不同生长年限独一味的形态组织学研究
2021-04-10钟世红
鲍 新,刘 琴,古 锐,钟世红,2
(1.成都医学院药学院,成都 610500;2.西南民族大学药学院,成都 610200;3.成都中医药大学民族医药学院,成都 610075)
独一味来源于唇形科独一味属植物独一味[Lamiophlomis rotata(Benth.)Kudo],是历史悠久的藏族常用药材,具有活血止血、祛风止痛的功效,广泛用于治跌打损伤、外伤出血、风湿痹痛等症[1,2]。近年来,独一味止痛效果良好且无成瘾性的优势,引起了业界的广泛关注,以独一味为主要原料的成方制剂及下游产品的开发日益增多。逐年增长的药用需求,导致其连年过度采挖,加之生态环境不断退化,独一味药材资源日益减少,于2000 年被列为一级濒危藏药品种[3,4]。由于独一味尚未实现规模化种植,商品全部来源于野生资源,因此研究独一味种群特征,进而开展规模化种植已迫在眉睫。
种群中各年龄期个体数在种群中所占的比例是种群的年龄结构[5],植物种群年龄结构的确定,是分析种群结构特征、分布格局、演替规律的重要前提,可以解释个体、种群与生境的相互关系。植株龄级的判断是研究种群年龄结构的基础[6,7],且其生长年限的确定也与药材的品质息息相关[8]。与木本植物可通过其年轮的环数确定年龄不同[9-11],草本植物在形态结构上未见如此普及的鉴别特征,仅局限于禾本科、莎草科等植物依据分蘖类型判断植株年龄[12,13]。多年生草本植物龄级划分的困难,导致草本植物种群特征研究相对不足。近年来,一些多年生草本植物已陆续报道具有生长轮构造[14,15]。包括药用植物中的类似生长轮构造,如人参[16]、芍药[17]和川续断[18]可在主根观察生长轮,黄芩[19]根木质部导管的排列方式可作为其生长年限辨别依据。然而,并非所有的多年生草本植物都具有生长轮构造,有的植物生长轮构造仅在部分产地植株中出现[20],多年生草本植物的年龄确定相对较为困难。
本研究采集1~3 年生栽培独一味植株,观察并比较了不同生长年限独一味的外观性状和显微构造,初步获得了不同生长年限独一味的形态组织学鉴别特征。为验证结果,亦采集野外不同大小等级的独一味进行观测,来源于同一样地的小、中、大等级野生独一味植株表现出类似1~3 年生栽培独一味的形态组织学特征差异。研究结果可为独一味种群年龄结构研究、种群动态预测提供参考,进而为合理利用、保护野生独一味资源提供科学依据。
1 材料与方法
1.1 材料
于2016 年9 月至2018 年9 月,在四川省阿坝州若尔盖县热尔大草地收集野生独一味种子,播撒于四川省阿坝州茂县三龙乡进行栽培种植,于2020 年5 月采集1~3 年生独一味植株各15 株。2020 年9 月在四川省甘孜州稻城县采集未知年限的小、中、大3组独一味样品各15 株,经西南民族大学药学院钟世红教授鉴定为唇形科植物独一味。
1.2 仪器与试剂
10000XL 根系分析扫描仪(日本EPSON 公司,WinRHIZO 分析系统);BX41 和BX63 正置荧光生物显微镜(日本OLYMPUS 公司);SMZ168 体式显微镜(中国Motic 公司);TK-218 型自动轮转切片机(德国Leica 公司);B7126-A 型包埋机(德国Leica 公司);番红O-固绿植物组织染色液(上海源叶生物科技有限公司)。
1.3 方法
1.3.1 测量性状特征 独一味新鲜植株用去离子水洗净后滤纸吸干水分,电子分析天平分别称量独一味地上部分和地下部分鲜重;用坐标纸测定叶片长度和宽度;采用根系扫描仪扫描独一味根系,保存图片后运用WinRHIZO 2009 根系系统分析软件分析总根长、总根表面积、总根体积、主根长、主根表面积、主根体积、主根直径及根尖数等指标。
1.3.2 石蜡切片的制备 取新鲜1~3 年生栽培独一味与未知生长年限小、中、大等级野生独一味各15株,放入FAA 固定液中固定后,梯度乙醇脱水、梯度二甲苯透明、浸蜡、包埋、切片(厚度为8 μm),番红O-固绿染色,封片,分别制成叶横切片与根横切片,置于生物显微镜下观察与拍照。
1.3.3 徒手切片的制备 撕取独一味下表皮,稀甘油装片,取任意清晰可见的视野置于显微镜下观察与拍照,每张装片观察3 个视野,观察气孔、腺鳞的形态与数目,计算腺鳞密度[单位面积内腺鳞的个数(个/mm2)]和气孔指数[气孔指数=100×气孔个数/(气孔个数+表皮细胞个数)]。
1.4 数据处理
采用SPSS 19.0 软件进行数据处理与分析,使用ANOVA 和Duncan’s 法对试验数据进行差异性分析。所有指标的数据均以15 个生物重复的平均值±标准误差(±s)表示。
2 结果与分析
2.1 不同生长年限栽培与野生独一味外观性状
独一味为多年生草本;贴地生长;叶对生,4~6枚,多为菱状圆形,基部下延至叶柄,边缘具圆齿,叶表面皱缩,具短柔毛。一年生独一味主根细长,无分枝,须根较少;二年生独一味主根分枝0~2 个,须根较多;三年生独一味主根粗大,偶见剥落的栓皮,较粗的根偶见枯心,根中部及下部出现分枝2~5 个,须根茂密(图1)。

图1 栽培独一味
由表1 可以看出,栽培独一味鲜重、根重、株高、叶长、叶宽、叶面积、根表面积、根体积、主根表面积、主根体积及主根直径等随着年限的增加均呈上升趋势且有显著差异(P<0.05)。说明1~3 年均为独一味的快速生长期。不同生长年限栽培独一味的生物量积累趋势存在差异,一年生根冠比为0.50,二年生根冠比为0.52,三年生为0.40,结合地上、地下生物量随着生长年限呈上升趋势,说明独一味幼苗最初的2 年地上、地下部分增长接近,从第3 年开始,地上部分增长更为迅速。
表1 不同生长年限栽培独一味生物量比较(±s,n=15)
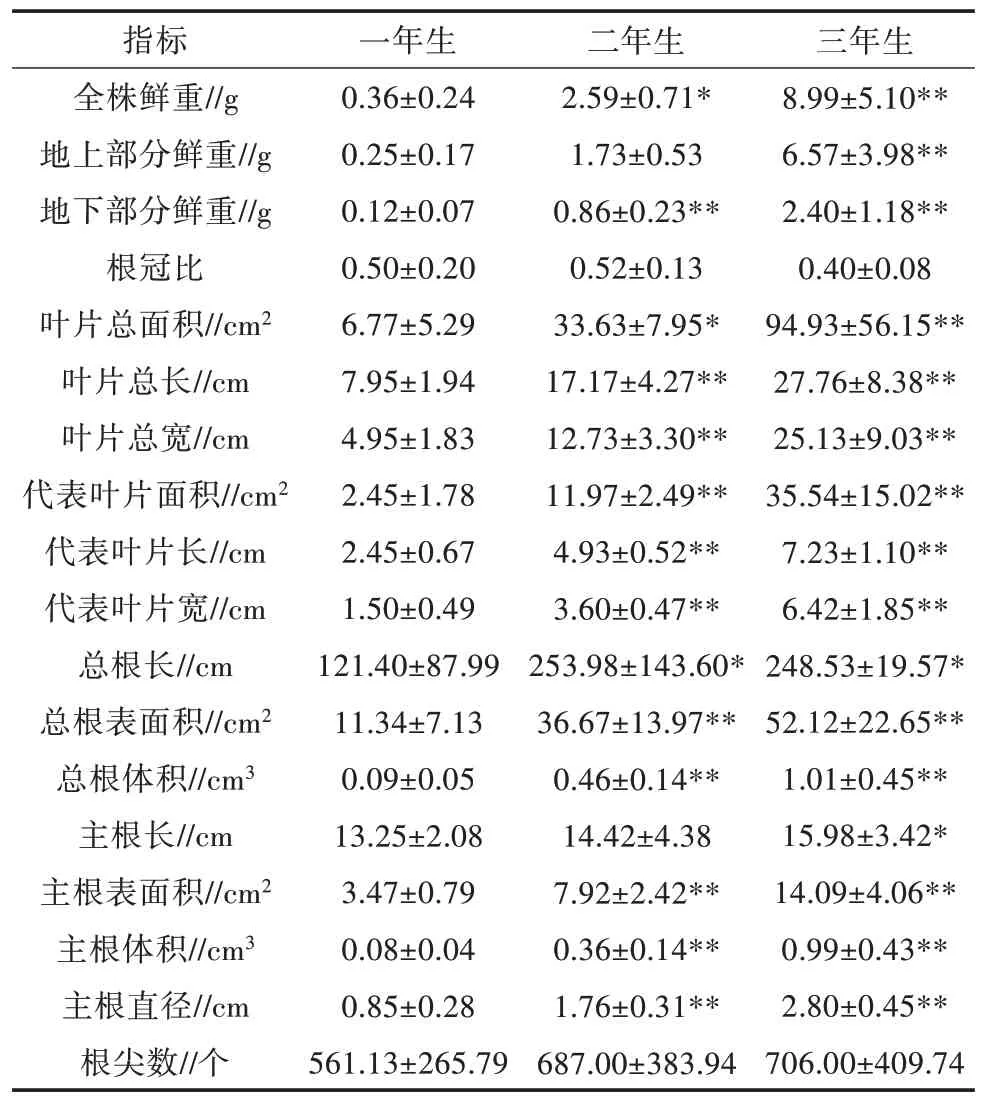
表1 不同生长年限栽培独一味生物量比较(±s,n=15)
注:“*”为差异显著(P<0.05),“**”为差异极显著(P<0.01);表2同
指标 一年生 三年生二年生全株鲜重//g地上部分鲜重//g地下部分鲜重//g根冠比叶片总面积//cm2叶片总长//cm叶片总宽//cm代表叶片面积//cm2代表叶片长//cm代表叶片宽//cm总根长//cm总根表面积//cm2总根体积//cm3主根长//cm主根表面积//cm2主根体积//cm3主根直径//cm根尖数//个0.36±0.24 0.25±0.17 0.12±0.07 0.50±0.20 6.77±5.29 7.95±1.94 4.95±1.83 2.45±1.78 2.45±0.67 1.50±0.49 121.40±87.99 11.34±7.13 0.09±0.05 13.25±2.08 3.47±0.79 0.08±0.04 0.85±0.28 561.13±265.79 8.99±5.10**6.57±3.98**2.40±1.18**0.40±0.08 94.93±56.15**27.76±8.38**25.13±9.03**35.54±15.02**7.23±1.10**6.42±1.85**248.53±19.57*52.12±22.65**1.01±0.45**15.98±3.42*14.09±4.06**0.99±0.43**2.80±0.45**706.00±409.74 2.59±0.71*1.73±0.53 0.86±0.23**0.52±0.13 33.63±7.95*17.17±4.27**12.73±3.30**11.97±2.49**4.93±0.52**3.60±0.47**253.98±143.60*36.67±13.97**0.46±0.14**14.42±4.38 7.92±2.42**0.36±0.14**1.76±0.31**687.00±383.94
未知年限的野生独一味外观性状与人工栽培独一味性状呈相似规律,无论是地上部分还是地下部分生物量均出现增大趋势。随着植株的增大,根愈发粗壮且栓皮脱落,分枝逐渐增多,根部枯朽加重(图2)。

图2 野生独一味
2.2 不同生长年限栽培与野生独一味显微特征
2.2.1 叶片下表皮 栽培独一味的下表皮细胞垂周壁平直或微弯曲,气孔多为不定式或不等式,可见腺鳞,腺头多为4 细胞(图3)。由表2 可以看出,随着年限的增加,腺鳞密度减少且有极显著差异(P<0.01),气孔指数无明显变化。
野生独一味的下表皮气孔、腺鳞和表皮细胞形态相似,不同大小等级的植株特征无明显变化(图4)。
2.2.2 叶横切面 栽培独一味叶上、下表皮细胞各1 列,外被角质层,偶见非腺毛和腺鳞。栅栏组织排列紧密,多为2~3 列;海绵组织排列疏松,不过中脉。中脉维管束为外韧型,“U”形排列。1~3 年生独一味下表皮内侧均为2~3 列厚角组织;一年生独一味上表皮内侧未发现厚角组织;二年生独一味上表皮内侧偶见厚角组织1~2 列;三年生独一味上表皮内侧厚角组织2~3 列。一年生独一味叶中脉维管束1个,木质部导管十几个聚集呈团状排列;二年生独一味叶中脉维管束1~2 个,木质部导管数十个聚集呈环状排列;三年生独一味叶中脉维管束1~3 个,木质部导管数十个聚集呈环状排列(图5)。
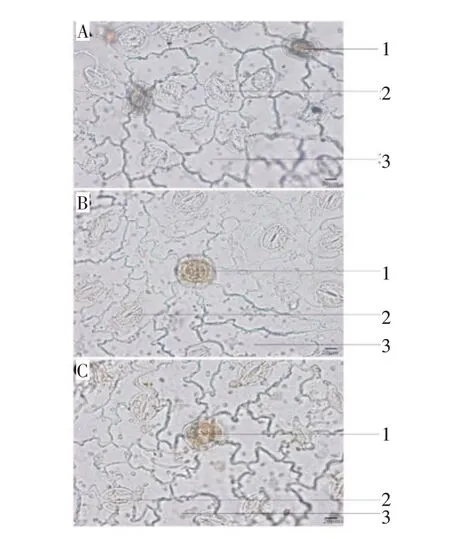
图3 栽培独一味叶下表皮(×400)

图4 野生独一味叶下表皮(×400)
结合表2 可知,随着年限的增加,中脉逐渐膨大,叶片厚度逐渐增加且有极显著差异(P<0.01),上、下表皮变化不显著。
野生独一味叶横切面内部结构与栽培独一味相似。小株独一味中脉维管束1~2 个,上、下表皮内侧厚角组织2~3 列;中级大小独一味中脉维管束1~3个,上、下表皮厚角组织3~5 列;大株野生独一味中脉维管束1~4 个,上、下表皮内侧厚角组织4~5 列(图6)。
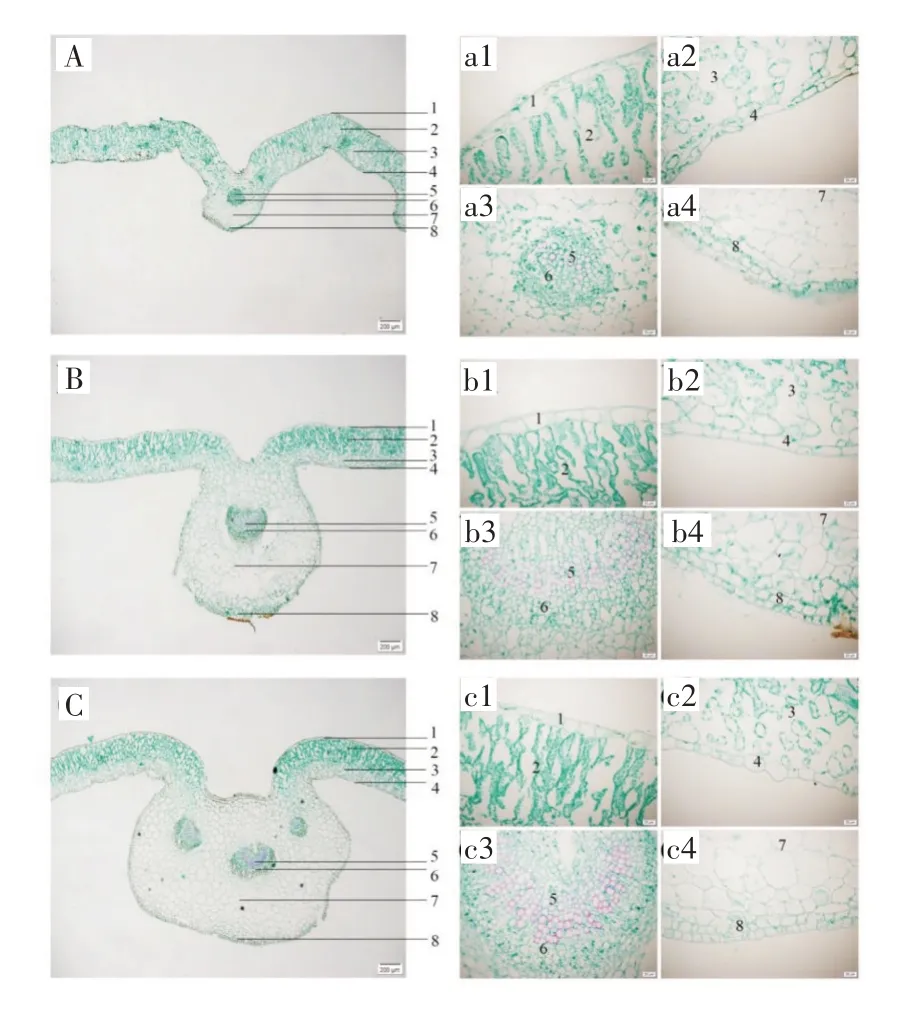
图5 栽培独一味叶横切面
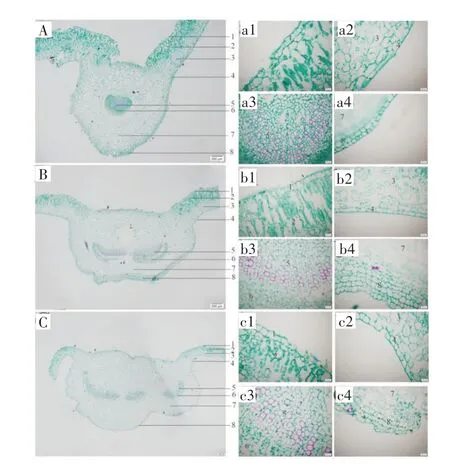
图6 野生独一味叶横切面

表2 不同生长年限栽培独一味叶片指标比较(x±s,n=15)
2.3 根横切面
栽培独一味根木栓层由1~2 列细胞组成,呈深棕色;韧皮部细胞排列较紧密,形成层明显,呈环状,木质部宽广,不同生长年限栽培独一味根导管排列方式不同。1~2 年生根导管单个散在或数个成束,呈扁平层状排列,一年生根导管排列紧密,二年生根导管排列较稀疏;三年生根导管数十个成束切向连续排列成与环状结构(图7)。

图7 栽培独一味根横切面
野生独一味根部显微结构与栽培独一味相似,随着植株的增大,枯朽逐渐加重。小株独一味根中部细胞排列紧密,导管数十个成束切向连续排列成环状结构;中级大小独一味根中央部位具多轮木栓细胞环,其内侧薄壁细胞消失,有较大裂隙,各木栓细胞环层之间导管亦呈断续环状分布;大株独一味根中部亦出现木栓细胞环,其内侧枯朽严重,呈较大空洞(图8)。

图8 野生独一味根横切面
3 小结与讨论
本研究对1~3 年生栽培独一味从外观性状和组织构造进行了比较研究,不同生长年限的独一味表现出一定的差异。为了进一步验证,亦采集野生独一味进行观察,虽不能确定野生植株具体生长年限,但基于前期多年持续的野外观察数据,叶片大小与龄级大小呈正相关趋势,因此观察来源于同一野生样地小、中、大3 种大小级别的植株,获得野生独一味植株不同生长年限的特征趋势。观察结果表明,同一野生样地小、中、大3 种大小级别的独一味植株,具有类似栽培1~3 年生样品的特征规律。
性状特征比较显示,随着生长年限的增加,栽培独一味出现分枝,二年生独一味分枝0~2 个,三年生独一味分枝2~5 个,且主根粗大,部分较粗的主根出现裂隙、枯心现象,偶见剥落的栓皮,须根较繁茂。而1~2 年生独一味根未见枯心和剥落的栓皮,可据外部性状初步判断三年生独一味。不同大小等级野生独一味外观性状出现类似栽培1~3 年生栽培样品的特征规律,随着植株的增大,主根出现裂痕,且栓皮剥落程度、中部枯朽现象逐渐加重。
显微组织观察显示,三年生栽培独一味叶中脉维管束较发达,可达3 个,呈“U”形排列,而一年生叶的中脉维管束仅1 个;二年生叶的中脉维管束1~2个;三年生独一味叶中脉上下表皮以下均可见厚角组织2~3 列,一年生叶中脉上表皮以下未见厚角组织,二年生叶的中脉上表皮以下偶见厚角组织1~2列。三年生独一味根木质部导管数较多,数十个成束切向连续排列成环状结构,而一年生根和二年生根导管单个散在或数个成束,未形成较连续的环状结构。显微组织方面可以从叶“U”形排列的中脉维管束,中脉上下表皮以内的厚角组织,根环状排列的木质部导管判断三年生独一味。
野生独一味叶中脉维管束和厚角组织的发达程度与植株的大小呈正比,与上述栽培独一味规律相似。不同大小等级野生独一味根内部结构生长规律类似栽培1~3 年生样品的特征规律,小株独一味根中心细胞排列紧密,木质部导管数十个成束切向连续排列成环状结构;中等大小独一味根中心开始出现栓化,中部木栓层含有导管与木质部导管呈平行环状;大株独一味根中心出现较大空洞,中部木栓层与外部木栓层呈同心环状。
独一味为多年生高原草本植物,但目前仅能采集可确定栽培年限的1~3 年生独一味植株进行试验。课题组亦采集了不同产地的野生独一味植株进行形态组织学观察,尤其仔细观察了粗壮的大型植株,表现出主根粗壮、枝根数量多、须根繁茂、主根横切面多有不同程度枯心的性状特征;在根的显微构造上导管聚集明显,切向排列成多轮断续的扁平带状。随着生长年限的增长,独一味是否体现出类似生长轮的构造,课题组将持续开展研究。
试验试图从次生代谢产物积累角度,寻找不同生长年限的独一味植株差异。独一味地上部分主要含有环烯醚萜类、苯乙醇苷类、酚酸类及黄酮类成分,地下部分主要含有环烯醚萜类和苯乙醇苷类成分。测定结果表明,无论是1~3 年生栽培独一味样品,还是不同大小等级野生独一味样品,其次生代谢产物含量均无显著差异,即从次生代谢产物积累的角度暂未发现规律。
