高温胁迫对黄瓜幼苗生理生化及光合作用的影响
2021-04-12崔庆梅吴利荣冯晓东
崔庆梅,吴利荣,陈 斐,冯晓东,2*
(1.延安大学 生命科学学院;2.陕西省红枣重点实验室(延安大学),陕西 延安 716000)
高温是植物种植中的逆境之一,高温使植物失水枯败,光合作用下降,树干容易出现干燥、枯萎、断裂的情况;叶片颜色变褐、变黄、变深,呈现坏死斑纹的现象;新鲜的果实出现日灼的情况,严重影响植物的品质和产量[1]。随着大气中的CO2含量逐年增加,全球气候不断变暖,温室效应带来的恶劣影响进一步加剧,夏季炎热期可能不断增加,日平均温度超过30℃,气温的升高对多种植物的生长和发育都产生了不良的影响,并且给农业造成严重的损失[2,3]。
黄瓜(CucumissativusL.)现栽培于温带以及热带的广大地区,为中国各地方夏天常用菜蔬之一,也是设施蔬菜的主栽品种,黄瓜是一种喜温性蔬菜,最适宜的温度为25℃-30℃,超过35℃就会导致伤害[4],给黄瓜生长及产量造成了严重影响[5,6]。在设施黄瓜的栽培中,从春季到秋季,这一时期都会遇到高温条件,棚室内最高温度可达50℃以上,7、8月间昼平均温度常在35℃左右,影响黄瓜的正常生长发育和光合作用,孟令波等[7]的研究表明高温导致黄瓜过早衰老,产量和品质下降,表现为瓜柄变长,畸形瓜率增加,果皮变硬,VC含量下降;李建建[1]的研究表明,高温处理下植株有一定程度的徒长现象,而茎粗、冠鲜重与冠干重、根鲜重与根干重增加均受到抑制,42℃/27℃(d/n)处理下各项生长指标均受到明显抑制;王立涵等[8]的研究表明,外源喷施精胺、亚精胺、硝普钠和硫氢化钠,皆可减轻高温胁迫对黄瓜幼苗光合器官的伤害,维持抗氧化酶的功能,进而提高黄瓜植株的耐热能力;孙胜楠等[9]的研究表明,适宜光强下短时间高温处理黄瓜幼苗不会产生明显光抑制,高温胁迫会对其PSⅡ反应中心造成严重损伤,光合酶受高温胁迫诱导,但其诱导效应与温度升高幅度和高温持续时间有关。本试验采用不同高温胁迫时间处理,通过测定生理生化和光合作用,研究高温胁迫时间长短对延安设施黄瓜主栽品种生理生化及光合作用的影响程度,旨在探究本地主栽设施黄瓜品种的耐热性,从而为设施黄瓜夏季种植的科学管理提供栽培依据。
1 材料和方法
1.1 材料
试验材料为中研二十一优F1黄瓜。
1.2 研究方法
1.2.1 材料培养
选择颗粒饱满,大小均匀、厚度一致的黄瓜种子,用75%酒精消毒10 min,蒸馏水反复冲洗若干次,用于去掉种子表面的防腐剂,置于2 L的盆中,取一定量的水浸没种子,水的体积以浸过种子为宜。在30℃、生化培养箱中培养,每12 h换1次水,浸泡种子至种子发芽,将催芽的黄瓜种子播种在盛有陶粒的小花盆中,出苗后置于智能光照培养箱内进行培养,设定温度为昼温28℃/夜温18℃,光照12 h,光强为1200 μmol·m-2·s-1,相对d/n湿度控制在74%左右。待幼苗长出第3片真叶时进行高温40℃/25℃(d/n)胁迫处理。
1.2.2 测量方法
净光合速率、胞间CO2浓度(ci)、蒸腾速率的测定使用便携式光合作用-荧光测量系统GFS-3000仪器测定。测定选在晴朗天10∶00-12∶00光照充分且相对稳定的时间,测定部位为同一高度、同一方向营养枝条的叶片中部,重复3次。
叶绿素含量的测定参照文献方法[10];丙二醛含量的测定参照文献方法[11];可溶性糖的测定参照文献方法[12]。
1.3 数据处理与统计分析
试验结果以测定的平均值表示。用SPSS 21.0软件对数据进行统计分析,采用LSD检验和Duncan法进行数据差异显著性多重比较。
2 结果与分析
2.1 对黄瓜幼苗净光合速率的影响
由图1可知,随着高温胁迫时间的增加,黄瓜幼苗净光合速率呈现下降的趋势,与对照相比较,高温胁迫第1 d后光合速率下降较明显,存在显著差异;与高温胁迫1 d后相比较,高温胁迫的第2 d到第5 d,光合速率持续下降,但差异不显著。表明高温胁迫在初期对黄瓜幼苗的伤害较大,随后植物对高温胁迫具有一定的抵抗能力,虽然随着胁迫时间的增加光合速率呈下降的趋势,但下降程度不大。

图1 高温胁迫时间对黄瓜幼苗净光合速率的影响
2.2 对黄瓜幼苗胞间CO2浓度和叶片蒸腾速率的影响
由图2可知,随着高温胁迫时间的增加,胞间CO2浓度在前3 d持续上升,与初始对照相比较差异显著,但与第3 d相比较,高温胁迫的第4 d和第5 d略有下降,差异不显著。随着光合能力的下降,胞间CO2浓度在胁迫的初期上升较为明显,后期下降趋势降低,表明黄瓜幼苗在高温胁迫一段时间后,光合能力可以维持在一定水平。

图2 高温胁迫时间对黄瓜幼苗胞间CO2浓度的影响
由图3可知,随着高温胁迫时间的增加,黄瓜幼苗的蒸腾速率呈现先增加后降低的趋势,但蒸腾速率均高于对照,与对照相比较,蒸腾速率在胁迫后第2 d达到最大,差异极显著,随后开始下降,到胁迫第5 d,与对照相比较,差异不显著。初期蒸腾作用的增加与高温胁迫密切相关,导致幼苗失水增加,但后期蒸腾作用逐渐降低,与对照接近,幼苗具有一定的调节水分代谢的能力。

图3 高温胁迫时间对黄瓜幼苗叶片蒸腾速率的影响
2.3 对黄瓜幼苗叶绿素含量和丙二醛含量的影响
由图4可知,随着高温胁迫时间的增加,黄瓜幼苗叶片中叶绿素含量呈下降的趋势,高温胁迫的前2 d,差异不显著,从第3 d开始,下降幅度较大,与对照相比较,存在显著差异。从叶绿素含量的变化来看,与对照相比较胁迫初期下降幅度较缓,胁迫后期下降幅度较大,对光合作用的影响大。由图5可知,随着高温胁迫时间的增加,黄瓜幼苗叶片中丙二醛含量呈现先升高后下降的趋势,与对照相比较存在显著差异,随着处理时间的增加,黄瓜叶片丙二醛含量在高温胁迫的后期含量增加趋势减缓。

图4 高温胁迫时间对黄瓜幼苗叶绿素含量的影响
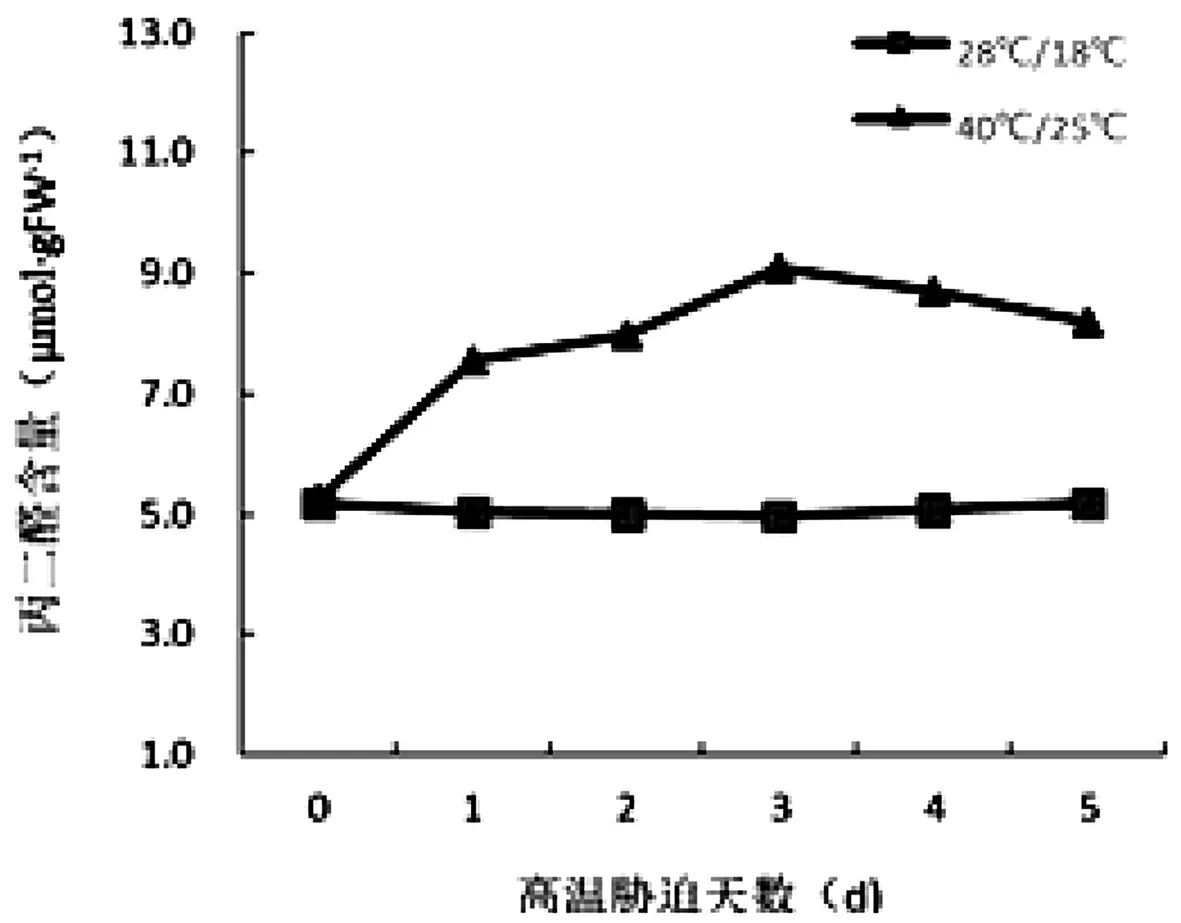
图5 高温胁迫时间对黄瓜幼苗丙二醛含量的影响
2.4 高温胁迫时间对黄瓜幼苗可溶性糖含量的影响
由图6可知,对照组的黄瓜幼苗叶片中的可溶性糖含量随处理时间的增加而逐渐增加,但增加不明显,试验组随着高温胁迫时间的增加,黄瓜幼苗叶片中的可溶性糖含量呈逐渐下降的趋势。胁迫后期试验组和对照组叶片中可溶性糖的含量差异显著,表明高温胁迫对糖的积累影响较大。
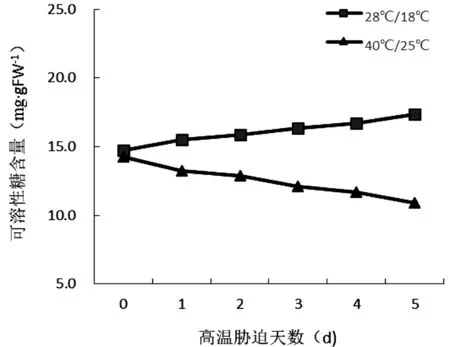
图6 高温胁迫时间对黄瓜幼苗可溶性糖含量的影响
3 讨论与结论
高温胁迫环境可以造成植株生长发育受阻和抵抗逆境能力降低,阻碍植物的养分积累和细胞的代谢,从而导致蔬菜的产量和品质降低[13,14]。从试验结果来看,随着黄瓜幼苗高温胁迫处理时间的增加,与对照组相比较,幼苗的叶绿素含量、净光合速率、胞间CO2浓度、蒸腾速率、丙二醛含量和可溶性糖含量均存在显著差异。
在一般情况下,随着温度的增长,高温逆境损伤植物的进程大抵依次为阻挠植物的生长发育—植物“饥饿”—有毒有害物质进行累积—生物化学伤害导致蛋白质发生降解—有机离子从细胞膜溢出—丢失半透性膜的能力—化学解体[15]。根据马德华等[16]的研究,高温使光合作用下降的真正原因在于叶片同化CO2的阻力升高,并且羧化位点会产生羧化阻力,植物受到高温胁迫时,也会损害细胞膜的通透性,致使细胞液向外渗漏,H+的透性不断增加,叶绿体内外正常稳定的H+浓度被破环,光系统反应受到遏制,致使合成的ATP含量下降,进而导致1,5-二磷酸核酮糖的合成减慢。由图1、图4可知,随着高温胁迫时间的增加,黄瓜幼苗的净光合速率和叶绿素含量呈现不断地下降的趋势,叶绿素含量下降,表明幼苗叶片对于光的吸收与转化能力不断下降,叶绿素含量降低的可能缘由在于合成叶绿素的中间产物被破坏,或者是因为高温导致植物的氧化作用加剧。由图6也可以看出,可溶性糖的含量也在持续下降,表明光合产物也在减少,原因可能是高温胁迫影响了与可溶性糖代谢有关的酶系统,同时也影响在植物体内的输送以及储藏[17];由图2可知,黄瓜幼苗胞间CO2浓度呈现出先升高后降低的趋势,一般胞间CO2浓度与光合作用为正相关,与气孔导度为负相关,在胞间CO2浓度升高的情况下,幼苗的净光合速率下降,表明高温胁迫造成了黄瓜幼苗光合系统的损伤,与CO2的供应关系不大。
由图3可知,随着高温胁迫时间的增加,黄瓜幼苗蒸腾速率呈现先增加后减少的趋势,高温胁迫初期幼苗叶片保水能力较弱,非气孔蒸腾的比例较大,导致蒸腾速率快速上升,胁迫后期,随着幼苗叶片含水量的降低,蒸腾作用也随之降低。丙二醛是膜脂过氧化反应的生成物,植物细胞膜被破坏后,过氧化物丙二醛含量增多,它的堆积会对植物的细胞膜造成进一步的损伤[18]。由图5可知,随着高温胁迫时间处理的增加,黄瓜幼苗丙二醛含量先增加后降低,表明高温胁迫初期对幼苗叶片细胞膜的损伤较大,后期缓慢降低,可能与幼苗具有一定适应高温胁迫的能力。
综上所述,随着高温胁迫处理时间的增加,黄瓜幼苗叶片中的叶绿素含量、净光合速率、胞间CO2浓度、蒸腾速率、丙二醛含量和可溶性糖含量均发生了显著变化。其中净光合速率、叶绿素含量和可溶性糖含量呈现逐渐下降的趋势,相反,胞间CO2浓度呈现先升高后缓慢下降的趋势,表明高温引起的光合作用的下降非气孔因素引起,同时,随着高温胁迫时间的增加,幼苗叶片中丙二醛的含量呈现先增加后缓慢下降的趋势,表明黄瓜幼苗在高温胁迫下,细胞膜的结构受到了损伤,高温胁迫降低光合的主要原因可能是光合系统受损造成的;蒸腾作用表现出先增加后降低的趋势,主要原因是高温胁迫初期幼苗叶片保水能力较弱,高温导致蒸腾作用增强,幼苗失水,也是导致光合能力降低因素之一;高温胁迫后期,丙二醛含量及蒸腾作用降低,同时净光合速率和叶绿素含量下降减缓,表明黄瓜幼苗对高温胁迫具有一定的抵抗能力。
