两者在空间分布在也相邻,都主要分布在坝子地区,耕地受人为因素影响较大,所以与自然植被相比开始较晚、结束较早、生长期较短。
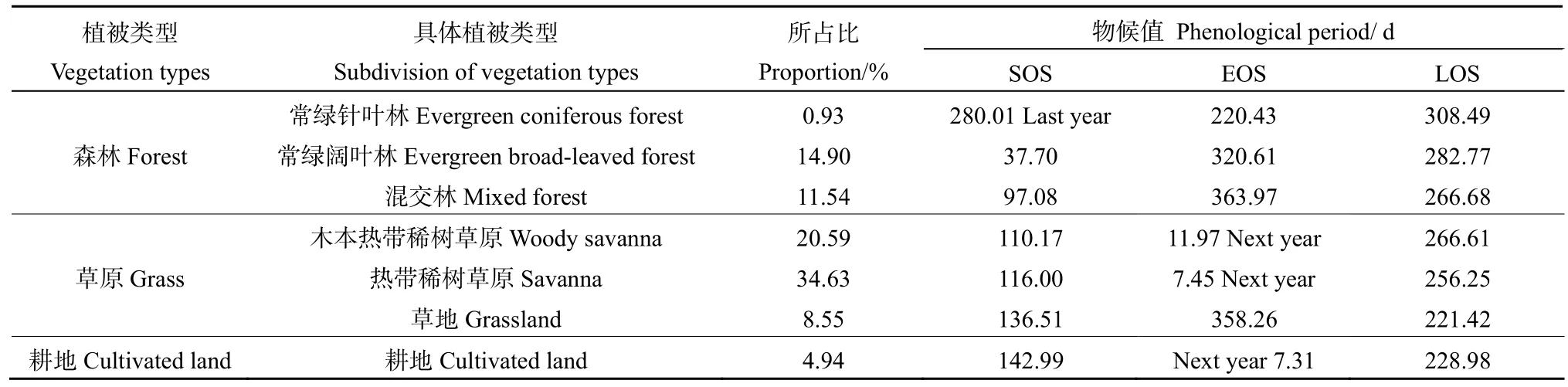
表1 不同植被类型物候期和所占比Table 1 Phenological period and their proportions of different vegetation types
3.3 物候年际变化特征
3.3.1 物候年际变化总特征
图4所示为计算得到的2001—2019年北回归线(云南段)地区植被物候年际变化特征。SOS具有提前趋势,且通过了显著性检验(R2=0.51,P=0.001<0.05),平均提早 1.14 d·(10 a)-1。这一趋势与全球 SOS物候值变化趋势一致,但比全球1981—2003 年的-3 d·(10 a)-1(Julien et al.,2009)、北半球 2000—2008 年间的-(4±0.8)d·(10 a)-1(Jeong et al.,2011)的幅度小。EOS呈推迟趋势,平均推迟了 0.07 d·(10 a)-1,未通过显著性检验(R2=0.01,P=0.756>0.05)。变化幅度也比全球1981—2003 年间的 0.45 d·(10 a)-1(Julien et al.,2009)和北半球 2000—2008年间的 2.3 d·(10 a)-1小(Jeong et al.,2011)。研究区LOS在19 a中为延长趋势,且通过了显著性检验(R2=0.47,P=0.001<0.05),平均延长 1.09 d·(10 a)-1,这一趋势与全球及北半球地区LOS延长趋势一致(Julien et al.,2009;Jeong et al.,2011)。
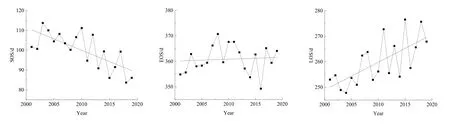
图4 2001—2019年北回归线(云南段)地区植被物候年际变化趋势Fig. 4 Interannual phenological change trend of vegetation in the tropic of cancer (Yunnan section) region from 2001 to 2019
3.3.2 物候年际变化趋势空间分布
根据年变化率k和相关系数r值的大小,将北回归线(云南段)地区19 a的遥感物候年际变化趋势分为3种情况:(1)当k<0且r<0.3时,显著提前;(2)当k>0 且r<0.3 时,显著推迟;(3)当r>0.3时,无显著变化。从图5a来看,北回归线(云南段)地区SOS显著提前地区占比为49.13%,集中在研究区西部山地地区。显著推迟地区占比为22.88%,主要分布在西部河谷地区和背风坡山麓地带。无显著变化地区主要分布在较为平坦的河谷平原和中东高原坝子地区,占比为27.99%。EOS变化趋势比SOS变化稍弱(图5b),有45.35%地区的EOS无显著变化,无显著变化区分布在东部低海拔地区、河谷平原和中东高原坝区。显著提前的地区减至39.61 %,分布在海拔较高的山地地区。显著推迟的地区减至15.04%,主要分布在西部山地地区。植被LOS有近一半的区域变化趋势为延长(图5c),占比为41.47%,58.42%的区域无显著变化,显著缩短区域仅占0.12%;显著增加的区域主要分布在海拔较高地区,无显著变化的分布在低海拔地区、河谷平原和坝区。
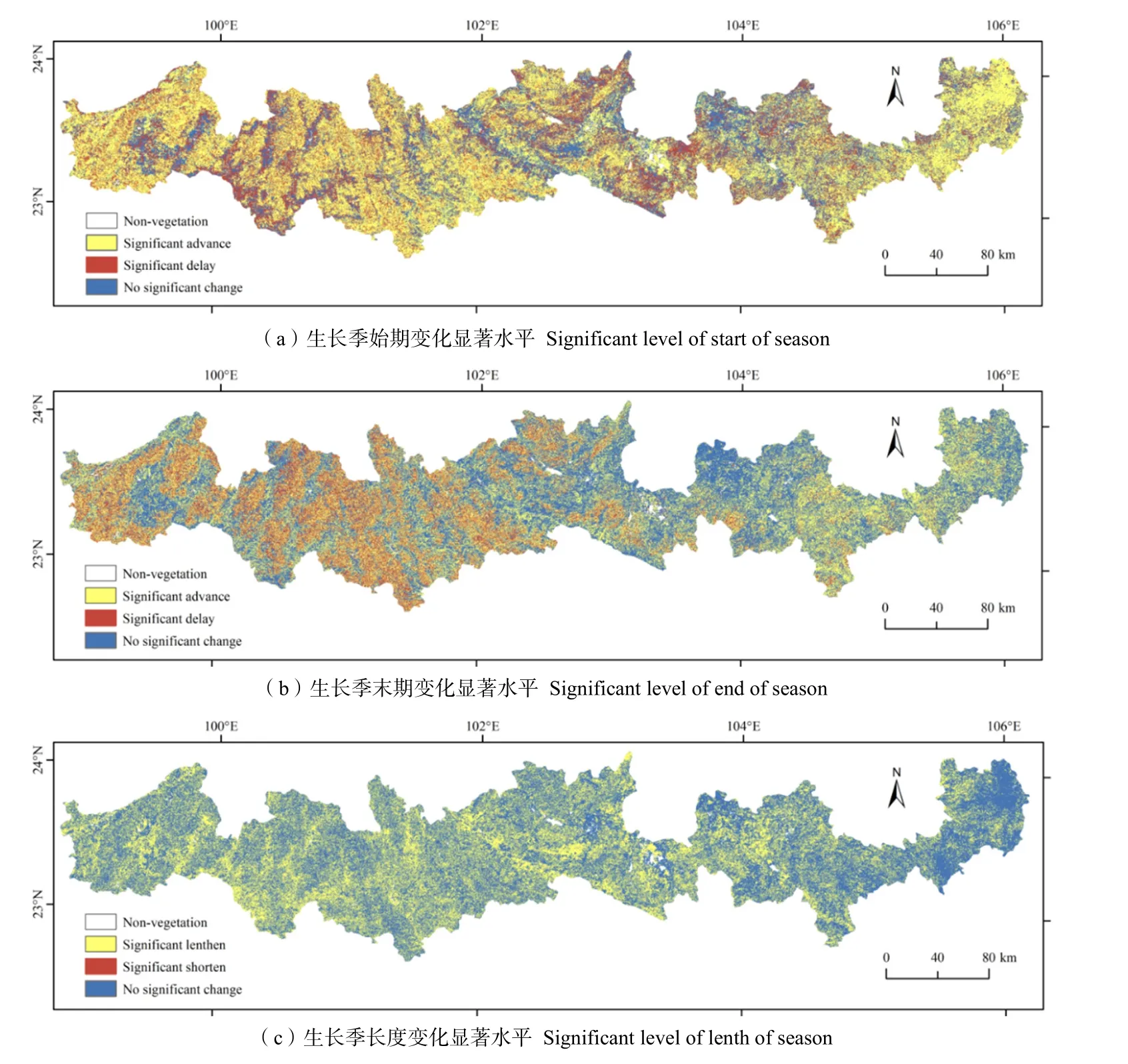
图5 北回归线(云南段)地区2001—2019年植被物候年际变化显著水平Fig. 5 The significant level of interannual phenological change of vegetation in the tropic of cancer (Yunnan section) from 2001 to 2019
3.3.3 物候年际变化速率空间分布
研究区大多数区域植被SOS变化速率在-4—2 d·(10 a)-1,占全区89.9%(图6a)。其中变化速率在-4—0 d·(10 a)-1之间的区域占30.47%,分布在西部山区;0—2 d·(10 a)-1之间的区域占比为8.3%,分布在西部河谷和山麓地区;变化速率为0的占比最大,为51.1%,分布在低海拔区、河谷地区和坝子地区。变化速率小于-5 d·(10 a)-1的区域占比为6.09%,主要分布在西部高山地区的迎风坡地带;变化速率大于2 d·(10 a)-1的区域占比为4.01%,主要分布在海拔稍高的西部山腰地区。
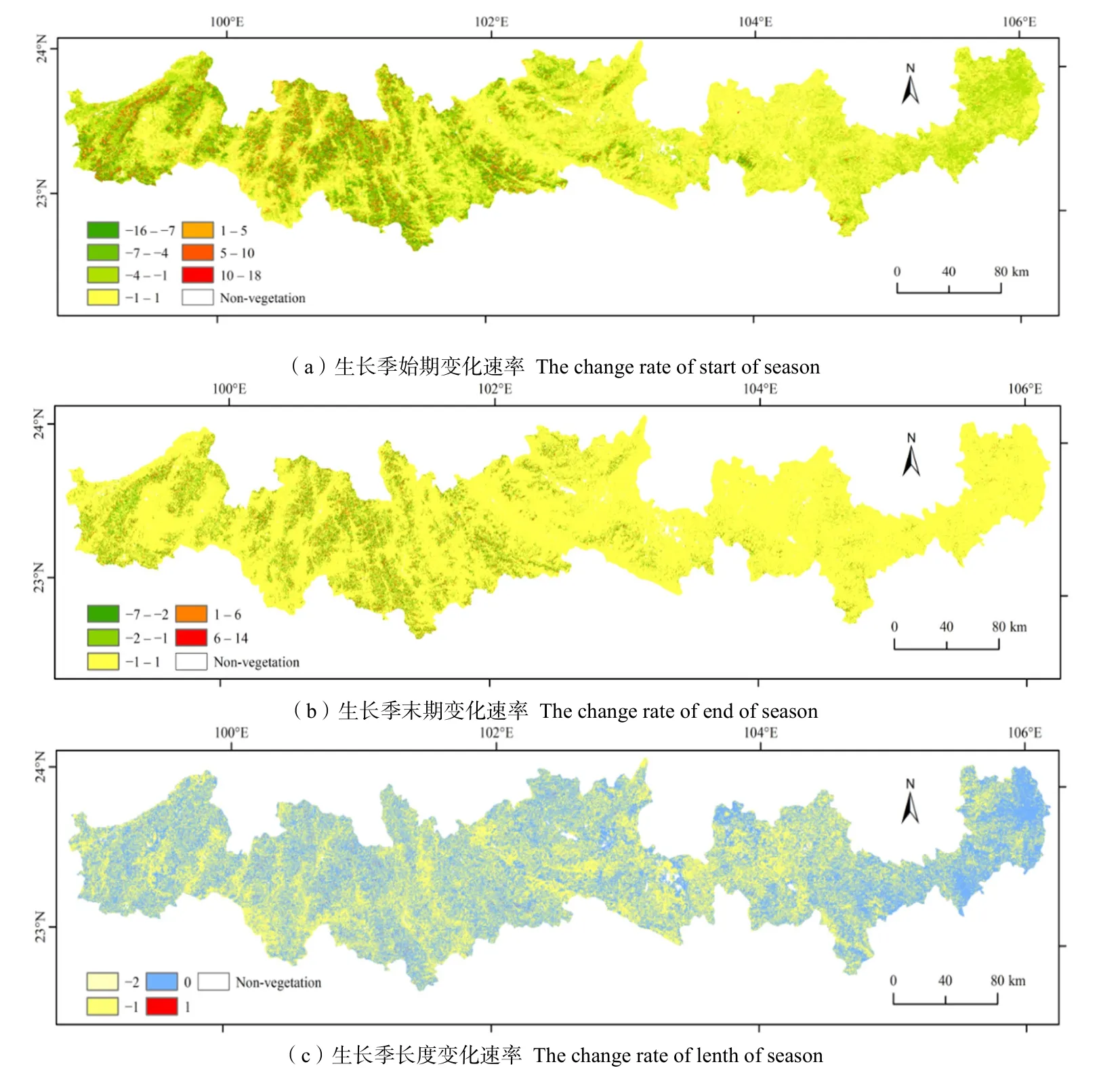
图6 北回归线(云南段)地区2001—2019年植被物候年际变化速率Fig. 6 The rate of interannual phenological change of vegetation in the tropic of cancer (Yunnan section) from 2001 to 2019
植被EOS变化速率与SOS相比明显变弱(图6b),变化速率在-1—1 d·(10 a)-1之间的占比为92.22%,分布在除西部山地之外的各个地区,变化速率小于-1 d·(10 a)-1的占比仅为5.02%,分布在迎风坡山腰和山顶地区;变化速率大于1 d·(10 a)-1的占比为2.76%,分布在西部山腰地区。
植被 LOS变化速率主要分布在-1—0 d·(10 a)-1,该区间占研究区面积的99.80%(图6c),其中变化速率为-1 d·(10 a)-1的占比为50.24%,主要分布在山腰和稍有起伏的河谷地区,变化速率为 0 d·(10 a)-1的占比为49.56%,分布在研究区平坦的河谷、坝子、起伏较小的山地和东部低海拔区。
3.3.4 不同植被类型物候值时间变化趋势
从SOS变化来看(图7),除了草地和耕地2种植被类型为SOS推迟,其余地区均为SOS提前。变化最大的是分布在西部山地的常绿阔叶林,变化绝对值从大到小依次为常绿阔叶林 (-1.68 d·(10 a)-1)>混交林 (-1.04 d·(10 a)-1)>木本热带稀树草原 (-0.79 d·(10 a)-1)>草原 (0.48 d·(10 a)-1)>热带稀树草原(-0.42 d·(10 a)-1)>耕地 (0.41 d·(10 a)-1)>常绿针叶林(0.05 d·(10 a)-1)。从EOS变化来看(图7),3种森林植被表现为EOS提前,3种草原植被和耕地植被表现为 EOS推迟,变化率从大到小依次为耕地 (1.25 d·(10 a)-1)>草原 (1.22 d·(10 a)-1)>常绿阔叶林 (-1.03 d·(10 a)-1)>常绿针叶林 (-0.74 d·(10 a)-1)>木本热带稀树草原 (0.47 d·(10 a)-1)>热带稀树草原 (0.46 d·(10 a)-1)。从LOS变化来看(图7),所有植被类型均表现出生长期长度延长的趋势,草地植被的LOS变化略大于森林植被。变化率从大到小依次为木本热带稀树草原 (1.28 d·(10 a)-1)>热带稀树草原 (0.89 d·(10 a)-1)>耕地 (0.84 d·(10 a)-1)>混交林 (0.83 d·(10 a)-1)>草原 (0.74 d·(10 a)-1)>常绿阔叶林 (0.66 d·(10 a)-1)>常绿针叶林 (0.12 d·(10 a)-1)。
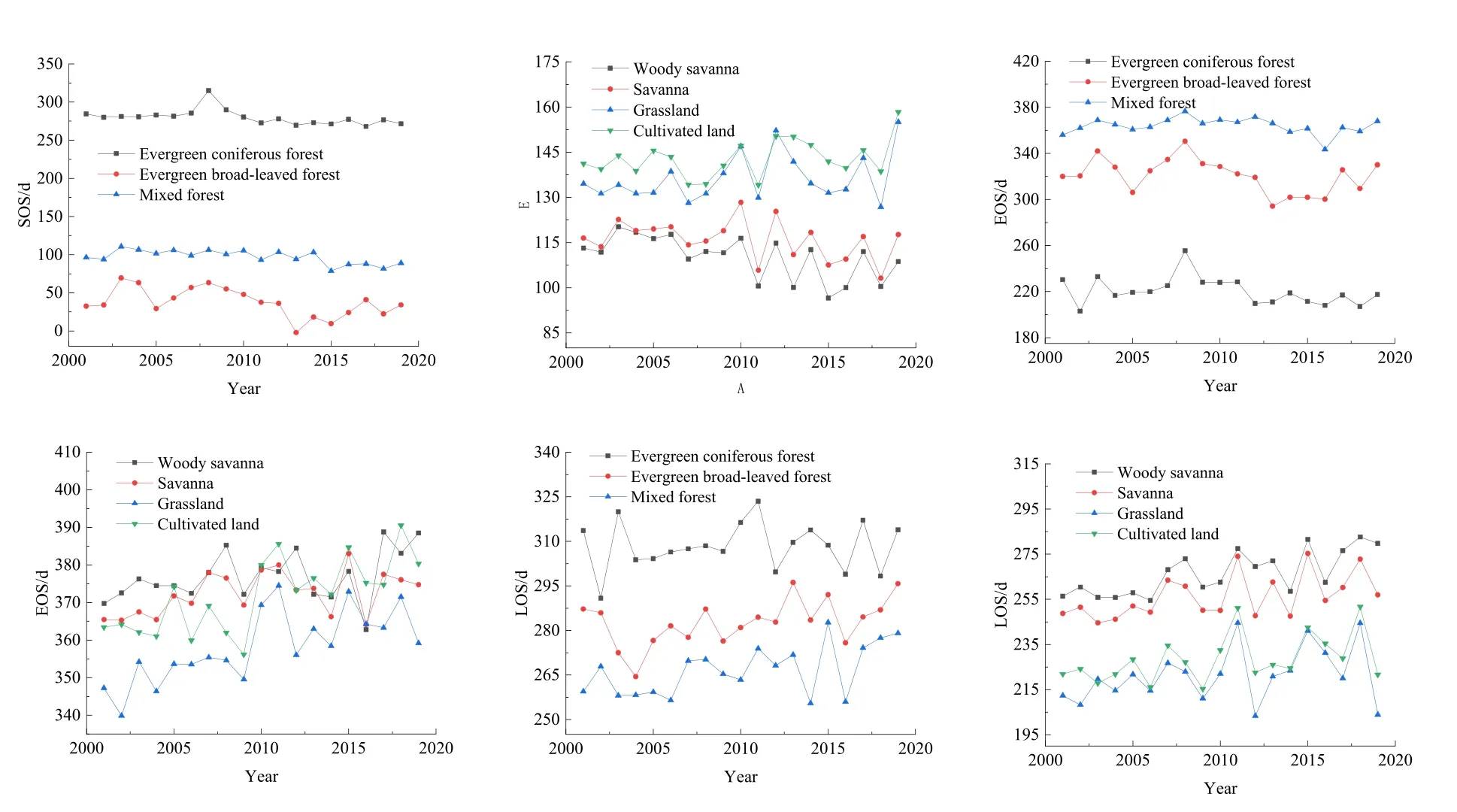
图7 2001—2019年不同植被类型物候值变化趋势Fig. 7 Phenological change trends of different vegetation types from 2001 to 2019
3.4 植被物候期对水热因子的响应
3.4.1 植被物候期对年际气候因子变化的响应
从图8可知,19 a间北回归线(云南段)地区气温呈波动式上升;降水量呈波动下降。从气温变化来看,2004、2008、2011年出现明显下降趋势,对应的物候值变化情况为 2004年的 SOS较上一年出现略微提前趋势,EOS和 LOS变化不大,2008年表现出的SOS提前、EOS推迟、LOS延长,2011年为SOS提前、EOS基本无变化、LOS延长。可见年气温变化对物候值存在响应关系,且与植被生长呈负相关,即年均温升高会抑制植被生长。从降水来看,2003、2012年出现了明显的明显下降趋势,2007、2013年出现明显的上升趋势,对应的物候值变化为2003年SOS推迟、EOS推迟、LOS略微缩短;2012年SOS推迟、EOS提前、LOS缩短;2007年SOS略微提前、EOS推迟、LOS延长;2013年为SOS提前、EOS略微提前、LOS延长。年降水量变化对物候值也存在一定响应关系,且与植被生长呈正相关,即降水量增多会促进植被生长。
从图8来看,近19 a内,月降水量除了2月、5—6月、10月呈减少趋势外,其余月份均呈增加趋势,其中5月减少最多,1月和7月增加最多,雨季来临有推迟趋势,在多数植被开始生长的 3—4月,降水呈增加趋势,有助于植被生长。月气温除了1—4月呈降低趋势外,其余月份呈增加趋势,其中增温最明显的在5月,这一变化不利于山地地区的植被在夏初的生长,而 1—4月的气温降低有助于河谷、坝子、东部低海拔区的植被生长。
3.4.2 植被物候值对单一气候因子的响应
由于北回归线(云南段)地区河谷、坝子、东部低海拔区和山地地区的物候值存在明显差异,分别计算河谷、坝子、东部低海拔区的SOS值与上年10—12月和翌年1—5月气温和降水的相关性、山地SOS值与5—9月气候因子的相关性(图9,文中仅展示部分月份)。在降水方面,河谷、坝子、东部低海拔区10—11月、翌年4—5月的降水和SOS呈正相关,12月至翌年3月呈负相关,相关性系数平均最大值出现在 2月,该月负相关区域占比为61.83%,正相关区域占比38.17%,两者为负相关表明降水越多,SOS来临越早,即降水对植被的生长起促进作用。山地地区5—9月降水和SOS均呈正相关,即降水越多,山地SOS来临越晚,这与山地植被在6—7月的EVI值下降有关,降水多时山地植被生长状况好,EOS来临晚,SOS也会随之推迟。在气温方面,河谷、坝子、东部低海拔区10月至翌年5月气温与SOS均呈正相关,平均最大值出现在4月,该月正相关地区占比为94.96 %,负相关地区仅占5.03%,表明气温升高会抑制植被生长,延迟生长期到来。山地地区5月和9月气温和SOS呈正相关,6—8月呈负相关,相关性系数平均最大值出现在5月,其中负相关地区占比为22.21%,正相关地区占77.79%,即春末夏初气温升高会抑制植被生长,导致植被生长季始期的推迟。
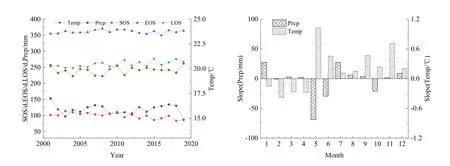
图8 2001—2019主要物候参数和气候因子的年变化及其月变化Fig. 8 Changes in the annual main phenological parameters and climatic factors and the monthly values from 2001 to 2019
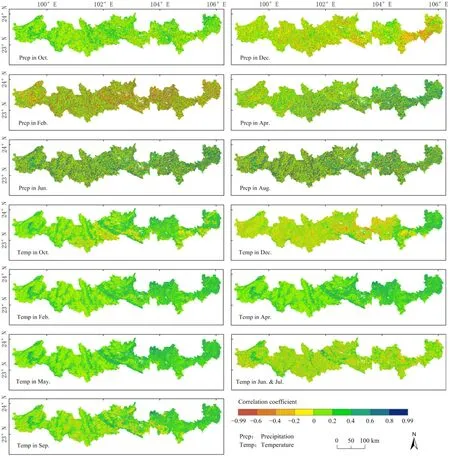
图9 北回归线(云南段)地区植被SOS与各月气象因子的相关系数空间分布Fig. 9 Spatial distribution of correlation coefficients between SOS and each month climatic factors on in the tropic of cancer (Yunnan section)
河谷、坝子、东部低海拔区EOS集中在冬季,山地地区EOS集中在夏初,分别计算9月至翌年1月、1—5月EOS与水热因子的相关性(图10)。在降水方面,河谷、坝子、东部低海拔区9—11月降水与EOS呈正相关,12月至翌年1月呈负相关,其中相关性最大的出现在9月,该月正相关地区占比为59.56 %,负相关地区占40.44%,11月正相关地区达到61.20%,负相关地区为38.80%,表明降水增多会延缓植被生长结束期的到来,降水对植被生长仍起促进作用。山地地区1—3月份降水与EOS呈负相关,4—5月呈正相关,平均最大值出现在5月份,该月正相关地区占比为57.35%,负相关地区占42.65%,表明5月份降水增多会延迟植被生长结束期的到来,对植被生长起促进作用。在气温方面,河谷、坝子、东部低海拔区9—11月气温和EOS呈正相关,12月至翌年1月呈负相关,其中相关性系数平均最大值出现在9月,该月正相关地区占比为75.76%,负相关地区占比为24.23%,这一阶段气温呈下降趋势,表明秋冬季适当增温有利于植被生长。山地地区1—5月气温和EOS均呈正相关,平均最大值出现在 4月,该月正相关地区占比为75.80%,负相关地区占比为24.20%,表明4月气温升高仍对山地地区植被生长起促进作用。
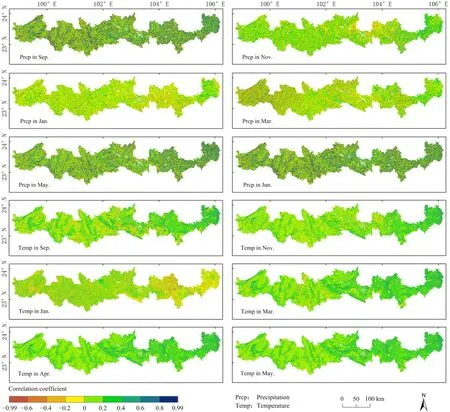
图10 北回归线(云南段)地区植被EOS与各月气象因子的相关系数空间分布Fig. 10 Spatial distribution of correlation coefficients between EOS and each month climatic factors on in the tropic of cancer (Yunnan section)
4 讨论
4.1 植被物候空间分布特殊性
西部山地物候参数与河谷、坝子、东部低平原区明显不同,图11a、b分别为研究区整体和山地地区年内EVI的变化趋势图,可以看出山地EVI值在夏季最低,出现“逆季”现象。原因可能主要在于:研究区本身纬度低,年均温基本在 10 ℃以上,山地与其他地区相比,海拔高,光照强,在夏季气温升高时,蒸发量比低海拔区更强,植被生长由于干旱缺水就会出现“逆季”现象。秋冬季节,山地地区接受的热量比河谷等地区更多,气温更加适合植被生长。而气温升高对植被生长季始期起抑制作用的原因可能主要在于,(1)研究区干湿季分明,降雨主要集中在5—11月,其余时间气温增长不但不会促进植被生长,反而会加剧干旱,抑制植被生长。(2)适当增温可以满足植被生长所需热量供应,但温度过高会改变植物群落的小气候环境,直接或间接地影响植被的生物量生产,对植被覆盖产生抑制作用(Fu et al.,2013;Zhang et al.,2013)。
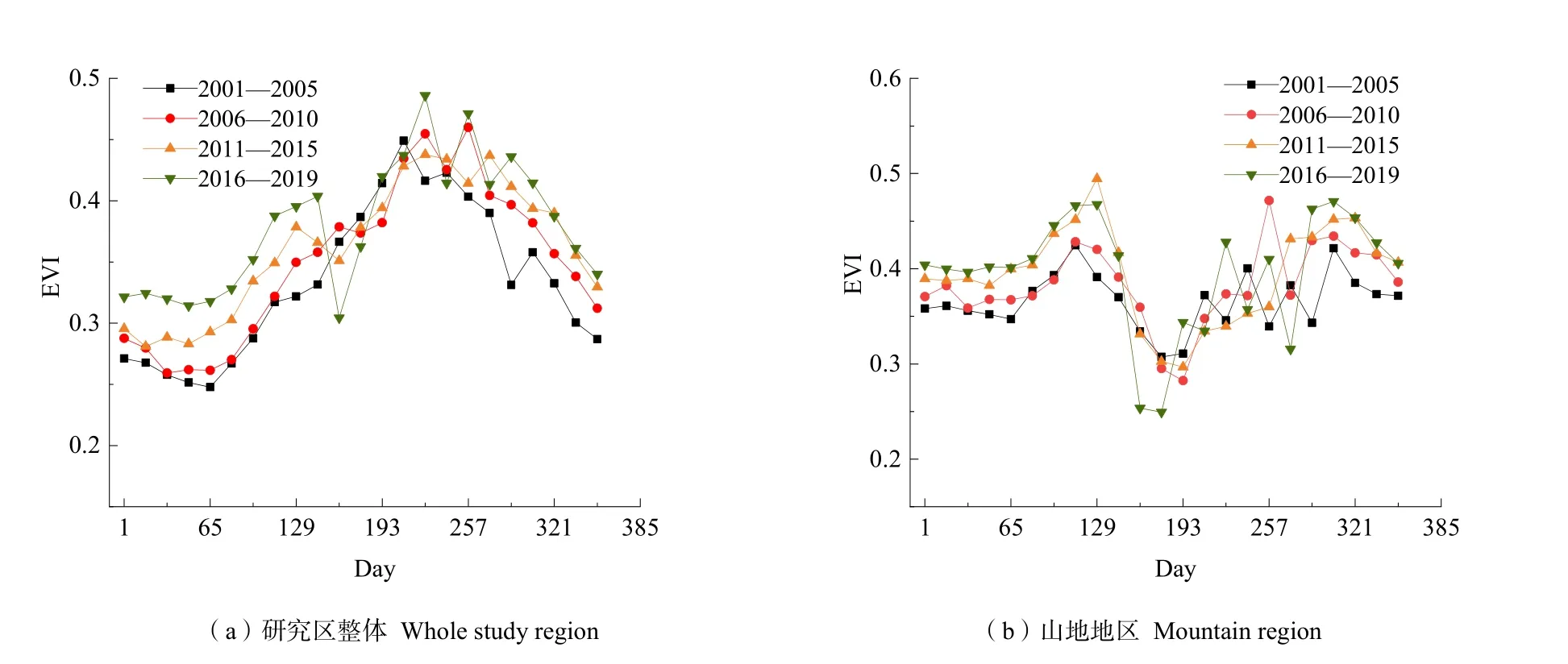
图11 植被EVI值年内变化曲线Fig. 11 Curve of EVI value of vegetation during the year
4.2 植被物候时间变化趋势分析
研究区植被生长始期提前、末期推迟,生长期延长普遍出现在海拔较高地区,反之则出现在海拔较低、地势较平坦的地区,这与气候变暖具有一定关系,高海拔区的植被主要是自然草地和森林植被,物候期变化主要受气候变化的影响。而低海拔区的植被多数是农作物或者人工林等,气候变暖会使得播种期推迟、成熟期提前、生育期缩短。因此从时间变化的空间分布来看,SOS推迟趋势出现在河谷地区,提前趋势出现在山地;EOS提前趋势出现在河谷地区,推迟趋势的出现在山地;LOS延长趋势的出现在山地,无显著变化的出现在河谷、坝子、东部低海拔区。
5 结论
研究基于2001—2019年的MODIS EVI植被指数数据,探讨了北回归线(云南段)地区近20年的植被生长季始期、生长季末期、生长季长度3个物候值的空间分布状况、时间变化特征以及年际和年内尺度下植被生长对水热因子的响应,主要结论如下:
(1)北回归线(云南段)地区海拔和地势起伏在物候地域分异中作用显著,植被物候存在明显的垂直地带性分布特征。山地地区与河谷地区、低海拔区、坝子地区的植被物候值明显不同。山地地区的植被SOS在192—240 d,EOS在次年144—192 d,LOS在272—304 d;河谷地区、低海拔区、坝子地区的植被SOS在80—112 d,EOS在337至次年17 d,LOS 在 224—256 d。
(2)19 a间北回归线(云南段)地区植被物候年际变化的总体特征呈现 SOS具有显著提前趋势(R2=0.51,P=0.001<0.05),平均提前 1.14 d·(10 a)-1;EOS具有推迟趋势(R2=0.01,P=0.756>0.05),平均推迟0.07 d·(10 a)-1;LOS年际变化具有显著延长趋势(R2=0.47,P=0.001<0.05),平均延长 1.07 d·(10 a)-1。
(3)不同植被类型生长状况和变化趋势有一定区别,SOS、EOS、LOS在19 a间变化最大的分别是常绿阔叶林(-1.68 d·(10 a)-1)、耕地(1.25 d·(10 a)-1)、木本热带稀树草原(1.28 d·(10 a)-1)。
(4)水热组合对植被生长影响显著,植被SOS总体与降水呈负相关、与气温呈正相关,EOS与气温和降水均呈正相关。2月降水(负相关)和4月气温(正相关)对河谷、坝子、东部低海拔地区SOS影响最显著;6月降水(正相关)和5月气温(正相关)对山地地区SOS影响最显著。9月的降水和气温(正相关)对河谷、坝子、东部低海拔地区的EOS影响最显著,5月降水(正相关)和4月气温(正相关)对山地地区EOS影响最显著。
目前的MOD13Q1物候产品与其他遥感物候监测产品类似,也存在着数据分辨率不高、噪声干扰、结果验证不充分等问题(范德芹等,2016;张晓萱等,2019),但尽管存在一些限制,基于大范围、长时序的遥感数据仍可从宏观上把控研究区的植被物候的空间分布和时间变化情况,提供关于植被物候的有价值的空间连续的信息。未来植被物候分析的发展需要从遥感数据源质量、数据处理方法、识别结果验证及物候预测等几个方面开展深入研究。
