黄土区土坎植被群落草本优势种的种间关联性研究
2021-04-08申旭芳康永祥巫翠华张利利
申旭芳,康永祥,李 华,巫翠华,张利利
(西北农林科技大学 林学院,陕西 杨陵 712100)
种间关联性是植物群落重要的数量和结构特征之一[1],是群落形成和演化的基础。植物群落中的每一个物种都不是单一存在的,物种间存在着复杂的关系,种间联结则是种间关系最直接的一种表现形式[2]。种间关联是指不同物种在空间分布上的相互关联,主要是由于物种生境的差异对物种分布产生影响而引起的[3-5],研究植物的种间关系可以了解不同物种间的相互作用以及不同植物在环境中的适应程度,对认识群落结构组成以及群落的动态变化有重要意义[6]。种间关系不仅可以反映现阶段的群落结构现状,也可以预测群落未来演替进程和演替方向,为群落植被恢复提供理论依据[7-8]。
黄土高原是我国水土流失严重的地区之一,其重力侵蚀又是水土流失的重要策源,尤其是在黄土丘陵沟壑区,土坎、陡坡密集,重力侵蚀严重,目前防控土坎、陡坡滑塌已成为治理水土流失的重要措施。土坎所处地理位置特殊,人为干扰小,植被恢复时间长,其长期处于极度干旱的状态,生长占优势的植物大部分为旱生植物,且草本物种居多,研究这些植物的特性是解决重力侵蚀最为有效的方法[9],同时充分认识土坎群落结构特征对于人工进行黄土区植被恢复和重建起着重大作用。但是目前国内大部分学者对黄土高原植被种间关系的研究多集中于人工植被和沟坡植被种间关系,关锐敏等[10]对黄土高原马栏林区顶级群落进行了种间关系研究,结果表明群落总体上呈现显著正关联,种对间的竞争在环境容许范围内,群落处于稳定状态;许金石等[11]对陕北桥山林区主要木本植物进行了种间关联分析,发现本区物种之间的联结大部分比较松散,并根据种间联结关系划定了生态种组;韩锦涛等[12]对丘陵区人工植物优势种种间关系进行了研究,说明大多数人工植物群落结构简单,且结构不稳定,在植被恢复过程中,应选择不同层次间呈现极显著关联的物种;陈文思等[13]对黄土区沟坡植被优势种的种间关系进行了研究,结果表明茭蒿、铁杆蒿、阿尔泰狗娃花等适应性比较强,在该区分布广泛,且发现大多数植物分布较为独立,群落不稳定,仍处于演替中。关于黄土区天然土坎植被群落优势种种间关系的研究相对比较少。本研究则以永寿县、长武县、西峰区的黄土区土坎植被群落为研究对象,采用方差比率法、χ2检验、联结系数AC、共同出现百分率PC和Spearman秩相关系数分析土坎草本植物优势种种间关系,揭示不同物种间的相互关系和物种共存机制[14],评价了优势种对环境的适应性,为重力侵蚀治理和固沟保塬提供理论支撑和技术依据。
1 研究区概况
黄土高原土层深厚,但是由于自然因素再加上人为破坏,土层表现比较疏松,造成严重水土流失,重力侵蚀严重。本研究选取3个区域作为研究地点,分别为永寿县、长武县和西峰区,永寿县属于黄土高原丘陵沟壑区,长武县和西峰区属于残塬沟壑区,具体研究区概况见表1。
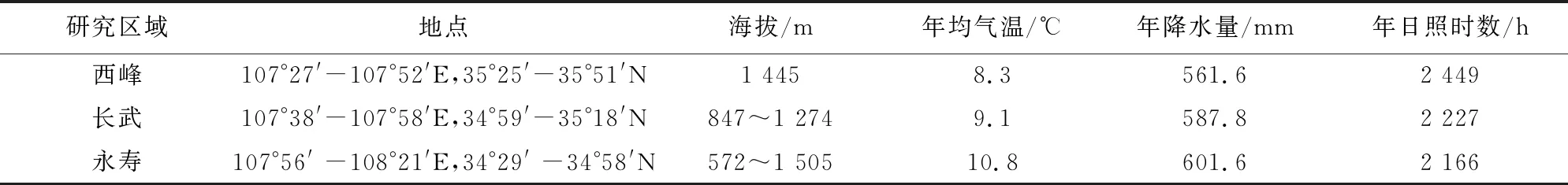
表1 研究区基本概况
2 研究方法
2.1 样地设置与调查方法
2019年7-9月,在西峰区、长武县和永寿县分别选取22块、17块、11块典型样地。样地设置选取土坎自然生长的典型植被群落作为对象,以土坎(崖)为天然样线,根据土坎(崖)地面植被带宽窄,向内扩展2~3 m,样地设置为狭长的长方形,面积为30 m2,在样地内沿着天然样线均匀布设3个2 m×2 m的灌木样方和3个1 m×1 m的草本样方,记录样方内灌木的高度、冠幅、盖度、丛数和草本的高度、多度、盖度、丛数,同时记录所在样地的经纬度、海拔、坡度和土坎高度等信息。对于现场难以鉴定植物种类,采集标本带回实验室进行鉴定。
2.2 数据分析
2.2.1 重要值和优势种的确定 以样地为单位,计算每个物种的重要值。
计算公式如下:
IV=(相对多度+相对频度+相对盖度)/3
根据野外调查结果,以物种重要值作为指标进行排序,选择草本优势种。
2.2.2 总体关联性检验 根据物种在样地中是否存在,形成物种与样地的二元矩阵,采用方差比率法,确定优势种间的总体关联程度,计算公式如下:
(1)
Pi=ni/N
(2)
(3)
(4)
式中,VR代表的是物种总体关联性指数,ni表示物种i出现的样地数,Tj表示在样地j内目标物种数量,t表示样地中物种的平均数,N表示样地总数,S表示总的物种数。当VR>1时,物种总体上正关联;当VR<1时,物种总体上负关联;当VR=1时,表示所有种间无关联。

2.2.3 种对间关联性检验 基于2×2列联表(表2),用Yates的连续校正系数公式[15]计算χ2检验统计量,公式如下:
(5)
ad-bc>0,表示2个物种正关联;ad-bc<0,表示2个物种负关联;ad-bc=0,表示2个物种相互独立。如果χ2>6.635,表示2个物种间联结极显著;如果3.841<χ2<6.635,表示2个物种间联结显著;如果χ2<3.841,表示2个物种间联结不显著。

表2 种间关联性测定2×2列联表
2.2.4 种间关联度的测定 采用联结系数AC和共同出现百分率PC进行计算,公式如下:
如果ad≥bc,则:
AC=(ad-bc)/[(a+b)(b+d)]
(6)
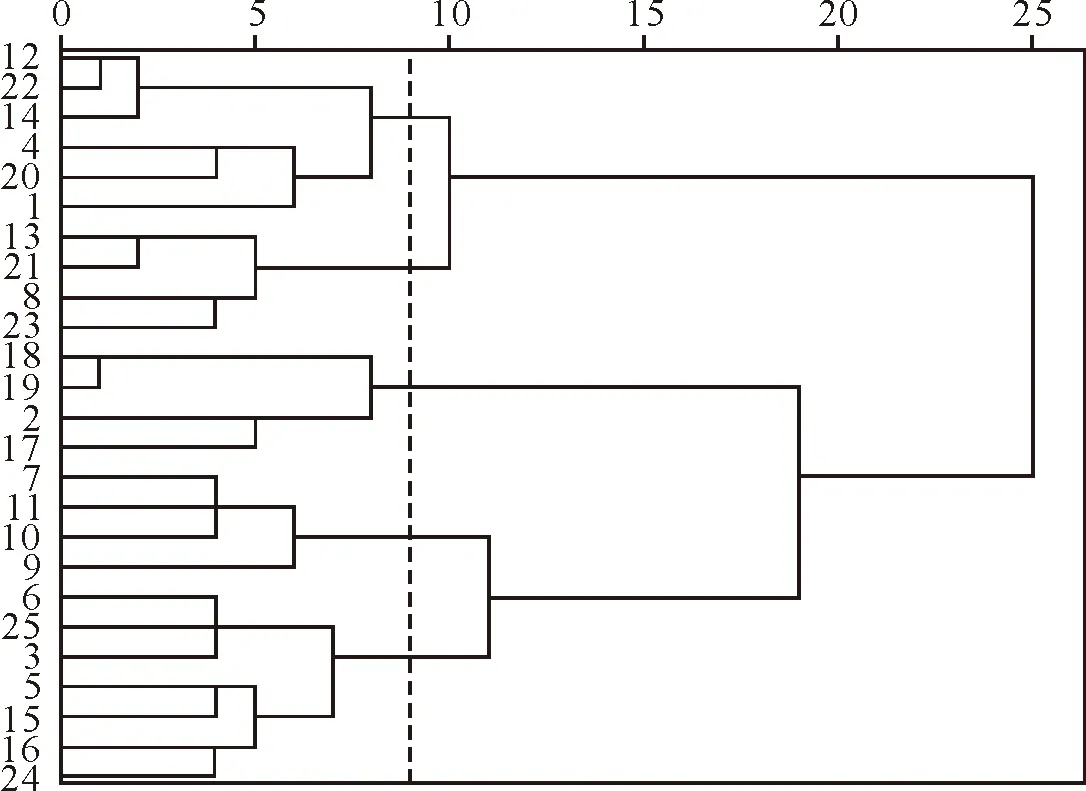
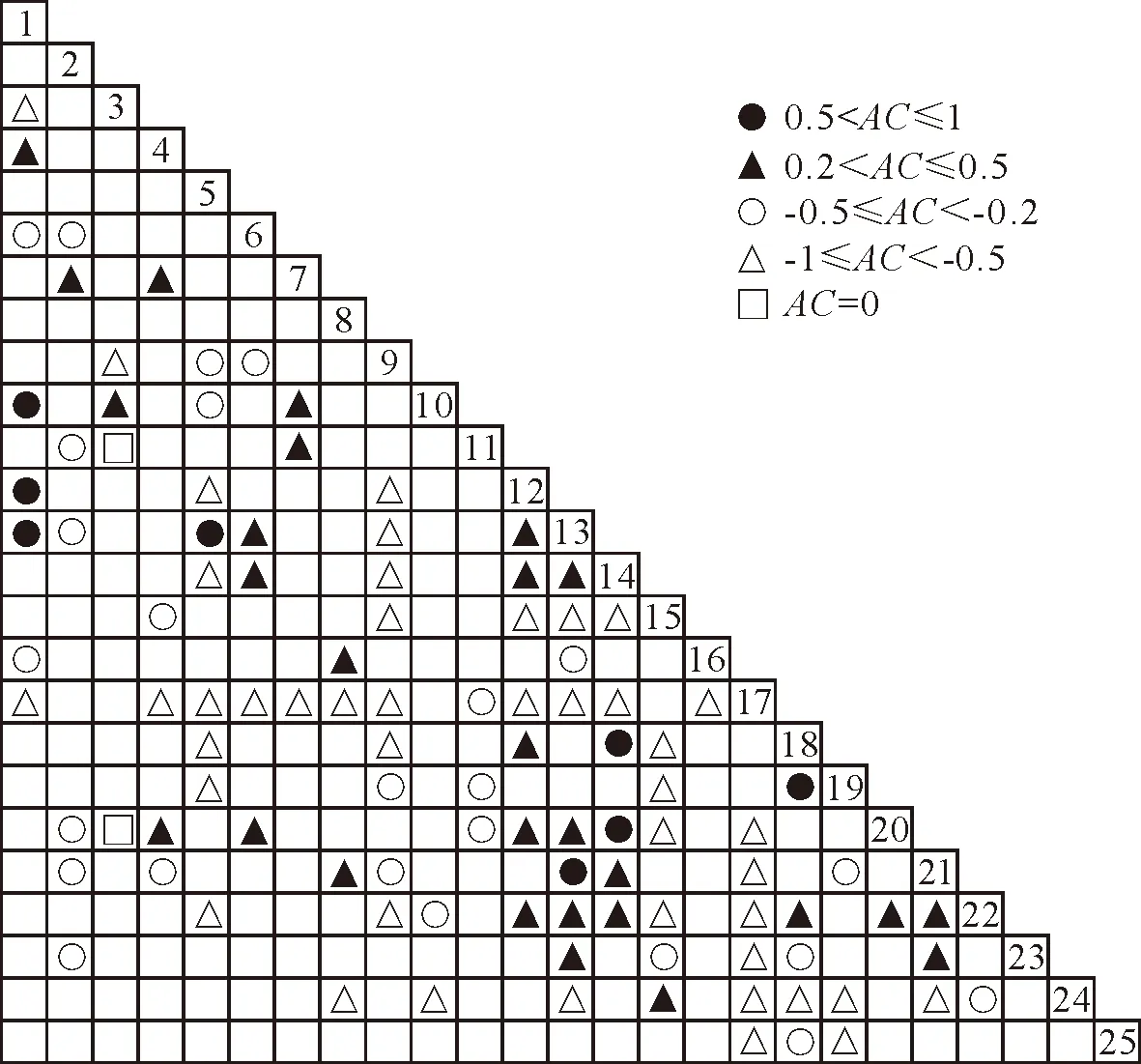
ad AC=(ad-bc)/[(a+b)(a+c)] (7) ad AC=(ad-bc)[(b+d)(d+c)] (8) 式中,AC表示种间联结强度,AC距离1越近,表示2个物种正关联越强,AC距离-1越近,表示2物种负关联越强,当AC=0时,说明2个物种无关联,单独出现[16]。 PC=a/(a+b+c) (9) 式中,字母表示的含义见表2。 2.2.5 Spearman秩相关分析 形成样地×物种数据矩阵,物种数据使用重要值,对优势物种种间关系进行Spearman秩相关分析[4]。 2.2.6 聚类分析 根据Spearman秩相关系数对25个草本优势种进行聚类分析,按照欧式距离进行生态种组划分[2]。 数据处理使用SPSS23.0和Excel2007软件完成。 对黄土区土坎植被的调查结果显示,共出现115种植物,其中有94种草本植物,包含29科76属,主要以菊科(Compositae)、禾本科(Gramineae)和豆科(Leguminosae)为主,其中菊科有14属21种,禾本科有14属15种,豆科有8属14种,分别占草本植物总数的22.34%、15.96%、14.89%;灌木共有14种,分属7科12属,其中豆科植物共6种,占灌木植物总数的46.86%;乔木7种,共6科7属,蔷薇科(Rosaceae)有2种,占乔木总数的28.57%,其余科均有1种。由于立地条件的限制,草本植物数量居多,故本研究筛选出重要值>1%的25种草本优势种进行种间关联性分析(表3)。 表3 草本优势种频度及重要值 3.3.1 χ2检验分析 种间关联性不仅反映了物种间的相互关系,也反映了物种本身的生物学特性以及对环境的适应程度[17-18],物种间呈现正关联,主要是由于物种对生境有着相同的适应性,而负关联是由于物种需要不同的资源环境[19-20]。25个草本优势种共组成300个种对,根据χ2检验结果(图1)可知,相互独立的种对有2对,正关联的种对163对,占总对数的54.33%,负关联的种对135对,占总对数的45%,正负关联比为1.21∶1,其中呈极显著正关联和显著正关联的种对分别为10对和5对,呈极显著负关联的种对和显著负关联的种对分别为2对和3对,显著关联和极显著关联的种对数占总种对数的6.67%,表明虽然草本物种间具有一定的关联性,但是大部分关联性比较弱,表现出较大的随机性。茭蒿和阿尔泰狗娃花、茵陈蒿和阿尔泰狗娃花、野菊与披针叶薹草等种对呈现显著正关联,可能由于这些植物的生态幅宽,适应能力强,均为旱中生或中生植物,多见于土石干燥山坡;茵陈蒿和异叶败酱、硬质早熟禾和披针叶薹草、野菊等种对呈现显著负关联,前者属于旱生植物,多分布于干燥的山坡,后者则属于中生植物,喜相对湿润的环境,这些种对环境的需求不同,缺乏共生的生态基础,故呈现负关联。 图1 草本优势种χ2检验半矩阵 3.3.2 关联度指数分析 对种对间关系进行分析时,χ2检验仅反映了种对间联结性是否显著,但并不代表经χ2检验没有显著性的种对间就不存在联结性,联结系数AC和共同出现百分率PC则弥补了χ2检验的缺陷,能体现出种对间联结性强弱[21]。 由联结系数AC半矩阵图(图2)可以得出,AC=0的种对数有2对,分别为赖草和茜草、赖草和北柴胡,表明这2个种对间完全独立,对生境要求不同。AC值在0.5 图2 草本优势种AC值半矩阵 根据共同出现百分率PC半矩阵图(图3)可得,PC=0的种对数为29对,占总对数的9.67%,表明物种间趋向于独立分布,对生境有着不同的要求,不会同时出现;PC>0.5的种对数为12对,占总对数的4%,这些种对间联结性较强,物种对生境要求相似,同时出现的概率比较大,说明彼此可以通过合理利用生境资源,实现和谐共处;0.2 图3 草本优势种PC值半矩阵 3.3.3 种间相关性分析 Spearman秩相关分析对于数据的要求没有Pearson相关分析严格,数据不需要服从某一种分布类型[22],适合分析物种间的相关性[23],故本研究采取Spearman秩相关分析研究草本优势种的种间关系。根据Spearman秩相关分析可得(表4),正相关的种对有151对,占总对数的50.33%,其中显著正相关的有10对,极显著正相关的有9对,共占总对数的6.33%;负相关的种对数有149对,占总对数的49.67%,其中显著负相关的有9对,极显著负相关的有5对,共占总对数的4.66%,可知呈现显著相关和极显著相关的对数仅有33对,大部分种对间的相关性不显著,优势种间联结比较松散,各自趋向于独立分布,Spearman秩相关分析与χ2检验结果存在略微差异,这可能因为Spearman秩相关分析是以物种重要值为原始数据,对种间关联性进行定量的表达,由此导致了结果的略微偏差[24]。 表4 Spearman相关系数检验半矩阵 生态种组是指由对生境要求相似的物种组成的种组。根据Spearman秩相关系数结果,对25个草本植物优势种进行聚类分析(图5),结合组内各种对间的正关联关系和各种的生态学特性[25-26],将其划分为5个生态种组。 生态种组Ⅰ:包括披针叶薹草(12)、葛缕子(22)、野菊(14)、铁杆蒿(4)、北柴胡(20)、茭蒿(1),组内各优势种间呈正关联,其中呈现极显著的有6对,显著的有1对,表明物种具有相似的生态特性,均为旱生植物,多生于极其干燥的硬土石山坡和土坎,抗旱能力极强,多出现于群落演替后期的群落。 生态种组Ⅱ:包括拂子茅(13)、委陵菜(21)、白羊草(8)和蓟(23),种对关联为正关联,呈现极显著正关联和显著正关联的均为1对,这些物种均为中生植物,抗旱能力相对比较弱,多生长于土壤条件相对比较好的土坎。 生态种组Ⅲ:包括长裂苦苣菜(18)、野豌豆(19)、茵陈蒿(2)和菊叶香藜(17),种对间均为正关联,呈现极显著关联和显著关联的分别为1对和2对,这些物种对生长环境的需求相似,存在一定的依存关系,其常出现于演替早期阶段的群落。 生态种组Ⅳ:包括阿尔泰狗娃花(7)、茜草(11)、芦苇(10)和针茅(9),组内各物种间的关联性均为不显著正关联,联结性弱,该组物种根系相对比较发达,多见于土质条件较差的干燥土坎上。 生态种组Ⅴ:包括异叶败酱(6)、刺儿菜(25)、赖草(3)、硬质早熟禾(5)、小冠花(15)、蛇莓(16)和艾蒿(24),组内各优势种间呈正关联,但物种之间的联结性偏弱,本组植物主要为土坎经过扰动后,3~5 a生长发育的植物,多见生长在土质较为疏松的土坎上。 黄土区土坎植被群落25个草本优势种总体关联性表现为显著性正关联,这与陈文思等[12]对陕北黄土区自然恢复的植物群落优势种种间关系研究结果一致,说明土坎的干旱环境筛选出适合该区生境的植被种类,成为该区域的优势种,在群落中得以共存和协调发展。种间总体关联性反映了环境和植物群落的统一,这主要体现在群落种类组成上,由于调查区土坎生境条件比较恶劣,土壤贫瘠,水土流失严重,故生长的物种大部分都是耐旱植物种类。 图4 25个草本物种的Spearman秩相关系数聚类图 种对间χ2检验结果表明,黄土区土坎植被群落25个草本优势种组成的300个种对中,正关联的种对163对,负关联的种对135对,其中显著关联和极显著关联的种对数占总种对数的6.67%,大部分种对间关联性不显著,说明不同种类的物种间相互依赖程度比较低。其中,茭蒿在调查区域中出现的频率最高,是研究区域的绝对优势物种,与13个物种呈现正关联,与11个物种呈现负关联。茭蒿与铁杆蒿、阿尔泰狗娃花等物种呈现正关联,主要是因为这些物种为地带性植物种类,适应能力比较强,为旱中生或中生植物,多见于土石干燥山坡;茭蒿与赖草、茵陈蒿等物种呈现负关联,这主要是因为赖草和茵陈蒿常出现于受到一定程度干扰的样地,茭蒿常出现于演替后期未受到人为干扰的样地中。 在关联度指数计算中,联结系数AC表现为联结性较强(AC>0.5)的种对数为8对,共同出现百分率PC表现为联结性强(PC>0.5)的种对数为12对,其余大部分种对间的联结性都比较弱。2种计算方法中硬质早熟禾和拂子茅、野菊和北柴胡联结性都比较高,这主要是由于物种的生物学特性相近,对生境要求相似,故同时出现的概率比较大。在进行种间关联性测定时,若只使用联结系数AC和共同出现百分率PC中的一种,会存在一定的局限性。如:联结系数AC结果显示茭蒿和阿尔泰狗娃花联结性弱,共同出现百分率PC则表现为联结性强,茭蒿是土坎植被群落恢复中的优势物种,频度较大,阿尔泰狗娃花在样地内的数量比较少,但是在各样地中出现的频率比较高,同时与茭蒿具有类似的生态特性,即耐旱、适应性强等,故两者同时出现的概率比较高,表现为正关联,这也与实际调查结果相一致,因此可以将这些方法的检验结果相互补充,得到更精确的种间关联程度。 通过种间χ2检验和种间Spearman秩相关系数的种间关系分析,均得出草本优势种间正关联的种对数高于负关联的种对数,正负种对比值>1,说明大部分群落在物种组成上比较相似。但两者的部分计算结果有出入,其原因在于种间关联χ2检验是根据物种在样地中出现与否的二元数据来分析种对间的关系,反映了物种的存在与否,但缺乏物种的数量信息,而Spearman秩相关分析是根据物种重要值,从数量特征来分析种对间的关系[19]。种间关联χ2检验测定茭蒿和芦苇为正关联,而Spearman秩相关分析得出茭蒿和芦苇呈现负相关,这主要是因为芦苇出现的样地频率高,但是在样地中的数量比较少,所以在χ2检验中呈现正关联,而在Spearman秩相关分析中呈现负相关。由此可见,Spearman秩相关分析能够避免因为样地中稀少种的出现对种间关系造成误判,所以在研究中将两者结合起来,可以更为准确地揭示种间的关系。 在一定的环境条件下,植物群落的组成具有一定的规律性,并不是杂乱无章的堆积。根据种间Spearman秩相关系数和物种的生态特性,使用聚类分析法将25个草本优势种划分为5个生态种组,同一生态种组内,大部分物种对资源和环境有着相似的需求,在黄土高原护坎、护坡植物选择和群落构建时,应根据环境条件,以生态种组为依据,选取植物种类构建群落类型,其有助于土坎植被群落的稳定性和固土效益的持续发挥。2.3 数据处理
3 结果与分析
3.1 物种组成与优势种重要值

3.2 总体关联性分析

3.3 种对间关联性分析



3.4 生态种组的划分
4 结论与讨论