两种光照下低海拔引种云锦杜鹃的光合作用日变化*
2021-04-12宋绪忠王秀云郑国良
杨 华 宋绪忠 王秀云 沈 剑 郑国良
(1.浙江省林业科学研究院,浙江 杭州 310023;2.金华市林业技术推广站,浙江 金华 321000)
云锦杜鹃Rhododendron fortunei 原产我国,为常绿灌木或小乔木,花期5—6 月,总状花序大如碗,花粉红色或白色带粉,带有淡淡的香气,分布于长江流域各省,在高山上大多呈零星分布[1],也有部分区域成片分布,如浙江天台华顶山、浙江景宁上山头、湖南省城步苗族自治县金童山,具有很高的园林观赏价值。目前,相关文献对云锦杜鹃的形态特征、分布范围、种子形态以及自然条件下生长规律、光合特性、群落特征等进行了描述[2-7],介绍了高山杜鹃引种驯化情况[8]。云锦杜鹃适宜在海拔高、气候凉爽而湿润的环境下生长,而城市夏季常有温室效应,最高气温可达38℃以上,因此引种成功率并不高,对云锦杜鹃人工栽培方面的研究少有报道[9-10]。
光合作用作为植物生长发育的基础,涉及多种物理化学过程,受诸多因素影响。开展植物的光合特性及其与环境生态因子关系的研究,对了解植物生长状况和明确植物对环境的适应性来说,是一种有效的方法。在诸多的影响因子中,光合有效辐射是一个重要生态因子,光合有效辐射过强时植物容易受到灼伤,过弱时无法满足植物发挥最大光合能力。因此,众多学者研究了不同植物的需光性及遮光处理对植物的影响[11-13],以指导植物栽培。夏季晴天高温、强光照时,牡丹Paeonia suffruticosa 叶片光系统Ⅱ(photosystemⅡ, PS Ⅱ)反应中心发生了可逆失活,PS Ⅱ功能下调,发生光合作用的光抑制现象,进行遮荫可消减光抑制,改善光合功能并促进植物生长[11]。潘远智[12]等人发现长期在遮荫环境中生长会导致一品红Euphorbia pulcherrima 光合速率下降,但将其再放入自然光照下,其光合速率不升反降,说明遮荫处理已改变了一品红对光环境的适应。云锦杜鹃多生长在高山或纬度较高的地区,即使在全光照条件下,也因山高气温相对较低、湿度相对较大而正常生长。本研究以引种到低海拔区域——杭州的云锦杜鹃盆栽苗为研究对象,分析10%光照和全光照对其光合作用的影响,以期为人工栽培云锦杜鹃的光环境人工调控提供一定的依据。
1 材料与方法
1.1 材料
3 月,从浙江省松阳县枫坪乡海拔1 000 m左右的高山上,移栽5-6 年生云锦杜鹃野生苗于浙江省林业科学研究院苗圃地 (30°13′09″N,120°01′26″E)。该 苗 圃 地 海 拔 为24 m,年 均 气温达16.2 ℃,年均降水量为1 500 mm。移栽苗高为30~45 cm,地径为0.3~0.5 cm,冠幅为25 cm×15 cm~20 cm×30 cm。栽种基质为酸性黄土,盆大小为20 cm×25 cm(直径×高)。移栽当年适时遮荫、浇水,移栽1 年后,苗木全部成活且生长正常。次年5 月,按生长情况将10 株苗分成2 组,其中5 株放置在近樟树Cinnamomum camphora 树干的树阴下,此树冠幅为10 m×8 m,高为5 m,生长旺盛。用LX1010B 照度计对树干四周多点多次测定树阴下的光合有效辐射,树下光合有效辐射约为自然光照下的10%。另5 株置于空旷地,无任何遮荫设施,认为是全光照条件。在10%光照和全光照两种光照中处理2 个月后,进行光合作用日变化的测定。根据水分亏缺情况及时补水。
1.2 测定方法
1.2.1 光合作用日变化测定 8 月下旬,在晴天6: 00—18: 00 时,使用LI-6400 便携式光合作用测定系统,采用透明标准叶室,选择生长势最接近的3株,每株选3 片最佳功能叶各测3 次,测定净光合速率(Pn/ μmol CO2·m-2·s-1)、蒸腾速率(Tr/ mmol H2O·m-2·s-1)、气 孔 导 度(Gs/ mol H2O·m-2·s-1)、胞间CO2浓度(Ci/ μmol CO2·mol-1),同时记录的指标有光合有效辐射(PAR / μmol·m-2·s-1)、大气CO2浓度(Ca/ μmol CO2·mol-1)、空气温度(Ta/ ℃)、叶片温度(Tl/ ℃)、空气湿度(RH / %)等,每次测定相隔2 h。
1.2.2 数据统计与分析 所有数据取平均值,使用Excel 2003 进行计算和曲线图绘制。
水分利用效率(WUE / μmol CO2•μmol-1H2O)计算公式为:WUE =Pn/ Tr。Pn代表净光合速率,Tr代表蒸腾速率。
气孔限制值(Ls)计算公式:Ls=(Ca-Ci) / Ca。Ca代表大气CO2浓度,Ci代表胞间CO2浓度。
2 结果与分析
2.1 环境因子的日变化
分析各环境因子(图1),光合有效辐射日变化呈“单峰”型,两种光照在中午12:00 时达到最高点。两种光照强度下大气CO2浓度值较接近,不同时间亦无明显变化。随着时间的推移,空气温度不断上升,空气湿度逐渐降低,到14:00时,两种光照下空气温度达到最高点,空气湿度达到最低点,随后温度降低,湿度升高。10%光照下的空气温度比全光照低,特别是在14:00 时,全光照下空气温度达42.12℃,10%光照下也有39.2℃,云锦杜鹃处于高温的环境。经T 检验,两个处理的光合有效辐射有显著差异,其它环境因子无显著差异。
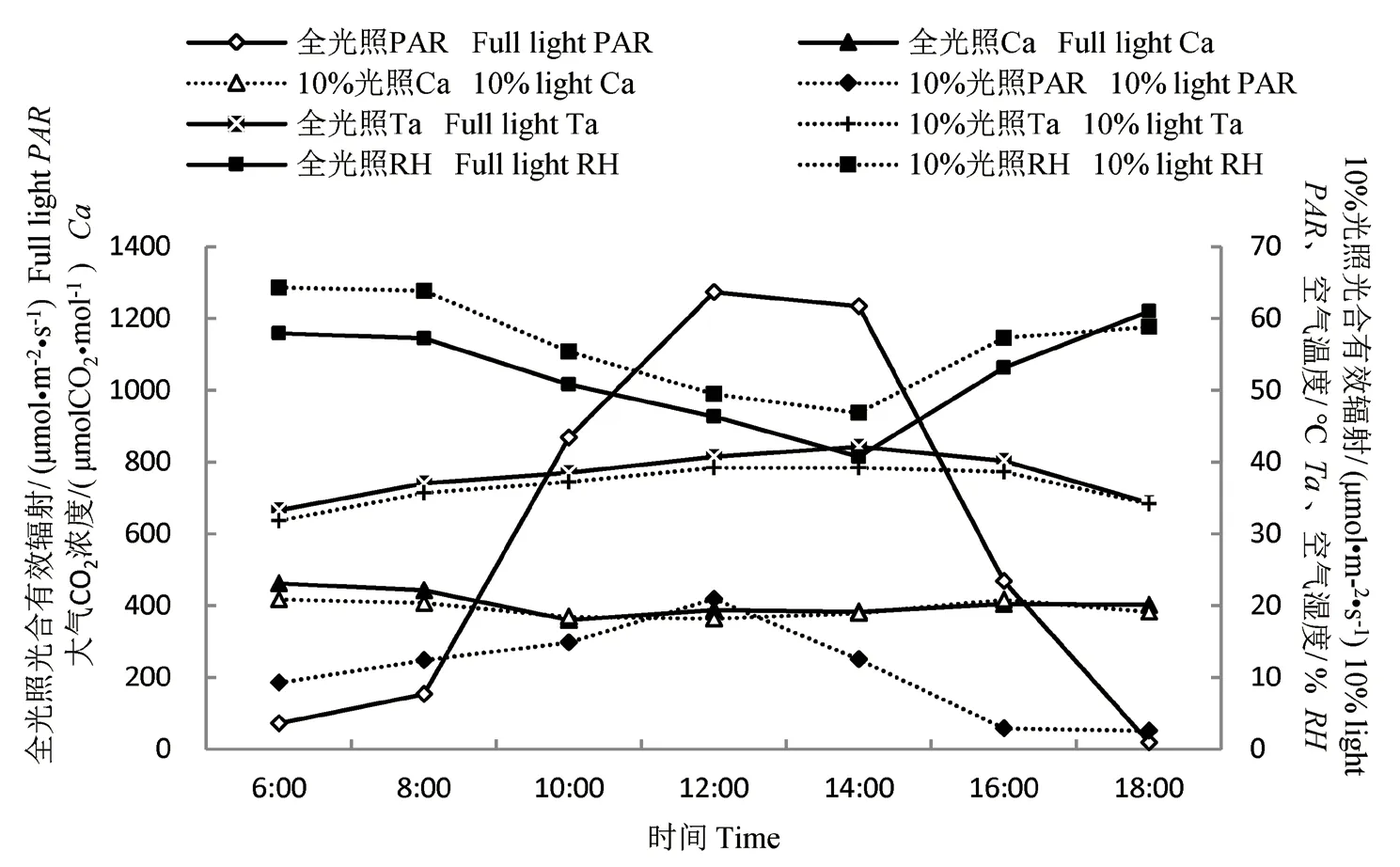
图1 两种光照条件下环境因子的日变化Fig.1 Diurnal variation of environmental factors under two kinds of lights
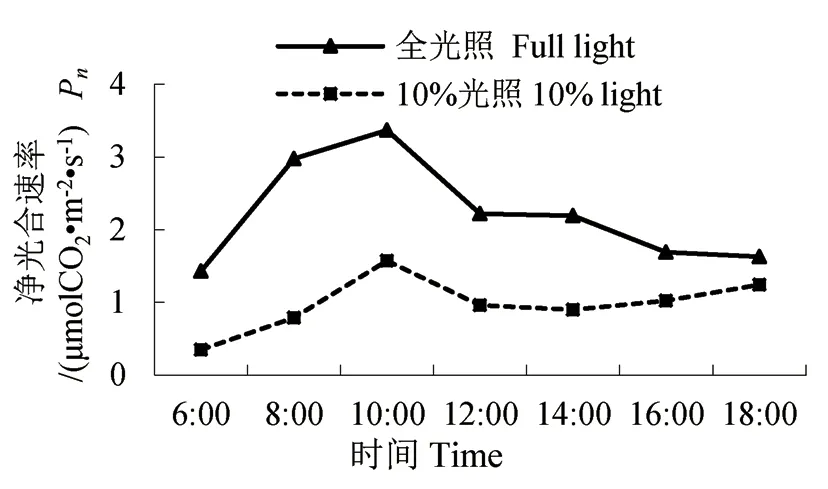
图2 净光合速率的日变化Fig.2 Diurnal variation of PAR
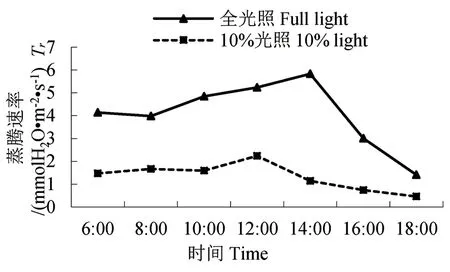
图3 蒸腾速率的日变化Fig.3 Diurnal variation of Tr

图4 气孔导度的日变化Fig. 4 Diurnal variation of Gs

图5 胞间CO2 浓度的日变化Fig. 5 Diurnal variation of Ci
2.2 净光合速率、蒸腾速率的日变化
两种光照下的云锦杜鹃净光合速率、蒸腾速率日变化呈现“单峰”型(图2-3)。它们在10:00时净光合速率达到最高点,10%光照时的净光合速率为1.57 μmolCO2·m-2·s-1,全光照时的净光合速率为3.37 μmolCO2·m-2·s-1,是10%光照时的2.15 倍。10%光照下的云锦杜鹃在12:00 时蒸腾速率达到最高点,为2.24 mmolH2O·m-2·s-1;全光照下的云锦杜鹃在14:00 时蒸腾速率达到最高点,为5.83 mmolH2O·m-2·s-1,是10%光照时的2.60倍。经T 检验,两种处理下净光合速率、蒸腾速率差异极显著。
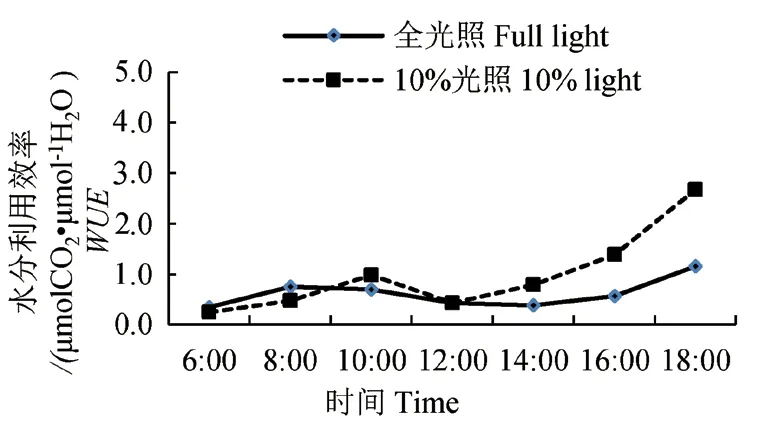
图6 水分利用效率的日变化Fig. 6 Diurnal variation of WUE
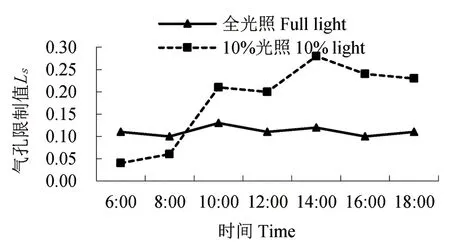
图7 气孔限制值的日变化Fig. 7 Diurnal variation of Ls
2.3 气孔导度、胞间CO2 浓度的日变化
从图4、5 可以看出,云锦杜鹃在全光照下气孔导度日变化呈现“单峰”型,10:00 时达到高峰,为0.209 μmolH2O·m-2·s-1。10%光照下气孔导度日变化呈现“双峰”型,8:00 和12:00 时达到高峰,但都比全光照下的气孔导度小。两种光照下的胞间CO2浓度日变化呈现“双峰”型,6:00和16:00 时达到高峰。这与大气CO2浓度日变化相一致。胞间CO2浓度的日变化基本与净光合速率相反,因为当净光合速率较大时,固定的CO2较多,引起了胞间CO2浓度降低。经T 检验,两种处理下气孔导度差异极显著,胞间CO2浓度不存在显著差异。
2.4 水分利用效率、气孔限制值的日变化
两种光照下的云锦杜鹃水分利用效率日变化呈现“双峰”型(图6),全光照条件下在8:00和18:00 时达到高峰,10%光照条件下在10:00和18:00 时达到高峰,其中18:00 都为最大值,全光照和10%光照条件下分别为1.16 和2.67 μmol-CO2•μmol-1H2O。
与胞间CO2浓度的日变化曲线相反,两种光照下的云锦杜鹃气孔限制值日变化呈现“双峰”型(图7),10:00 和14:00 时达到高峰。10%光照下的气孔限制值变化呈逐渐上升趋势。经T 检验,两种处理下水分利用效率和气孔限制值不存在显著差异。
3 结论与讨论
云锦杜鹃是喜阳植物,需阳光却不耐高温,因此常在平均温度比较低的湿润高山环境下生长[16]。通过光合作用日变化分析,研究结果与之相符合,全光照下云锦杜鹃的净光合速率更高。虽然种植在低海拔环境,但其喜光的特性并不因为温度高而改变。但夏季,光合有效辐射高,会受到光抑制,城市气温又偏高,通过遮荫可以降低温度,提高土壤湿度,减少光合有效辐射,保证云锦杜鹃安全渡夏。午后,10%光照条件下云锦杜鹃水分利用效率高于全光照条件的60.18%,全天蒸腾速率低于全光照条件下的蒸腾速率。这与分布在海拔较高的猴头杜鹃引种到低海拔地区时的表现相似[17]。
两种光照下的云锦杜鹃净光合速率和蒸腾速率日变化都显现“单峰”型。亚热带地区森林植物在高温强光下常有“午休”现象,且日变化为“双峰”型[18],本研究与之不同,净光合速率和蒸腾速率“午休”现象和次高峰不明显。金则新等人[19]曾对自然分布在高山生长的云锦杜鹃进行日变化分析,结果阳叶呈现“双峰”型,其阴叶却呈现“单峰”型,说明叶片光合“午休”由叶片所处的环境因素决定,并非其固有的生理现象。本研究云锦杜鹃引种到低海拔环境后,到夏季需要适应39℃以上的高温,其生长受到一定的影响,改变了原有的光合特性。同时温度可以通过许多酶促反应制约植物光合生产力,高温引起的CO2溶解度、Rubisco 对CO2的亲和力以及光合机构关键成分的热稳定性的降低[20],都可能引起光合速率的降低。
全光照下净光合速率在10:00 时达到最大值后就下降,而胞间CO2浓度却转为上升,表明全光照下净光合速率午后降低不是由于气孔关闭引起胞间CO2亏缺(气孔限制)造成的,而是非气孔限制引起的。这在多种植物的研究中都有相似的结果[21-22]。10%光照下净光合速率在10:00 时达到最大值后下降一段时间后还有一定的上升,胞间CO2浓度的变化也是降低后增加,表明10%光照下净光合速率午后降低是气孔限制的,说明夏季遮荫措施还是有必要的。
