大兴安岭北部森林群落结构及植物多样性特征研究
2021-04-08崔佳佳
崔佳佳,铁 牛
(内蒙古农业大学 林学院,内蒙古 呼和浩特 010019)
大兴安岭地区是我国唯一的寒温带地区,是我国重要的森林植被带,其森林群落结构和物种多样性有许多独特之处[1]。但是由于多年采伐使用,对森林生态系统产生较大影响,而生态系统功能的基础是群落结构,决定了植物物种在群落的生态位和竞争机制,能够预测群落的演替进程和发展趋势,对于揭示群落维持稳定具有重要意义[2]。且已有研究表明,植物多样性影响着大兴安岭的群落稳定性[3-4],但是,多样性与稳定性的关系并非呈简单的线性关系[5]。因此,为了更好地认识大兴安岭北部森林群落组成、变化和发展,本研究利用α物种多样性和谱系结构来计算大兴安岭北部过伐林与原始林的多样性指数和植物物种间的系统发育距离[6],结合植物多样性讨论群落的构建机制[7-9],揭示其构建过程是理解多样性格局形成机制的重要途径[10],进而了解在大兴安岭北部地区过伐林与原始林之间的植物关系及生长状况,对该地区兴安落叶松林群落结构调整及经营与管理提供系统的科学理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于内蒙古大兴安岭重点国有林管理局根河林业局潮查林场境内的内蒙古大兴安岭森林生态系统国家野外科学观测研究站(以下简称大兴安岭森林生态站),地理坐标121°30′-121°31′E,50°49′-50°51′N;该区域属寒温带大陆性季风气候,冬季严寒而漫长,夏季凉爽且多雨,四季和昼夜温差大,年平均气温-5.5℃,极端最低气温-49.6℃(1月),极端最高气温35.4℃(7月),林区海拔高一般为500~900 m;坡度一般为5~20°,土壤多为暗棕色针叶林土,属于大兴安岭植物区系。乔木树种较为单一,主要有兴安落叶松(Larixgmelinii)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana)等;灌木树种主要有兴安杜鹃(Rhododendrondauricum)、越橘(Vacciniumvitis-idaea)、山刺玫(Rosadavurica)、杜香(Ledumpalustre)等;草本主要有大叶章(Deyeuxialangsdorffii)、野豌豆(Viciasepium)和沙参(Adenophorastricta)等。
1.2 研究方法
1.2.1 数据来源及处理方法 于2018-2019年7-8月大兴安岭森林生态站进行调查,在坡度坡位、林型与海拔等立地条件基本一致的前提下,选择3块过伐林样地(40 m×50 m)和1块原始林样地(235 m×50 m)进行调查(表1)。在调查样地内,乔木样方调查种类、株数、胸径和树高冠幅等,DBH>5 cm为乔木层,其中DBH<2.5 cm标记为更新层;在过伐林样地内随机选择3~4个5 m×5 m小样方(共11个),原始林样地内随机选取11个5 m×5 m小样方对灌木和草本进行调查,灌木样方调查其种类和株数;草本样方记录植物种类,同时按照株数估计每个种的相对多度。
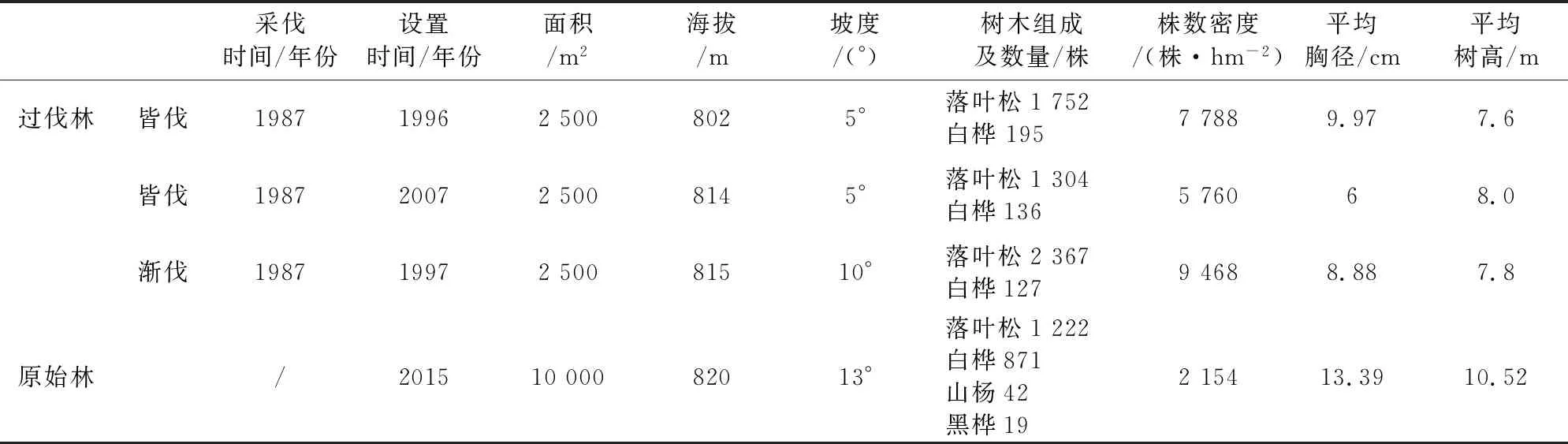
表1 样地基本情况Table 1 Basic situations of sample plots
1.2.1.1 谱系结构的计算 物种名参考《中国植物志》英文版。使用R语言结合群落物种组成构建谱系树,自动生成基于被子植物分类系统APGIII且具有进化时间的谱系树(phylomatic tree,version)[11]。并计算群落谱系结构,公式如下:
净谱系亲缘关系指数:NRIsample=-1×(MPDsample-MPDrandsample)/sd(MPDrandsample)
净最近种间亲缘关系指数:NTIsample=-1×(MNTDsample-MNTDrandsample)/sd(MNTDrandsample)
式中,NRI和NTI为净种间亲缘关系指数和净最近种间亲缘关系指数,MPD与MNTD为样方物种平均谱系距离和平均最近谱系距离。下角标sample为实际观测值,randsample代表随机999次组合计算后获得的期望值,sd为标准差。当NRI/NTI>0时,群落谱系聚集,群落中亲缘相似的物种趋向于聚集;当NRI/NTI<0时,群落谱系发散,群落中亲缘关系疏远的物种趋向于聚集;当NRI/NTI=0时,群落谱系随机。
以上数据通过R 3.5.2软件的picante软件包计算。
1.2.1.2 物种多样性测度 物种丰富度指数Petrick指数(R):R=S
物种多样性指数Simpson指数(λ):
物种多样性指数Shannon指数(H):
物种均匀度指数Pielou指数(Ep):
重要值(IV):IV=(相对高度+相对盖度+相对密度)/3
式中,S为每一样方内的物种总数;N为S个种的相对盖度之和;Ni为第i个种的相对盖度。所有样方的平均值为研究区域物种多样性指数。
以上数据获得利用R3.5.2软件的vegan包和ggplot2包计算并作图。
2 结果与分析
2.1 大兴安岭北部过伐林与原始林植物多样性
2.1.1 过伐林与原始林物种组成
2.1.1.1 过伐林物种组成及灌草层植物重要值 在研究区过伐林样地内调查共记录物种数36种(表2),乔木层只有落叶松和白桦2种。草本层物种多于灌木层,其中灌木层13种,隶属被子植物5科10属12种,裸子植物1科1属1种,占所调查物种总数的36.11%;草本层21种,隶属被子植物14科17属20种,蕨类植物1科1属1种,占所调查物种总数的58.33%,则乔木层物种占所调查物种总数的5.56%。
对灌草层重要值进行统计,结果表明灌木层中杜香和越橘的重要值达到15%以上,且2物种重要值之和>40%,其相对高度、相对盖度、相对频度均高于其他物种,为研究区域的优势种;灌木层的杜鹃、笃斯越橘(Vacciniumuliginosum)和草本层的小叶章(Deyeuxiaangustifolia)和大叶章的重要值在 5% ~ 10%,属亚优势;灌木层山刺玫、金露梅(Potentillafruticosa)、柴桦(Betulafruticosa)、忍冬(Lonicerajaponica)和草本层裂叶蒿(Artemisiatanacetifolia)等5种植物重要值在1%~5%,属伴生种;灌木层的绣线菊(Spiraeasalicifolia)、茶藨子(Ribesnigrum)、刺蔷薇(Rosaacicularis)、灌木层西伯利亚刺柏(Juniperussibirica)和草本层独活(Heracleumhemsleyanum)等14种植物重要值都<1%,属偶见种。

表2 过伐林灌草层植物物种组成及重要值Table 2 Composition and importance values of plant species in shrub-grass layer plants of over cutting forest
2.1.1.2 原始林物种组成及灌草层植物重要值 调查共记录原始林27种植物,在乔木层分布最多的是落叶松和白桦,偶见山杨(Populusdavidiana)和黑桦(Betula_dahurica)。同过伐林相同,原始林草本层物种数多于灌木(表3),灌木层7种,隶属被子植物2科6属7种,占所调查物种数的25.93%;草本层16种,隶属被子植物12科15属16科,占所调查物种数的59.26%,则乔木层物种占所调查物种总数的14.81%。
通过计算原始林灌草层的植物重要值发现,灌木层杜鹃的重要值>20%高于群落其他物种,为研究区域优势种,灌木层的绣线菊、杜香、越橘和草本层鹿蹄草(Pyroladahurica)的重要值在5%~10%,属亚优势种,灌木层山刺玫、笃斯越橘、葎草叶悬钩子(Rubushumulifolius)和草本层裂叶蒿等6种植物的重要值在1%~5%,属伴生种。蒙古风毛菊(Saussureamongolica)等6种草本植物的重要值<1%,属偶见种。

表3 原始林灌草层植物物种组成及重要值Table 3 Composition and importance values of plant species in shrub-grass layer plants of primitive forest
2.1.2 α物种多样性 物种多样性可以度量一个群落结构和功能的复杂性,并反映群落及其环境的保护及恢复状况[12-14]。通过对过伐林和原始林的物种多样性指数的对比可知(图1),原始林的Shannon-Wiener多样性指数(图3-H)和Margalef丰富度指数(图3-D)大于过伐林的Shannon-Wiener多样性指数和Margalef丰富度指数,这一结果与大多数研究结果一致[15-16],说明过伐林在采伐过程中对原有的生态造成一定程度的破坏,故原始林内生物的多样化和变异性以及物种生境的生态复杂性优于过伐林。过伐林和原始林的Pielou均匀度指数(图3-E)和Simpson优势度指数(图3-P)之间相差不到0.1,说明过伐林与原始林的物种分配状况和物种在群落中的地位与作用一致。
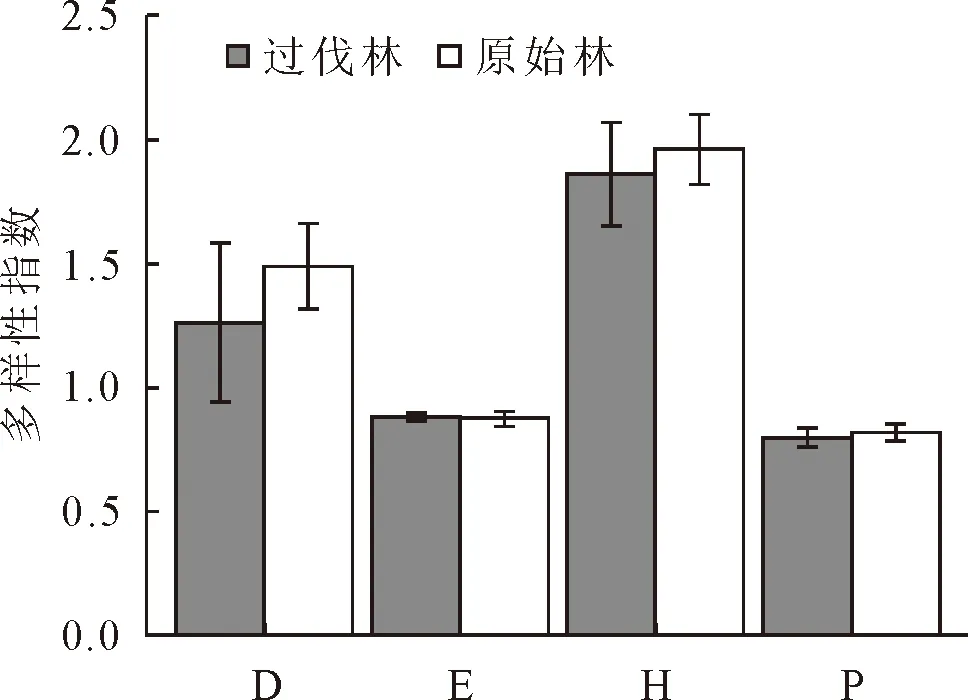
注:D-Margalef丰富度指数、E-Pielou均匀度指数、H-Shannon-Wiener多样性指数、P-Simpson优势度指数。
2.2 大兴安岭过伐林与原始林群落系统发育结构
2.2.1 谱系树 大兴安岭北部过伐林样地与原始林样地群落物种发育树(图1和图2),谱系树枝的长短表示物种发育所需要的时间(万年);分支表示物种间亲缘关系的远近。
白桦发育树的枝长小于兴安落叶松发育树的枝长,且白桦与其他物种之间的亲缘关系较近,兴安落叶松较远。这就解释了白桦(3.92万年)作为速生树在大兴安岭分布广泛,而兴安落叶松(266.72万年)作为大兴安岭的优势树种,要经过长时间的进化来适应环境。由图2可知,在原始林中除了树种落叶松和白桦外还有山杨,虽然山杨与其他物种的亲缘关系较近,但山杨(96.90万年)的生长慢于白桦,快于落叶松,但是对环境要求较高,故在过伐林鲜有山杨的存在。这就解释了在大兴安岭地区兴安落叶松是优势种,山杨是偶见种,白桦是速生种。

图2 大兴安岭北部过伐林植物谱系树Fig.2 Phylogenetic tree of subalpine meadow of over cutting forest in the Northern Greater Khingan Mountains
2.2.2 谱系结构
2.2.2.1 过伐林群落构建机制 过伐林的群落谱系结构(表4),种间平均进化距离与标准的差值(SD of MPD)、最近种间平均系进化距离与标准的差值(SD of MNTD)在过伐林调查区域的平均值分别为53.88和56.52,相差不大,原因可能是目前的群落处于发育中期,群落的物种组成与结构基本稳定。
过伐林样地内91%的NRI>0,NTI>0,因此在生态位理论中,生境过滤将会筛选出生态特征相似的物种共存,使共存的物种利用相似的资源,使得物种间的系统发育结构相对聚集,则过伐林在群落构建过程中环境过滤起主导作用。而样地P10的NRI<0,NTI>0,但已有研究群落谱系结构文献中NRI和NTI指数往往表现一致,这一现象与现有大多数研究结果不一致,无明显谱系结构,可能是这些样地中的竞争排斥与生境过滤作用相互平衡[17],且W.J.Kress等[18]和白家烨等[13]的研究也出现类似的情况,其解释认为植物科、属的重要值与谱系结构之间相关性较大,植物科、属重要值较大的样地谱系聚集,生态位优势较明显;而植物科、属重要值大小不明显的样地发散,生态位愈重叠,竞争排斥愈剧烈,从而导致群落的物种动态过程的不稳定。但在过伐林调查的11个样地内,只出现了1个样地NRI<0,NTI>0的情况,所以在整个过伐林中整体表现为聚集,说明在此群落构建过程环境过滤起主导作用。
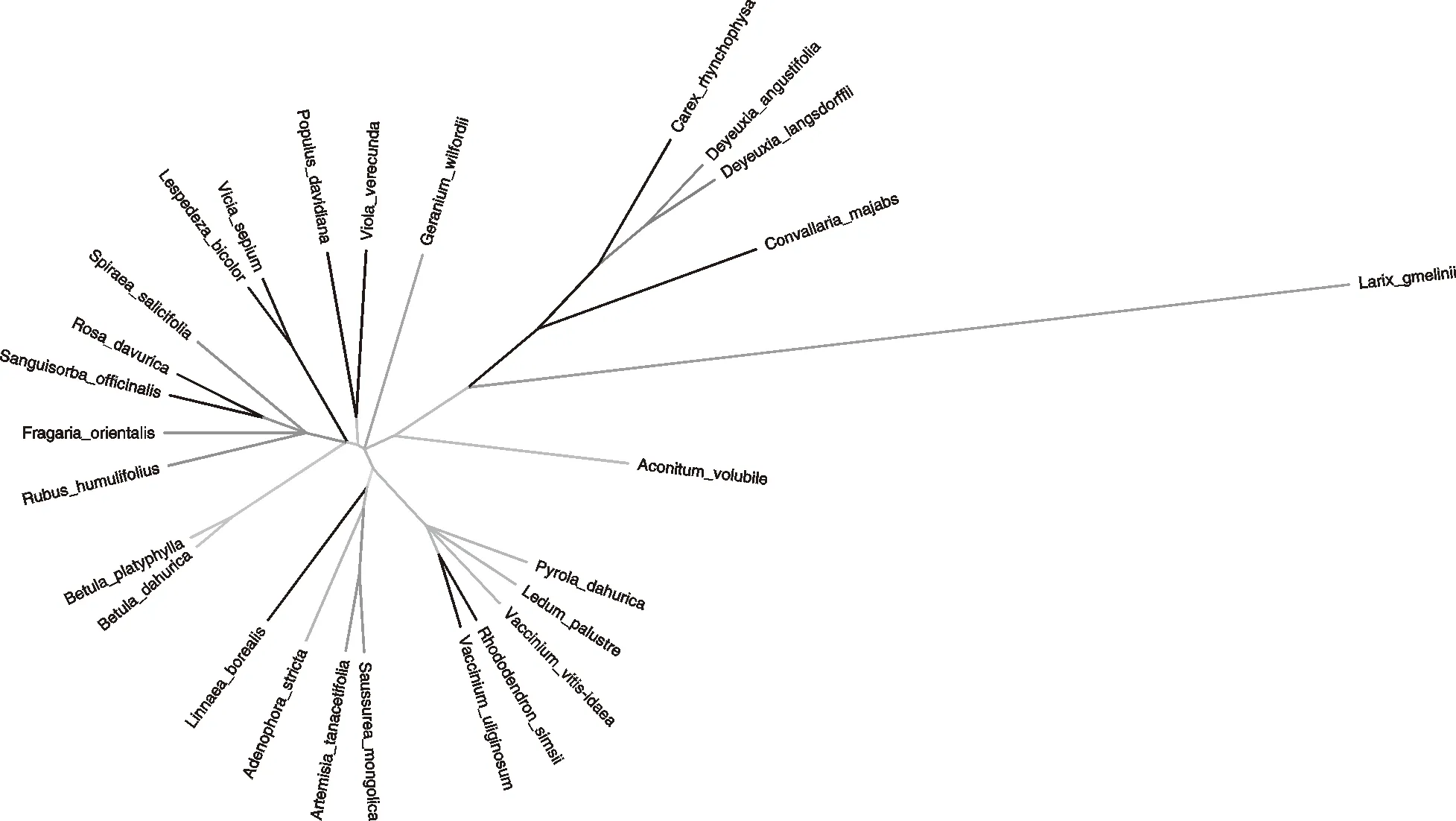
图3 大兴安岭北部原始林植物谱系树Fig.3 Phylogenetic tree of subalpine meadow of primitive forest in the Northern Greater Khingan Mountains
2.2.2.2 原始林群落构建机制 原始林的群落谱系结构(表5),种间平均进化距离与标准的差值(SD of MPD)、最近种间平均系进化距离与标准的差值(SD of MNTD)在原始林调查区域的平均值分别为49.34和77.62,相差较大,可能是目前群落处于发育末期,群落内的郁闭度增加、温度改变、土壤理化性质等环境的变化,渐渐对自身不利,新的物种迁入致使群落不稳定。
生态位理论认为,近缘物种通常利用较为相似的生态位,竞争排斥使群落中占有相似生态位的物种因竞争相同的资源而不能共存,群落整体系统发育结构相对发散;而中性过程认为,物种是没有差别,群落构建是随机的,群落发育结构也表现为随机。原始林样地64%的NRI>0,且NRI值均接近于0,这表现为原始林样地有轻微的发散,且趋向于随机分布;样地内9%的NTI<0,说明不同的物种竞争同一资源,使得物种的系统发育结构相对发散。在原始林调查区域中有4块样地(P01,P02,P03,P09)NRI<0,NTI>0,虽然亲缘关系近的物种比较聚集,但是物种间亲缘关系并不倾向于聚集在一起,因此原始林群系在物种形成过程中很有可能在初期由于地理条件及环境资源经过了竞争分化,后期经过生境过滤形成了目前的群系。
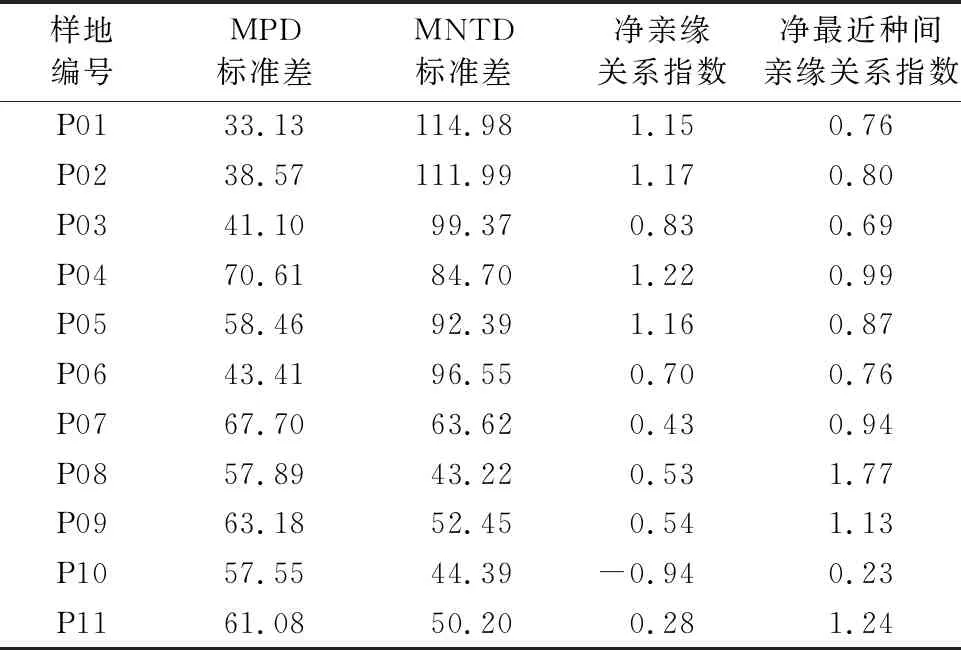
表4 过伐林SD of MPD、SD of MNTD、NRI和NTITable 4 SD of MPD,SD of MNTD,NRI and NTI of over cutting forest

表5 原始林SD of MPD、SD of MNTD、NRI和NTITable 5 SD of MPD,SD of MNTD,NRI and NTI of primitive forest
在原始林调查区域中P05的NRI>0,NTI<0,说明谱系结构较为聚集,种间谱系结构表现为发散,可能是环境过滤和负密度制约共同影响着群落的构建过程。在原始林群落中,谱系关系较近,但其物种之间聚集度并不高,而在整个群落中倾向于散落分布,因此原始林群系在物种形成过程中很有可能在初期经历了环境过滤,后期经历了负密度制约。随着物种的生长与数量的增加,可用资源有限,物种间的竞争加剧出现排斥现象,使亲缘关系最近的物种不再聚集生长而分散于整个群落,从而出现了NRI虽呈>0,但NTI却明显<0。
3 讨论
3.1 大兴安岭北部过伐林与原始林植物多样性
通过调查大兴安岭北部的过伐林和原始林不同植物群落发现,过伐林调查区域内的植物物种数较原始林多(表2和表3),造成这差异的原因可能是森林群落是动态变化的,森林是正向演替至顶级群落为止,而顶级群落的生产力及多样性都不及演替中的群落,原始林作为顶级群落,物种多样性少于演替中的群落过伐林,其林木间存在着竞争关系(表5),乔木冠幅较大,林内的光照强度、空气湿度、温度及土壤养分等得不到充分的利用,使原始林林下幼苗的天然更新较慢或部分死亡,致使原始林植物种数少于过伐林内植物种数,并且群落生长过程中植物物种的替代情况,是说明群落稳定的重要指标之一[17]。而物种多样性指数显示原始林内生物的多样化和变异性以及物种生境的生态复杂性优于过伐林,但物种分配状况和物种在群落中的地位与作用一致。这就验证了植物多样性影响着森林群落稳定性[3-4],但是,多样性与稳定性的关系并非呈简单的线性关系[5]。
3.2 大兴安岭过伐林与原始林群落系统发育结构
大兴安岭北部过伐林和原始林的群落谱系结构亲缘关系显示,其种间平均进化距离与标准的差值(SD of MPD)、最近种间平均进化距离与标准的差值(SD of MNTD)在调查区域的平均值相比,过伐林样地内的MPD标准差平均值大于原始林的MPD标准差平均值(表4和表5),造成这种差异的原因可能有:1)过伐林在群落构建的时间上少于原始林,故要达到进化鼎盛期所需要的时间多于原始林;2)生境过滤将会筛选出生态特征相似的物种共存,在过伐林研究区域内各物种之间群落聚集,只经历了环境过滤这一过程,而原始林研究区域内各物种在初期经历了环境过滤,后期经历了负密度制约,所以过伐林的种间进化距离与标准的进化距离的差值大于原始林。过伐林样地内的MNTD标准差平均值小于原始林MNTD标准差平均值,这说明原始林物种种间可能存在着较明显的竞争关系,另一种可能是负密度制约使原始林调查区域的最近种间平均进化距离与标准的差值(SD of MNTD)较大。
综上所述,在本研究中我们从物种多样性特征和群落构建机制2方面,对大兴安岭北部的过伐林和原始林进行研究,虽然对这2方面的研究较为详细,但更值得讨论的问题是:该如何对原物种的保护及利用,是否引入新的物种及引入方式;今后在对研究所在地区谱系的研究方法和手段上,可利用植物的DNA信息构建谱系树,并结合生态特征进行校正,从而精准高效的得出物种间的亲缘关系;可以利用数理统计模型和指数来精确最佳状态,利用统计分析提高解决问题的能力。
4 结论
通过研究发现,在植物多样性方面,原始林总体趋于稳定,但植物物种树少于过伐林植物物种数;在群落构建机制方面,过伐林表现出不断聚集的趋势,而原始林的聚集程度随着时间的增加而减小。上述结论说明大兴安岭北部原始林的植物多样性和群落构建在目前的环境背景下随时间逐步趋于单一化,因此根据相关研究结果表明,对该群落进行适当采伐来引进聚集度高(同一属)的植物以增加群落的稳定性,且偶见种对群落生态稳定起着不可或缺的作用;为调节林木间的关系及改变群落的动态变化,采伐原始林胸径和冠幅较大的老龄化乔木,使林下幼树及植被充分生长。采伐作业要遵循维持森林生境中的植物多样性原则,森林采伐期间要制定政策特别注意保留群落的关键生态结构,为启发森林管理者如何制定森林采伐策略。
