Adult human liver slice cultures: Modelling of liver fibrosis and evaluation of new anti-fibrotic drugs
2021-04-08DariaKartashevaEbertzJesinthaGastonLorianeLairMehiriPierrePhilippeMassaultOlivierScattonJeanChristopheVaillantVladimirAlexeiMorozovStanislasPolSylvieLagaye
Daria Kartasheva-Ebertz, Jesintha Gaston, Loriane Lair-Mehiri, Pierre-Philippe Massault, Olivier Scatton,Jean-Christophe Vaillant, Vladimir Alexei Morozov, Stanislas Pol, Sylvie Lagaye
Daria Kartasheva-Ebertz, Jesintha Gaston, Loriane Lair-Mehiri, Stanislas Pol, Sylvie Lagaye, Institut Pasteur, Immunobiologie des Cellules Dendritiques, INSERM U1223, Paris 75015, France
Daria Kartasheva-Ebertz, Jesintha Gaston, Loriane Lair-Mehiri, BioSPC, Université de Paris, Paris 75005, France
Pierre-Philippe Massault, Service de Chirurgie digestive, Hépato-biliaire et Endocrinienne, APHP, Groupe Hospitalier Cochin, Paris 75014, France
Olivier Scatton, Jean-Christophe Vaillant, Service de Chirurgie digestive et Hépato bilio pancréatique, AP-HP, Groupe Hospitalier La Pitié-Salpétrière, Medecine Sorbonne Université, Paris 75013, France
Vladimir Alexei Morozov, Center for HIV and Retrovirology, Department of Infectious Diseases, Robert Koch Institute, Berlin 13353, Germany
Stanislas Pol, Département d'Hépatologie, AP-HP, Groupe Hospitalier Cochin, Université de Paris, Paris 75014, France
Abstract BACKGROUND Liver fibrosis can result in end-stage liver failure and death.AIM To examine human liver fibrogenesis and anti-fibrotic therapies, we evaluated the three dimensional ex vivo liver slice (LS) model.METHODS Fibrotic liver samples (F0 to F4 fibrosis stage according to the METAVIR score) were collected from patients after liver resection.Human liver slices (HLS) were cultivated for up to 21 days.Hepatitis C virus (HCV) infection, alcohol (ethanol stimulation) and steatosis (palmitate stimulation) were examined in fibrotic (F2 to F4) liver slices infected (or not) with HCV.F0-F1 HLS were used as controls.At day 0, either ursodeoxycholic acid (choleretic and hepatoprotective properties) and/or α-tocopherol (antioxidant properties) were added to standard of care on HLS and fibrotic liver slices, infected (or not) with HCV.Expression of the biomarkers of fibrosis and the triglyceride production were checked by quantitative reverse transcription polymerase chain reaction and/or enzymelinked immunosorbent assay.RESULTS The cultures were viable in vitro for 21 days allowing to study fibrosis inducers and to estimate the effect of anti-fibrotic drugs.Expression of the biomarkers of fibrosis and the progression to steatosis (estimated by triglycerides production) was increased with the addition of HCV and /or ethanol or palmitate.From day 15 of the follow-up studies, a significant decrease of both transforming growth factor β-1 and Procol1A1 expression and triglycerides production was observed when a combined anti-fibrotic treatment was applied on HCV infected F2-F4 LS cultures.CONCLUSION These results show that the human three dimensional ex vivo model effectively reflects the in vivo processes in damaged human liver (viral, alcoholic, nonalcoholic steatohepatitis liver diseases) and provides the proof of concept that the LS examined model permits a rapid evaluation of new anti-fibrotic therapies when used alone or in combination.
Key Words: Human liver fibrosis; Hepatitis C virus; Alcoholic liver disease; Nonalcoholic steatohepatitis; Ex vivo model; Drugs
INTRODUCTION
Forty five percent of deaths in the developed countries may be attributed to fibroproliferative diseases[1].Liver fibrosis is frequently associated with viral infection [Hepatitis C virus (HCV) and Hepatitis B virus (HBV)] infection, chronic inflammation, and excessive alcohol consumption.Despite effective antiviral treatment, morbidity and hepatitis-related mortalities are still increasing.Moreover, the number of non-viral liver diseases such as nonalcoholic steatohepatitis (NASH) and alcoholic liver disease (ALD) is steadily growing[2].
Progression to liver fibrosis is a multistep process, whose development time varies.Fibrosis is initiated by the activation of hepatic stellate cells triggered by several signaling pathways[3].The activation of stellate cells induces cellular matrix production and collagen 1 expression This process is stimulated by transforming growth factor-β1 (TGF-β1), which is a crucial element involved in fibrogenesis[4].The progression of liver fibrosis frequently results in cirrhosis (liver acini are substituted by regeneration nodules surrounded by fibrosis) and, further on, in the development of hepatocellular carcinoma.Liver fibrosis can persist even with effective treatments.In most cases, the necro-inflammation leading to fibrosis can be effectively treated by treatments with antiviral drugs that target HCV, by nucleoside analogs in patients with HBV, by immune suppression in autoimmune hepatitis, by ethanol weaning and other dietary approaches in ALD and NASH, and iron chelation for hemochromatosis.However, if patients are not treated in a timely manner, and fibrosis progresses to decompensated cirrhosis, the only remaining option is liver transplantation The main obstacles (or delays) to liver transplantation are an insufficient number or a shortage of suitable organs, long waiting lists and high cost of this procedure[5].Thus, mortality remains high in patients on the waiting list and new anti-fibrotic agents and new clinical strategies to manage patients in the different stages of liver fibrosis are needed.
The liver slices (LS) cultures are appropriate models to study liver fibrosis, because they maintain the complex cellular interactions that occurin vivo, which cannot be obtained in co-cultures systems[6].These cultures can be used to study molecular biological events either in the fibrotic liver tissue or in hepatocellular carcinoma tissue.Although the LS cultures from non-fibrotic and fibrotic rat livers have been used to investigate the early and late phases as well as the resolution of liver fibrosis[7,8], the experiments are limited to 3 days in the rat model[7-9], and to 15 days in the human non-fibrotic LS model[10].In previous studies, we developed a three dimensional (3D)ex vivomodel of HCV replication using human LS cultures that were followed for 10 days[11]to evaluate a new antiviral drug[12].
Here, for the first time, human fibrotic LS cultures (stages F2-F4) were successfully maintained and evaluated for 21 days.Using theex vivoLS model for a 21-d period makes it possible to explore molecular fibrogenesis in more detail including the role of important factors such as HCV infection, ethanol (EtOH), or steatosis.Thus, this model can improve the understanding of the three of the main causes of liver injury in clinical practice[2].In addition, it was demonstrated that LS cultures are efficient instruments to study anti-fibrotic drugs and their combination[13,14].
This study provides the proof of concept that theex vivomodel of human LS culture can be used for the molecular evaluation of fibrosis and to perform follow-up studies of new anti-fibrotic drugs and therapies for a 21-days period.
MATERIALS AND METHODS
Patients and human liver tissue specimens
Adult human liver tissue samples were obtained from selected patients with different liver pathologies, as previously described[11,12].Written informed consent was obtained from each patient included in the study and the study protocol conforms to the ethical guidelines of the 1975 Declaration of Helsinki as reflected in a priori approval by the institution's human research committee.Experimental procedures were carried out in accordance with French laws and Regulations and ethic committees from Pitié-Salpétrière Hospital, Cochin Hospital, and Pasteur Institute (France).The tissue samples from twenty patients were divided into three groups according to their METAVIR score[15].Liver samples were either non-fibrotic F0-F1, obtained during surgery for colorectal cancer liver metastases or fibrotic ranging from F2 to F4 according to the METAVIR score (Table 1).Significant necrotic inflammation as defined by an activity grade (A) was not always available.
Liver slices preparation, culture and infection
We obtained between 32 to 48 liver slices for each donor sample.On the different days of the kinetic experiments, the results were obtained from the mean of three liver slices from each donor.The liver slices were infected with a same viral stock.The liver slice cultures were inoculated with viral supernatant diluted in fresh medium, at MOI = 0.1 (multiplicity of infection) and incubated overnight at 37 °C.In order to remove free virus, the slices were washed three times with phosphate-buffered saline (PBS) and fresh complete culture medium was added, after which cultures were followed in the absence of additional changes to the media composition or replacement with fresh culture medium.The preparation and culture of the liver slices, HCV RNA transfection, virus production, HCV RNA extraction were performed as previously described[11,12].

Table 1 METAVIR scores and description of clinical liver samples
Liver slices viability
Special attention was paid to the condition of clinical liver samples.It is evident that the condition of the liver sections that we obtained was different.Thus, they were carefully selected forin vitrostudies.In fact, cell viability was estimated by determining the percentage of viable cells upon microscopic examination 10X, using live/dead fixable dead cell stain kit (Molecular Probes, Invitrogen, ThermoFisher, France) and, as the percentage of ATP production determined by enzyme-linked immunosorbent assay (ELISA) assays, while observing the increasing albumin and urea secretion levels throughout the experiments, which indicates that the physiological and biochemical parameters of the liver slices are normal.On day 15, the immunostaining for Ki67, a cellular marker for proliferation confirmed the cell viability.Only slices with viability greater than 80% were used and allowed to obtain all the presented results.The architecture of human LS cultures was accessed by hematoxylin-eosin (HE) staining performed as following: Cryosections were washed with distilled water for 5-10 min and then stained for 8 min with hematoxylin, followed by a washing step with warm water at 30 °C for 10 min.After a short washing step with distilled water, the slices were counter-stained for 6 min with eosin.Washing was followed by dehydration steps in 2 min intervals in 50%, 60%, 70%, 80% and 90% of ethanol.
Experimental set up
The experimental set up was as follows (Figure 1).Non-infected liver slices obtained either from human non-fibrotic (F0-F1) or fibrotic (F2-F3, F4) liver resection and cut in 350 µm-thick slices (approximately 2.7 × 106cells per slice), were cultivated for up to 21 days either with or without HCV, ethanol (EtOH) (1 mmol/L, 5 mmol/L, 25 mmol/L) or palmitate (500 µmol/L).Liver slices were infected with hepatitis C virus infection from cell culture (HCVcc) supernatant [Con1/C3 (genotype1b)][16](MOI = 0.1) (INF LS) in presence or not either of EtOH (1 mmol/L, 5 mmol/L, 25 mmol/L) or palmitate (500 µmol/L).The different concentrations of EtOH were added on days 0, 5, 10, 15 during the kinetic studies.Palmitate (500 µmol/L) was added or not to non-infected and infected liver slices on days 0, 5, 10 and 15 of the kinetic studies.As previously described, infectivity (ffu/mL) was measured on days 1, 5, 10, and 21 post-treatment depending on the experiment[11,12].All experiments were performed in triplicate.All data were presented in relation to the percentage of viable liver slices in culture.Once the model was validated for the presence of “molecular fibrogenesis” defined as a significant increase in fibrosis biomarkers [TGF-β1, Hsp47, α-SMA,Procol1A1, matrix metalloproteinases 2 (MMP-2), MMP-9, and vascular endothelial growth factor (VEGF)], we evaluated the anti-fibrotic properties of two drugs, ursodeoxycholic acid (UCDA) (Sigma-Aldrich, Merck, Germany) and α-Tocopherol (Toco) (Sigma-Aldrich, Merck, Germany).UCDA (240 ng/liver slice) and / or α-Toco (170 ng/liver slice) were added to the culture media from day 0 and every day up to day 20 of the culture.The estimation of the triglyceride content was essential during the different kinetic experiments, since its accumulation in the cytoplasm of hepatocytes indicates cell metabolism disturbances, typical of non-alcoholic fatty liver disease[17].
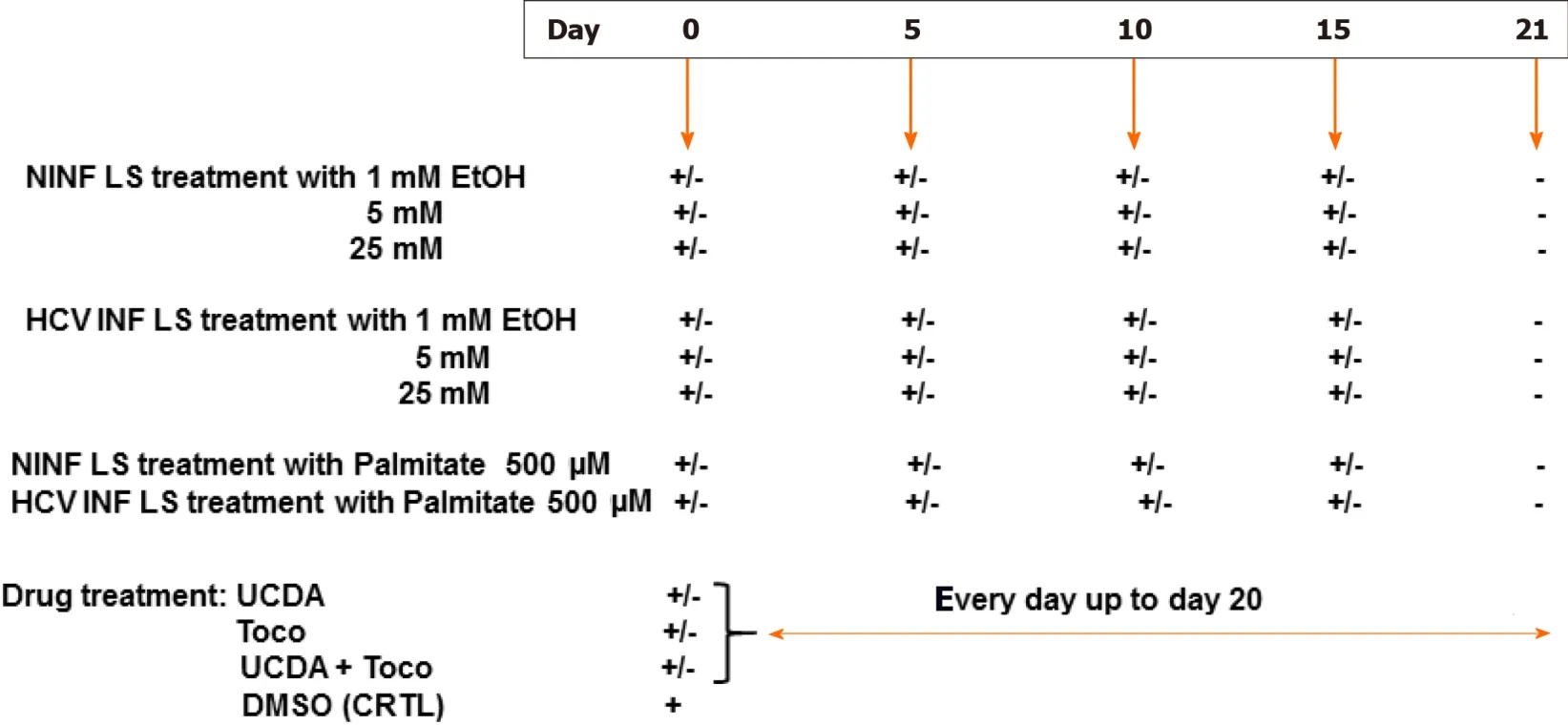
Figure 1 Experimental set up of the different liver slice treatments during the cultures.
Quantification of HCV RNA and liver-specific and fibrosis markers genes expression by real-time reverse transcription-quantitative polymerase chain reaction
The liver slices were washed three times in PBS at 4 °C.RNA was extracted from three combined slices using Trizol reagent as described in the protocol (Invitrogen, Cergy Pontoise, France).A strand-specific real-time reverse transcription-quantitative polymerase chain reaction (RT-qPCR) technique to quantify the intracellular levels of positive and negative-strand HCV RNA was performed during the experiments with the quantification of 28S rRNA used as an internal standard to quantify HCV in total liver RNA as previously described[11], (detection threshold: 25 copies/reaction).Briefly, reverse transcription was performed using an oligo primer and Moloney murine leukemia virus reverse transcriptase (Promega, Charbonnières, France) according to the manufacturer’s instructions.Real-time polymerase chain reactions were performed using Light CylclerR (Roche Applied Science, Grenoble, France) and FastStart DNA Master SYBR Green I kit (Roche Applied Science, Grenoble, France) according to the manufacturer’s instructions.
The relative expression of each liver-specific transcript (albumin, HNF-1β, HNF-4α transcription factors, cytochrome P450 enzymes, CYP2E1 and CYP3A4) was quantified by qRT-PCR and normalized to 18S RNA transcripts[11,12].The relative expression level of the transcripts was then determined in relation to the 18S RNA by the (Ct) method[13].The PCR conditions were as follows: Denaturation for 10 min at 95 °C, followed by 45 cycles of denaturation at 95 °C for 5 s, annealing at 60 °C for 20 s, and elongation at 72 °C for 30 s.The specificity of the PCR products was checked by a melting curve analysis after amplification.Primer sequences are listed in Table 2.
The expression of fibrosis markers in either non-infected liver slices (used as controls, CRTL) or in HCV-infected (INF) liver slices with or without the presence of EtOH or palmitate were evaluated by RT-qPCR with the SYBR PrimeScript RT-qPCR Kit (TaKaRa Bio Inc., Japan) and performed with the housekeeping gene, GAPDH as an internal control.Real-Time qPCR reaction for fibrosis markers including TGF-β1, heat shock protein 47 (Hsp47), alpha smooth muscle actin (β-SMA), procollagen1 A1 (Procol1A1), and VEGF was performed as follows: Denaturation for 10 min at 95 °C followed by 45 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C for 1 min, and elongation at 72 °C for 30 s.Concerning the MMP-2, MMP-9 gene expression, the Real-Time qPCR reaction was performed as follows: Denaturation for 10 min at 95 °C followed by 45 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C and 68 °C respectively, for 1 min, and elongation at 72 °C for 30 s.Ct (threshold cycle) values were corrected for the Ct values of the housekeeping gene GAPDH.Primer sequences are listed in Table 2.

Table 2 Primers used for real-time reverse transcription-quantitative polymerase chain reaction analysis
Albumin enzyme-linked immunosorbent assay: Human liver albumin concentrations were determined by a competitive ELISA as previously described[18,19].Purified human albumin and peroxidase-conjugated anti-human albumin were obtained from MP Biomedicals Europe (Illkirch, France).To ensure the specificity of the ELISA, human antibodies were incubated for 2 h at 37 °C with 3% BSA in 0.5% Tween-20 in PBS before the sample addition in order to block any cross reaction.
Urea assays:Urea concentrations were determined by colorimetric assay (640-1, Sigma-Aldrich) according to the manufacturer’s recommendations and analyzed with BioPhotometer 6131 (Eppendorf, Hamburg, Germany).
Western blotting and antibodies
Western blotting was performed as previously described[11,12], and the antibodies used are described as following.Mouse monoclonal antibodies (mAbs) to HCV core protein (C7-50; dilution 1/10000, Affinity BioReagents, Golden, CO, United States), HCV nonstructural protein 3 (clone1847, dilution: 1/2000, Viro-Stat, Portland, ME, United States) were used to analyze HCV expression, mouse monoclonal antibodies (mAbs) TGF-β1 (ab 190503, dilution: 1/2000, Abcam, United Kingdom), HSP-47 (M16.10A1, dilution: 1/1000, Enzo life sciences, France), Collagen I alpha 1 (NB600-450, dilution: 1/2000, Novus Biologicals, CO, United States), MMP-9 (ab119906, dilution: 1/2000, Abcam, United Kingdom), VEGF (ab69479, dilution: 1/2000, Abcam, United Kingdom), β-actin (A5316, dilution: 1/5000 Sigma-Aldrich, Merck, Germany), and rabbit polyclonal antibodies (rAbs) to MMP-2 (ab92536, dilution: 1/1000, Abcam, United Kingdom), alpha-smooth muscle actin [alpha-SMA (ab 5694, dilution: 1/2000, Abcam, United Kingdom)] allowing fibrosis analysis, were used as primary antibodies.Horseradish peroxidase-conjugated anti-mouse IgG and horseradish peroxidase-conjugated anti-rabbit IgG (Amersham, GeHealthCare Life Sciences, United Kingdom) secondary antibodies, taken 1:50000, were used as secondary antibodies.The reactions were developed using enhanced chemiluminescence detection reagents (ECL Advance kit, Amersham, GeHealthCare Life Sciences, United Kingdom), followed by exposure to X-OMAT film (Amersham, GeHealthCare Life Sciences, United Kingdom).
Histology and immunohistochemistry
Liver sections (7 µm) were stained with Goldner’s trichrome (Electron Microscopy Sciences, United States) or picrosirius red (Abcam, United Kingdom), performed standard protocols for collagen/connective tissue labelling using two slices per human liver sample and two different human liver samples per group.The images were taken with the EVOS XL Core Imaging System (Invitrogen, Thermo Fisher Scientific, France).The average integrated optical density (OD) of collagen deposition was calculated using the image quantification standard software, ImageJ2[20,21]or inform V2.1 (Perkin Elmer, MA, United States) used routinely in the histology (HISTIM) facilities (Cochin Institute, Paris, France).Immunostaining for TGF-β1 (mAbs, ab92486, Abcam, United Kingdom), MMP-9 (mAbs, ab119906, Abcam, United Kingdom), Ki67 (rAbs, ab15580, Abcam, United Kingdom), and alpha-SMA (rAbs, ab5694, Abcam, United Kingdom) was performed after paraffin removal in xylene, rehydratation in EtOH and then distilled water following the manufacturer’s instructions.Unmasking of the antigenic sites was performed at 120 °C in 10 mmol/L citrate buffer, pH 6.0.A solution of 3% H2O2was used to eliminate endogenous peroxidases.The sections were washed 3 times for 5 min.in TBS-Triton 0.1% solution.After incubation in a blocking solution (TBSTriton 0.1%-3% dry milk) for 1 h at room temperature, they were incubated with the primary antibodies.All primary antibodies were diluted at 1/50 in the blocking solution.After incubation for 2 h at room temperature, the sections were washed 3 times and incubated with secondary antibodies.The nuclei were stained with DAPI.All sections were counterstained with hematoxylin for tissue quality control.Control sections incubated with non-immune serum were used as negative controls.
TGF-β1 and Triglyceride quantification
TGF-β1 and triglyceride quantification were performed according to the manufacturer’s instructions (TGF-β1 Quantikine ELISA, RD Systems, United States; Triglyceride assays Kit–Quantification, ab65336, Abcam, United Kingdom).For TGFβ1, cellular lysates and culture supernatants were first treated with acid to lower the pH to 2.0, which denatures the latency-associated peptide and allows the detection of active TGF-β1.The supernatant was then brought back to neutral pH before the ELISA assays.
ATP production quantification and LDH assays
To check viability, the percentages of ATP was assessed at each point of the kinetics studies during the liver slices culture and determined by ELISA assays (CellTiter-Glo® 2.0 Assay, Promega, France)[19].The viability of liver slices and the potential cytotoxicity[20](cytoTox 96R Non-Radioactive Cytotoxicity Assay, Promega, France) induced by Ethanol, or Palmitate, or drugs treatments was estimated as described previously[11,12], in accordance with the manufacturers’ protocols.
Drugs inhibition of fibrosis markers expression and cytotoxicity assays
Human LS were infected or not with the HCVcc Con1/C3 supernatant as previously described[11,12].On day 0 of the culture, treatment either with (240 ng/Liver slice) UCDA or (170 ng/Liver slice) Toco or both (the recommended standard of care) or 0.5% of dimethyl sulfoxide (Sigma Aldrich, Merck, Germany) as a control, were added to HCV-infected or non-infected LS culture medium every day to day 20.TGF-β1 and Procol1A1 RNA expression were measured at different time points of the kinetic studies.All experiments were performed in triplicate.
Statistical analysis
Liver specimens from 20 individuals were examined.During the kinetic studies, the quantification of gene expression was determined in relation to the percentage of liver slice viability.The results were obtained from the mean of the three liver slices, on the different days of the kinetic studies.Statistical tests were performed using GraphPad Prism 8.0 software (GraphPad Software, La Jolla, CA, United States).Values are expressed as means ± standard errors of the mean.The data were compared using either the unpaired two-tailed Student’st-test or the two-way ANOVA test with multiple comparisons for a given day as compared to the standard LS.APvalue of 0.05 or less was considered significant.
RESULTS
Maintenance of phenotypic characteristics and viability of three dimensional human non-fibrotic (stages F0-F1) and fibrotic (stages F2-F4) LS cultures for 21 days of cultivation
The viability of human LS cultures during prolonged studies was and is a crucial factor.Liver slices viability (percentage of ATP production) and tissue morphology were assessed daily, until day 21.The architecture of the liver slices was normal (Figure 2A) and human liver slices (HLS) expressed the Ki67 protein, a proliferation marker (Figure 2B).Human LS cultures maintained their differentiation status throughout the entire study period, as previously described (Figure 2A-C)[10,12].Indeed, LS status was confirmed by analysing various parameters and biomarkers, in particular, albumin content, hepatocyte nuclear factors HNF-1β, HNF-4α, CYP2E1, and CYP3A4 (Figure 2D)[10-12,22-24].A comparison of the expression of hepatocytespecific genes in F0-F1 non-infected liver slices and Huh-7.5.1 cells showed increased expression in F0-F1 non-infected liver slices on day 21 compared to that in Huh-7.5.1 cells, either at an exponential growth phase or at the confluence (data not shown).CYP3A4 expression was undetected in Huh-7.5.1 cells whatever the growth stage[25].Albumin and urea secretion increased throughout, indicating that liver slices had retained normal physiological and biochemical parameters (Figure 2E-F)[11,12].As previously reported[11], the cell viability and expression of hepatocyte-specific genes were also evaluated post- HCVcc[11].Results were similar to those in uninfected liver slices, indicating that there was no evident cytopathic effect (Figure 3A-C).
The viability of non-fibrotic (F0-F1) and fibrotic (F2-F4) LS cultures and resistance to EtOH and palmitate treatments were tested during the 21 d follow-up studies by evaluating of the rate of ATP production in the liver slices (Figure 4A, C-E and G)[22], and by quantification of LDH release from liver slices (Figure 4B-F)[23].On day 21, the F0-F1 and F2-F4 non-infected liver slices had a viability rate of 75% and 50%, respectively (Figure 4A).Following treatment with 25 mmol/L of EtOH, ATP synthesis in F0-F1 infected liver slices was reduced by 55% on day 21 (Figure 4D), and tissue viability decreased by nearly 25% compared to untreated F0-F1 noninfected/infected liver slice cultures (Figure 4C and D).However, the addition of EtOH (25 mmol/L) did not change LDH release in F0-F1 and F4 non-infected LS cultures (Figure 4F).Treatment with palmitate (500 µmol/L) did not reduce significantly the viability rate of F0-F1 non-infected and infected LS cultures, compared to untreated and non-infected LS cultures (55% and 65%, respectively) (Figure 4G).There was no significant difference in LDH release from F0-F1, F2-F3, and F4 non-infected and infected LS cultures after treatment with the combination of UCDA and alfa-Toco (Figure 4H and I).Results of ATP production in F0-F1, F2-F3, and F4 non-infected and infected LS cultures after treatment with the combination of UCDA and alfa-Toco were significantly positive (Figure 4J-K) with increased ATP production in Fibrotic treated liver slices.These results, showing no significant changes in viability (with increasing levels of albumin, urea secretion as well as ATP production throughout the experiments) or morphology (Ki67 marker expression), confirm that the non-fibrotic (F0-F1) and fibrotic (F2-F4) LS cultures can survive for 21 days, and that the 3D LS cultures tolerated the different treatments (Figure 4H-K).Thus, LS cultures from selected donors can be used in extended research.
Evaluation of the expression of fibrogenesis liver biomarkers in 3D LS cultures from non-fibrotic (F0-F1) and fibrotic (F2-F4) livers
Activation or down-regulation of certain biomarkers reflects the process of the transition of the non-fibrotic liver to the fibrotic liver designated as the molecular fibrogenesis.We measured the expression of seven fibrosis biomarkers (TGF-β1, Hsp47, α-SMA,Procol1A1, MMP-2, MMP-9, and VEGF)[1,26]by RT-qPCR to analyse both non-fibrotic (F0-F1) and fibrotic (F2-F3, F4) stages of the liver in human LS cultures.
Induction of fibrogenesis by three exogenous factors: HCV, EtOH, and fatty acids (palmitate) in non-fibrotic (F0-F1) and fibrotic (F2-F4) LS cultures
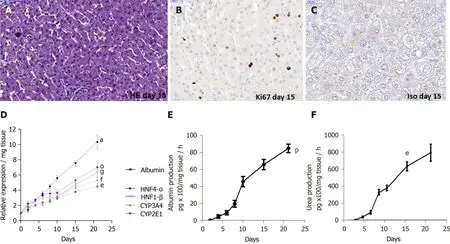
Figure 2 Maintenance of phenotypic characteristics of human non-fibrotic (F0-F1), and fibrotic (F2-F3, F4) liver slices during the culture, demonstrated by histochemistry, real-time reverse transcription-quantitative polymerase chain reaction and biochemical assays.
HCV efficiently replication in LS cultures: A model of the viral liver disease:Robust replication of HCVcc and production of infectious viral particles were detected up to day 21 in human F0-F1 LS (Figure 5).Intracellular replication of the viral genome was assessed by a strand-specific RT-qPCR, as previously described[11].The HCV RNA negative strand, proof of HCV genome replication, could be detected as early day 1 post-infection, and the intracellular levels of both negative and positive strands increased significantly during LS culture.These results confirmed active viral replication in LS cultures (Figure 5A).The HCV expression level was significantly increased in the LS culture on day 5 post-infection (Figure 5A).HCV protein expression was confirmed by Western blotting.Detection of core and nonstructural protein 3 proteins confirmed effective intracellular processing of the viral protein precursor[11](Figure 5B and C).
The virus titer was estimated in LS culture supernatants using a classic titration assay on Huh-7.5.1 cells to determine whether progeny virions released from the infected LS could replicate[11].Infectivity increased during the culture and reached a peak of up to 1.7 × 105ffu/mL respectively, by day 21 post-infection (Figure 5D).To further confirm that the new progeny virus produced by the human LS called the primary-culture-derived virus was indeed infectious, naive human LS were infectedde novowith primary-culture-derived virus Con1/C3 at MOI = 0.1.Ade novoproductive infection of LS was obtained with higher infectivity titers on day 21, genotype1b (180000 ffu/mL) (Figure 5E).Thus, HCV RNA replication, the expression of viral proteins, and the production of highly infectious particles were demonstrated.
HCVcc infection of non-fibrotic (F0-F1) LS activated the expression of the main profibrogenic markers.During follow-up studies, in non-fibrotic (F0-F1) LS cultures (Figure 6), RNA, and protein expression of TGF-β1 (Figure 6A-C), α-SMA, Hsp47, Procol1A1, (Figure 6D-F) had increased significantly in non-infected and infected LS on day 21.A marked 2.6 to 3.6 fold increase of α-SMA, Hsp47, Procol1A1 RNA expression was observed in non-fibrotic (F0-F1) HCV infected LS cultures, compared to non-infected LS cultures on day 21.MMP-2 RNA expression was also significantly increased after HCV infection in non-fibrotic F0F1 LS, (Figure 6G).On the contrary, there was no significant difference in MMP-9 RNA expression between F0-F1 noninfected and infected liver slices (Figure 6H).VEGF RNA expression increased irregularly up to day 21 and seemed to be influenced by HCV infection until day 5 compared to non-infected LS (Figure 6I).The triglyceride production increased in both F0-F1 non-infected and infected LS cultures (Figure 6J) with no significant difference between them.
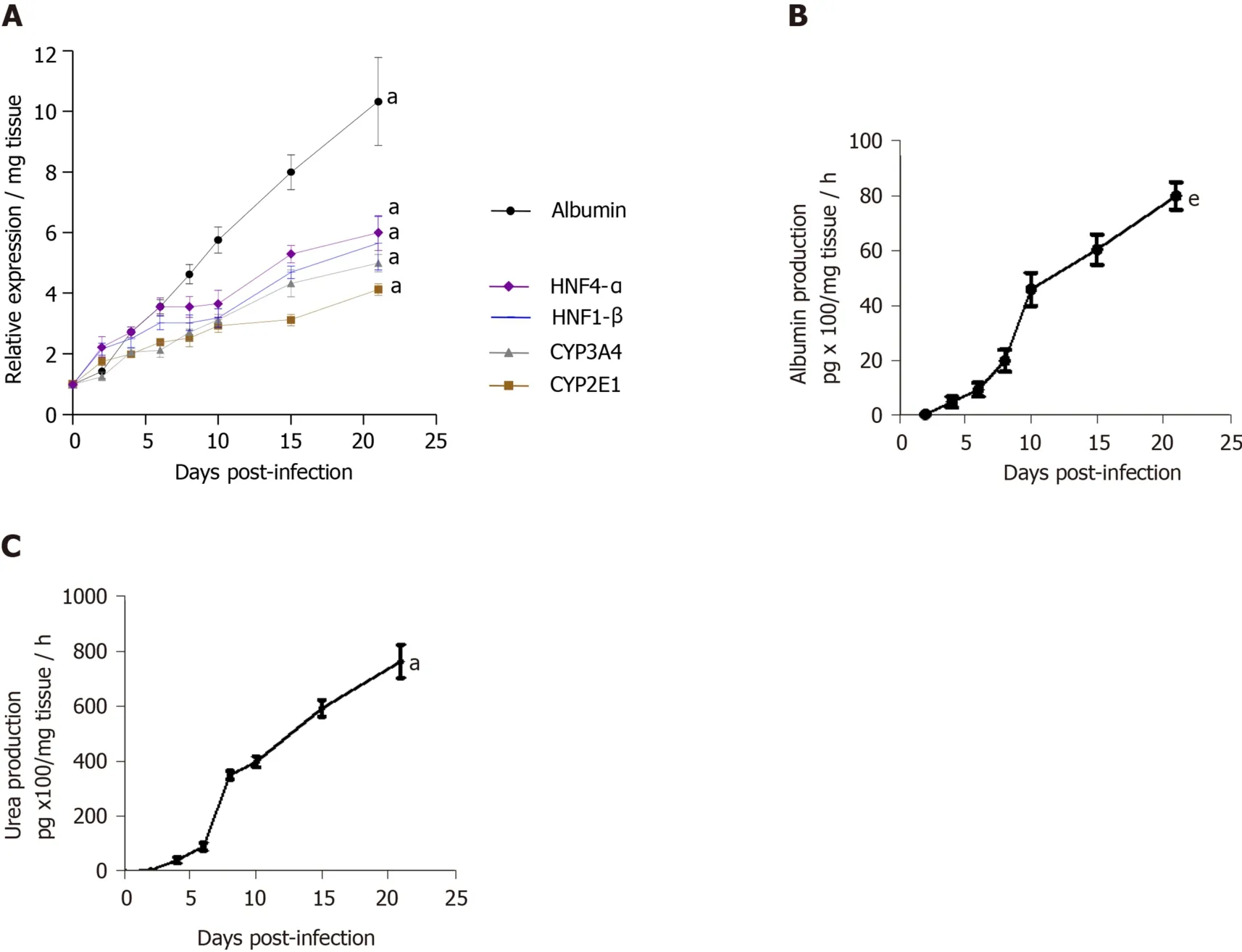
Figure 3 Maintenance of phenotypic characteristics of human non-fibrotic (F0-F1) hepatitis C virus-infected liver slices during the culture, demonstrated by real-time reverse transcription-quantitative polymerase chain reaction and biochemical assays.
Expression of fibrosis biomarkers was higher in fibrotic LS culture (stages F2-F3 and F4), than in non-fibrotic LS cultures, with a significant 4 to 8 fold increase compared to controls (day 1) (Figure 7).This mainly concerned TGF-β1 (Figure 7A), Procol1A1 (Figure 7C), α-SMA (Figure 7D), Hsp47 (Figure 7E) as well as an increased triglyceride production in fibrotic LS (approximately 3.2 fold) (Figure 7B).After day 10, RNA expression increased with the progression of fibrosis.MMP-2 RNA expression (Figure 7F), as well as MMP-9 and VEGF expression (Figure 7G and H), did not differ between fibrosis stages F2-F3 and F4.It is interesting to note that HCV infection significantly increased TGF-β1, Hsp47,α-SMA, Procl1A1,MMP-2, MMP-9, VEGF expression as well as triglyceride production in fibrotic (F2-F3, F4) infected LS cultures.A significant 2 to 4 fold increase in fibrosis biomarkers was observed on day 21 in F2-F3 and F4 HCV infected LS compared to F2-F3, F4 non-infected LS.Thus, the TGF-β1 (Figure 6A-C), α-SMA, Hsp47, Procol1A1, MMP-2, MMP-9, VEGF expression increased in non-infected and infected LS cultures with a greater increase in infected LS cultures than in controls.On day 21, a significant 2 to 13 fold increase in fibrosis biomarkers was observed in F2-F3, F4 infected LS cultures compared to F2-F3, F4 noninfected LS cultures.Triglyceride production increased in both non-infected and infected LS cultures, independent from the stage of fibrosis.After 21 d of the culture, the amount of triglyceride in the supernatant of F2-F3 and F4 LS cultures increased by 1.36 and 2.7 folds, respectively (Figure 7B).Increased expression of the TGF-β1, α-SMA, Procol1A1, MMP-2, MMP-9, and VEGF in F2-F3 LS cultures throughout the 21-d of follow-up was confirmed by Western blotting (Figure 7I and J).On day 10, immunohistochemistry showed that TGF-β1, α-SMA and MMP-9 expression (Figure 7K) was increased by about 20% in F2-F3 LS compared to day 0.

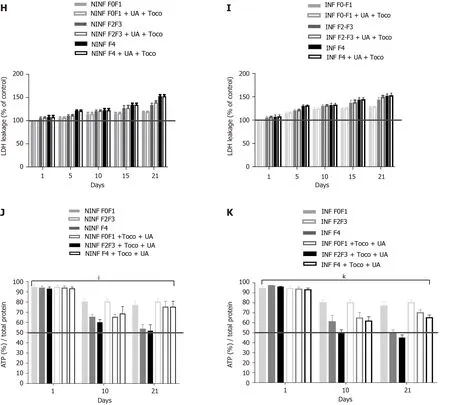
Figure 4 Viability of human non-fibrotic (F0-F1), and fibrotic (F2-F3, F4) non-infected or hepatitis C virus-infected liver slices during the different kinetic studies, with no treatment cytotoxicity as shown by ATP and LDH dosages.
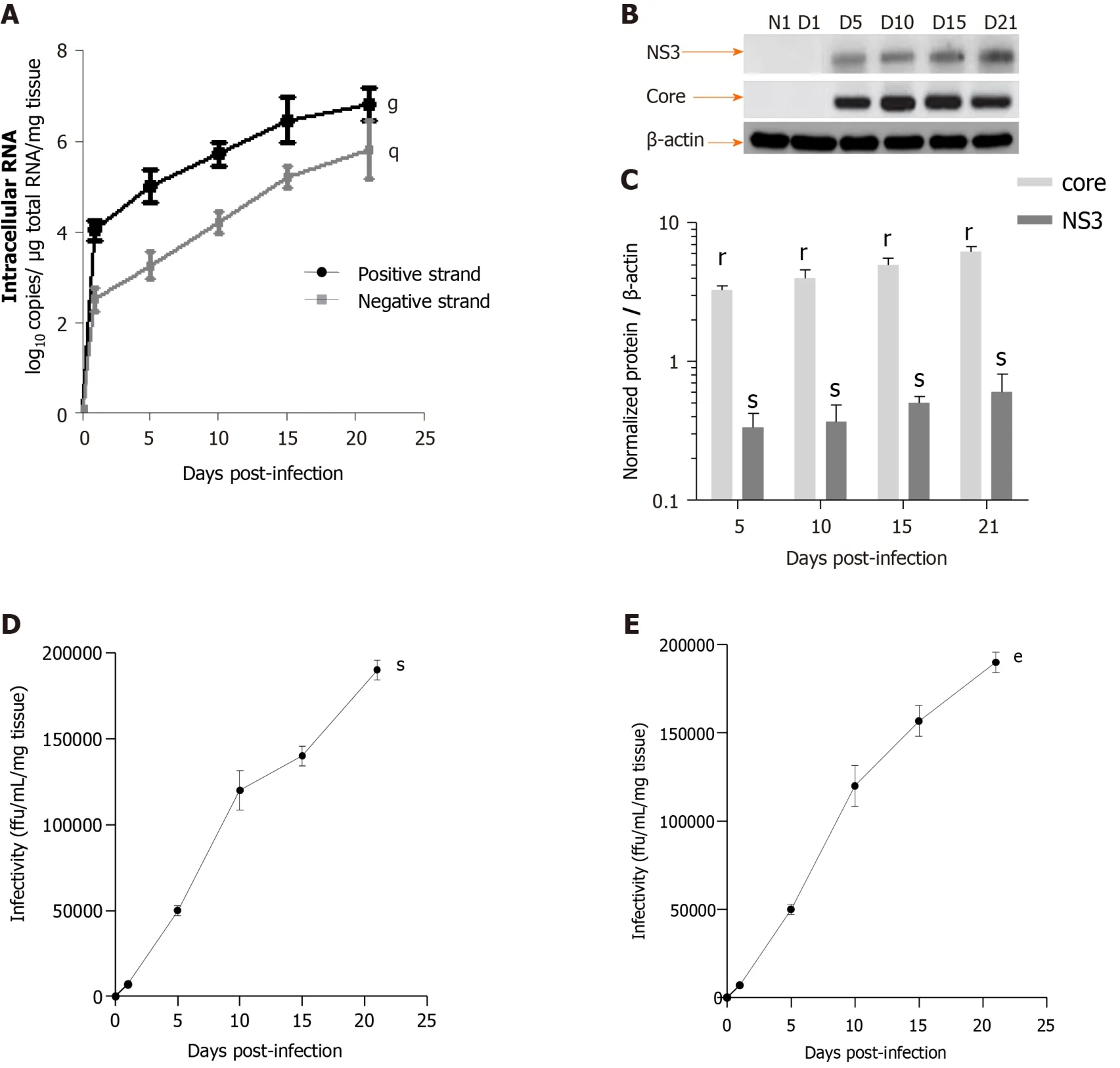
Figure 5 Efficient replication of hepatitis C virus RNA, and hepatitis C virus core and NS3 proteins expression in human F0-F1 liver slice culture as shown by real-time reverse transcription-quantitative polymerase chain reaction and western blotting analysis.
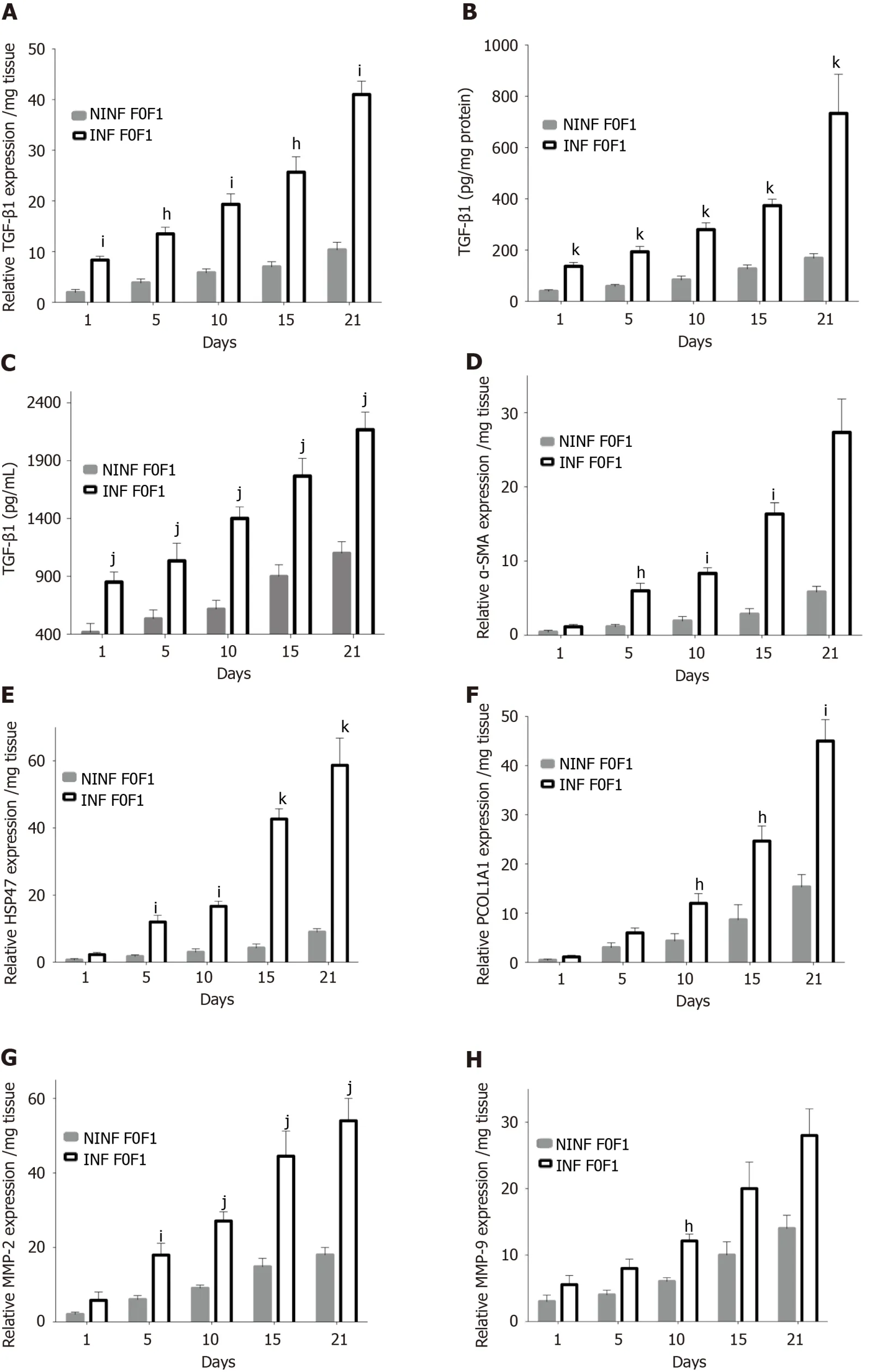

Figure 6 Real-time reverse transcription-quantitative polymerase chain reaction analysis evidencing the significant increase of fibrosis markers expression at the transcriptional level in human F0-F1 non-infected or hepatitis C virus infected liver slice during the kinetics.
Exposition of LS cultures to ethanol: A model of the alcoholic liver disease:The effect of EtOH exposure on LS cultures was estimated using non-fibrotic (F0-F1) HCV infected or non-infected LS cultures (Figure 8A-F and Figure 9) and fibrotic (F2-F4) HCV infected or non-infected LS cultures (Figure 8G-L) and Figure 10A-L).One mmol/L, 5 mmol/L or 25 mmol/L of EtOH was added to F0-F1 HCV infected, or noninfected LS cultures (Figure 8A-F).Only the highest concentration of EtOH was studied in fibrotic (F2-F3 and F4) non-infected or HCV-infected LS cultures, (Figure 8G-L and Figure 10A-L) respectively.During the follow-up studies (Figure 8A-F), EtOH enhanced the RNA expression of fibrosis markers in a dose-dependent manner in F0-F1 LS cultures.Increased expression of TGF-β1, Procol1A1 RNA was further detected in F0-F1 infected LS (Figure 8B, D and F, Figure 9B, Figure 10B), compared to noninfected LS (Figure 8A, C and E, Figure 9A, Figure 10A).Similar results were found in fibrotic F2-F4 LS (Figure 8G-L, Figure 10A-L).Interestingly, there was no significant increase in Procol1A1 or α-SMA RNA expression in F0-F1 non-infected LS except on day 21 when 25 mmol/L of EtOH was added to the culture (Figure 8E and Figure 9C, respectively).However, a significant dose-dependent increase of the Procol1A1 and α-SMA RNA expression occurred whatever the dose of EtOH added to F0-F1 infected LS cultures (Figure 8F, Figure 9D).There was a dose-dependent increase in the RNA expression of the other fibrosis markers such as α-SMA (Figure 9C and D; Figure 10E and F), and HSP47 (Figure 9E and F, Figure 10C and D) with the addition of EtOH in F0-F1 to F4 infected LS which was less marked in F0-F1 to F4 non-infected LS.Analysis of F0-F1 to F4 HCV non-infected or infected LS showed a significant dose-dependent increase in MMP-2, MMP-9, and VEGF expressions in response to EtOH (Figure 10GL).Masson’s trichrome staining showed a significant increase in collagen fibers (%) between day 1 (1.242% of collagen) and day 6 (2.076% of collagen) in F0-F1 HCV infected LS treated with 5 mmol/L of EtOH (Figure 8M) but not in F0-F1 non-infected LS with the same treatment (Figure 8N).Picro Sirius red staining confirmed the significant increase in collagen fibers (%) between day 1 (0.55% of collagen) and day 6 (1.53% of collagen) in F0-F1 HCV infected LS treated with 5 mmol/L of EtOH compared to non-treated LS (data not shown).
Exposition of LS cultures to palmitate: a model of NASH.To imitate NASH, nonfibrotic (F0-F1) LS cultures infected (or not infected) with HCV were exposed to 500 µmol/L of palmitate (Figure 11).More marked triglyceride synthesis was noted in F0-F1 palmitate treated HCV-infected LS cultures, than in F0-F1 untreated non-infected LS cultures (Figure 11A).The F0-F1 infected LS cultures treated with palmitate demonstrated more marked expression of the fibrotic markers such as TGF-β1 (Figure 11B), intracellular expression of TGF-β1 (Figure 11C), and secretion of the extracellular TGF-β1 (Figure 11D).A similar increase was observed with Procol1A1, α-SMA, and HSP47 (Figure 11E-G) on day 21.The expression of markers (RNA) involved in liver fibrolysis, (MMP-2, -9), and VEGF increased significantly in both F0-F1 non-infected LS cultures treated or not with palmitate (Figure 12A).But, the treatment of F0-F1 non-infected LS cultures with palmitate showed a greater significant increase of the expression of MMP-2, -9, and VEGF compared to those of F0-F1 untreated non-infected LS cultures.The treatment of the F0-F1 infected LS cultures with palmitate, increased significantly VEGF, MMP-2, and MMP-9 from day 10, 15, and day 21 respectively (Figure 12B).Fibrotic marker expression increased both in F0-F1 LS cultures HCV infected or non-infected treated with palmitate but with a greater increase in F0-F1 HCV infected LS treated with palmitate.


Figure 7 Real-time reverse transcription-quantitative polymerase chain reaction analyses of RNA expression of liver fibrosis markers (TGF-β1, Procol1A1, α-SMA, HSP47, MMP-2, MMP-9, VEGF), and triglyceride production in non-infected or hepatitis C virus infected liver slice cultures from fibrotic liver (F2-F3, F4) showing a significant increase during the kinetics.
LS treatment with a combination of the “hepatoprotective” UCDA and anti-fibrotic α-Toco drugs significantly reducing the expression of the main fibrosis markers TGFβ1, Procollagen1A1, and triglyceride production
To validate the LS culture as a model for drug screening, the “hepatoprotective” (UCDA) and “anti-fibrotic (Toco) drugs were tested on non-fibrotic (F0-F1), or fibrotic (F2-F3, F4) LS cultures, infected or non-infected with HCV.UCDA and Toco were dosed according to the standard of care in humans (Figure 13).On day 0, LS cultures were infected with HCVcc Con1/C3 (MOI = 0.1) and treated either with daily doses of UCDA and /or with Toco for up to day 21.

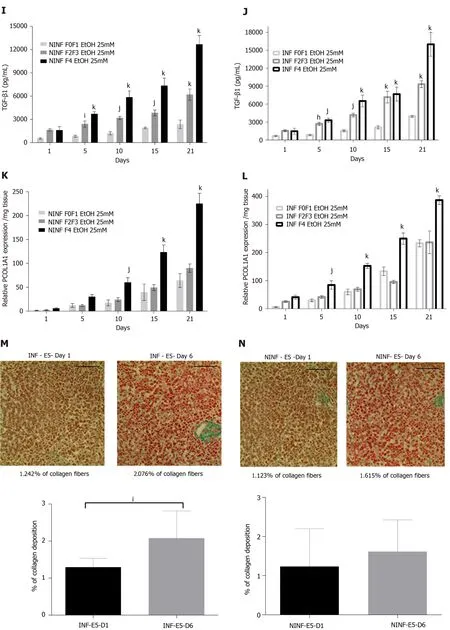
Figure 8 Significant increased expression of TGF-β1 and Procol1A1 mRNA with ethanol (1 mmol/L, 5 mmol/L, 25 mmol/L) treatment of non-infected or hepatitis C virus INF liver slice cultures from non-fibrotic (F0-F1) and ethanol (25 mmol/L) treatment of non-infected or hepatitis C virus infected liver slice cultures from fibrotic (F2-F3, F4) liver samples as shown by real-time reverse transcriptionquantitative polymerase chain reaction and ELISA and histochemistry.
During the 21-days long follow-up studies of F0-F1, F2-F3, and F4, LS cultures, a significant, 25% to 50%, reduction in TGF-β1 RNA expression was only identified in F4 LS cultures treated with Toco, from day 5 and day 10 of the culture in non-infected and HCV infected LS cultures, respectively (Figure 13A and B).Treatment with UCDA did not induce a significant reduction in TGF-β1 RNA expression in any non-infected F0-F4 LS cultures (Figure 13C).Interestingly, from day 15, at least a two-fold reduction in TGF-β1 RNA expression, at least, F2-F3, and F4 infected LS cultures was observed (Figure 13D).There was no change in TGF-β1 RNA expression in non-infected LS treated with both UCDA and Toco, whatever the stage of disease (Figure 13E).TGF-β1 RNA expression in F2-F3 and F4 infected LS cultures on days 5 and 15 was reduced by nearly two fold.On day 21, TGF-β1 RNA expression in F4 LS cultures were reduced 2.5 fold (Figure 13F).During the 21-days follow-up studies of infected and noninfected F0-F1, F2-F3, and F4 LS cultures treated with both UCDA and Toco, procollagen1A1 expression was significantly reduced in non-infected and infected F0F1- F4 LS cultures compared to untreated cultures from day 15 (Figure 13G and H).In particular, the significant reduction of procollagen1A1 RNA expression (around two-fold) in treated F2-F3 infected LS cultures was observed from day 10 and from day 15 for treated F4 infected LS cultures.Triglyceride production in HCV noninfected and infected LS from F0-F1, F2-F3 and F4 LS cultures was significantly reduced by the combination treatment from day 10 in F4 HCV non-infected LS cultures (Figure 13I) and from day 1 in F4 HCV infected LS cultures (Figure 13J).
DISCUSSION
For the first time, different stages of human liver fibrogenesis were investigatedex vivoand for a relatively long period.Indeed, liver tissue slices remained viable for at least 21 days, as shown by the secretion of albumin and urea, the percentage of ATP production and LDH release observed during the kinetic experiment.However, the secretion of albumin and urea was lower than that in micropatterned hepatocyte cocultures models[27].Both fibrotic (stages F2-F4) and non-fibrotic (stages F0-F1) liver samples remained viableex vivofor this period.Twenty one-day follow-up studies of LS cultures significantly improved the investigation of fibrogenesis in general, and fibrotic biomarkers, in particular.We obtained RT-qPCR analyses of the biomarkers (TGF-β1, procol1A1, MMP-2, MMP-9, α-SMA, HSP47, and VEGF) involved in molecular fibrogenesis, and estimation of anti-fibrotic drugs potency, in both nonfibrotic (F0-F1) and fibrotic livers samples (F2-F3, F4).Additional evaluation of fibrotic biomarkers performed by ELISA, histology, and by Western blotting supported RTqPCR data.With thisex vivomodel, sustaining hepatocyte-specific gene expression for 21 days, we induced molecular fibrogenesis using HCV, EtOH, or palmitate, thus mimicking human viral, alcoholic, and NASH liver diseases.
The most important property of this LS model is cell viability for a relatively long period of time.The expression of diverse biomarkers of fibrosis was analyzed in the presence of HCV, EtOH or palmitate using non-fibrotic F0-F1 LS cultures.The markers of fibrogenesis and triglyceride production were found to be increased in both noninfected and infected LS cultures.The addition of either EtOH or palmitate significantly increased the expression of fibrotic biomarkers.Moreover, this increase was found to be greater in HCV infected than in non-infected LS with increased triglyceride production higher in infected LS.HCV infection seemed to enhance fibrosis marker expression in the presence of ethanol or palmitate.

Figure 9 Significantly increase in TGF-β1 protein and RNA expression of α-SMA, and HSP47 in non-infected or hepatitis C virus-infected non-fibrotic (F0-F1) liver slice cultures treated with 1 mmol/L, 5 mmol/L and 25 mmol/L of ethanol was shown by enzyme-linked immunosorbent assay and real-time reverse transcription-quantitative polymerase chain reaction analyses.
It is important to mention, that TGF-β1 expression, the principal marker of fibrogenesis, was higher in non-fibrotic (F0-F1) LS cultures cultivated in the presence of HCV and /or EtOH, or palmitate treatment.This effect was greater in fibrotic (F2-F4) LS cultures.Moreover, when fibrotic LS cultures were exposed to EtOH, a significant increase of α-SMA, HSP47, procol1A1 expression as well as the other markers involved in liver fibrolysis such as (MMP-2, -9) and VEGF was identified in both non-infected and infected liver slices.
The increased expression of fibrogenesis biomarkers was throughout the twentyone days follow-up studies.RT-qPCR showed that the effect was more marked in LS cultures obtained from livers with advanced stages of fibrosis.This was confirmed for the following biomarkers: TGF-β1, α-SMA, HSP47, Procol1A1, MMP-2, -9, and VEGF.These results were further confirmed by Western blot analyses for TGF-β1, α-SMA, Col1A1, HSP47, MMP-2, -9, and VEGF.Thus, analyses of LS cultures revealed, that the progression of fibrosis is associated with an increase in the expression of certain biomarkers, in particular, α-SMA expression, and resembles a snowball effect, as shown by histochemistry results with a significant increase of collagen production in F0-F1 EtOH treated HCV infected LS on day 6 compared to day 1.As might be expected, a more marked fibrogenesis reaction was observed in fibrotic (F2-F3, F4) LS cultures, than that in non-fibrotic (F0-F1) LS cultures.

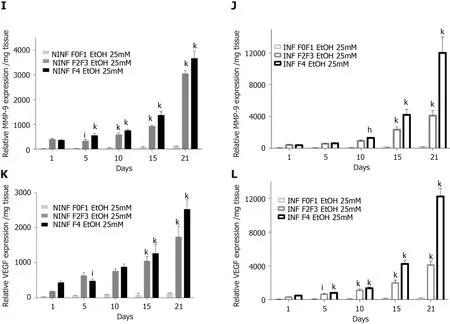
Figure 10 By real-time reverse transcription-quantitative polymerase chain reaction and enzyme-linked immunosorbent assay, significantly increase of TGF-β1 protein and RNA expression of fibrosis biomarkers HSP47, α-SMA, MMP- 2, MMP-9, VEGF increased in non-infected or hepatitis C virus-infected liver slice cultures from stages F0-F1 to stage F4 treated with 25 mmol/L of ethanol.
Thus, the LS model well responded to fibrotic inducers and, then, released a set of biomarkers that are usually detected during clinical studies in patients with fibrosis.In particular, this included TGF-β, α-SMA, Procollagen1A1, MMP-2, MMP-9, VEGF, the markers of liver fibrogenesis, whatever the origin of fibrosis.This study also showed the synergistic effect of liver comorbidities (virus, alcohol, and fat) on fibrogenesis and its consequences[28,29].Finally, the efficacy of hepatoprotective[28]or anti-fibrotic drugs[29]was suggested in the LS cultures model.Recently, Wuet al[10]demonstrated that Human liver slices collected from resected livers could be maintained inex vivoculture over a two-week period.
Several anti-fibrotic drugs are now in development[27-29], following validation in animal models[30], in particular, target inhibitors for the treatment of NASH-related fibrosis.This includes NGM282, an FGF19 analog that reduces steatosis, biliary acids injury, and lipotoxicityvia2 receptors, the MGL-3196, a THR-β1 agonist that decreases LDL-cholesterol, triglyceride and fatty liver, thus lipotoxicity[30,31].Randomized controlled trials are known to take time and the results may be disappointing despite the encouraging results of the recent REGENERATE trial, in obeticholic acid[32-36].For example,Cenicriviroc, a dual CCR2/CCR5 antagonist with positive results in mice, did not result in any significant reduction in NASH-related fibrosis after 2 years of studies[37], and Selonsertib, an ASK1 inhibitor with putative anti-fibrotic properties, was recently withdrawn in the Stellar-3 and Stellar-4 Randomized controlled trials (Gilead, Press release, April 2019).
Although randomized clinical trials are the best way to prove the drug efficacy of a drug, there are several important limitations to this approach including the need for serial liver biopsies, suboptimal dosage schedules, or placebo double-blinded controls with a single drug.All of this can require about three years.With existing LS models, anti-fibrotic drug testing can be performed for 2-3 weeks (wk).Testing is possible for single drugs, drug combinations with similar or different agents, dose effects, stability in the liver,etc…
In the present study, we used our 3D LSex vivomodels to investigate the antifibrotic properties of two drugs, being tested in clinical trials.Ursodeoxycholic acid is indicated in the treatment of primary biliary cirrhosis and dissolve radiolucent gallstones in patients with a functioning gallbladder.Alpha-Tocopherol (Toco, vitamin E) is tested currently in patients with high cholesterol and NASH.A meta-analysis in a sub-group analysis of random clinical trials has shown that alpha-tocopherol has an anti-fibrotic effect compared to UCDA[36,38].These drugs were tested alone and in a combination with the LS model.The combined treatment is not tested during the first phase of the clinical trials.The half-life of UCDA is 3.5 to 5.8 days and that of Toco is 44.5 hours.Patients must be treated daily with UCDA for 2 to 3 months and for 96 wk with Toco to obtain some clinical effects.In the LS model, Toco treatment only reduced the TGF-β1 expression in non-infected and infected LS with stage F4 after day 10.After day 15, UCDA reduced TGF-β1 expression in stage F2 to F4 infected LS.It is interesting to note that with a combination of both drugs, TGF-β1 and Procol1A1 expression was reduced significantly in LS.The level of TGF-β1 decreased nearly 2 fold in F2-F3 infected LS on day 15 and 2.5 fold on day 21, in F4 infected LS cultures.A significant reduction in procol1A1 RNA expression was found with the combination treatment in F2-F3, and in F4 infected and non-infected LS cultures with a two-fold decrease on days 15 and 21.Obviously, to confirm the results, the other dosages and proportions of drugs (in combination) should be tested.In fact, this model showed a clear decrease in the main hepatic fibrogenesis biomarker TGF-β1, in the presence of a combination of anti-fibrotic drugs (UCDA and Toco) in F2-F4 infected LS cultures with a significant decrease in both triglyceride production and Procol1A1 expression.Procol1A1 expression was significantly reduced in F2-F3, and F4 infected or noninfected LS cultures during combined treatment (UCDA and Toco).Thus, these data provide a proof of concept that this proposed 3Dex vivomodel effectively allows a rapid evaluation of new anti-fibrotic drugs.
CONCLUSION
In summary, the 3Dex vivoLS model provides hepatocyte-specific gene expression for 21 days, and effectively reproduces liver fibrogenesis related to HCV infection, EtOH, or lipids exposure, thus, mimicking human viral, alcoholic, and NASH liver diseases.Our study is the proof of concept that this relatively easy model can be used to study human liver fibrogenesis of different origins and evaluate the potency of new antifibrotic therapies that are currently under development.In particular, this system might estimate unpredictable side effects when testing certain drug combinations.
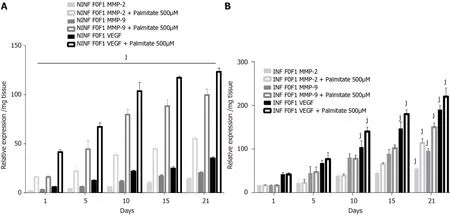
Figure 12 Significant increase of matrix metalloproteinases -2, -9, and vascular endothelial growth factor RNA expression after treatment of F0-F1 non-infected and infected liver slice cultures with palmitate (500 µmol/L).

Figure 13 During treatment with alpha-Tocopherol and ursodeoxycholic acid in combination, significant inhibition of the TGF-β1 mRNA expression of fibrotic (F2-F3, F4) hepatitis C virus INF liver slice cultures from day 5 and significant reduction of Procol1A1 mRNA expression and the triglyceride production in F0 to F4 non-infected and hepatitis C virus INF liver slice cultures during the follow-up kinetics, as evidenced the real-time reverse transcription-quantitative polymerase chain reaction analysis and enzyme-linked immunosorbent assays, respectively.
ARTICLE HIGHLIGHTS
Research background
Liver fibrosis is frequently associated with viral infection [Hepatitis C virus (HCV) and Hepatitis B virus] infection, chronic inflammation, and excessive alcohol consumption.Despite effective antiviral treatment, morbidity and hepatitis-related mortalities are still increasing.Moreover, the number of non-viral liver diseases such as nonalcoholic steatohepatitis and alcoholic liver disease is steadily growing.
Research motivation
In previous studies, we developed a three dimensional (3D) ex vivo model of HCV replication using human liver slice cultures that were followed for 10 days to evaluate a new antiviral drug.
Research objectives
We aimed to establish a 3D ex vivo liver slice model viable in vitro for 21 days allowing us to examine human liver fibrogenesis by fibrosis inducers and anti-fibrotic therapies.
Research methods
The adult human liver tissue samples from twenty patients were collected after liver resection, and divided into three groups according to their METAVIR score (F): Nonfibrotic F0-F1, obtained during surgery for colorectal cancer liver metastases or fibrotic ranging from F2 to F4.HCV infection, alcohol (ethanol stimulation), and steatosis(palmitate stimulation) were examined in non-fibrotic F0-F1 human liver slices (HLS)compared to fibrotic (F2 to F4) liver slices (FLS) infected (or not) with HCV [Con1/C3(genotype1b)] (INF).HLS of 350 μm (2.7 × 106 cells per slice) were cultivated for up to 21 days.At day 0, either ursodeoxycholic acid (only choleretic and hepatoprotective properties) and/or α-tocopherol (Toco, anti-oxidant properties which could reduce fibrosis progression) were added to standard of care concentrations on HLS and FLS.The following fibrosis markers expression were assayed in HLS, in FLS and in INF FLS, [tumor growth factor-beta (TGF-β1), Hsp47, Alpha smooth muscle actin,Procol1A1, Matrix metalloproteinases 2, 9 (MMP-2, 9), Vascular endothelial growth factor] and checked by real-time reverse transcription-quantitative polymerase chain reaction and the triglyceride production by enzyme-linked immunosorbent assay assays.
Research results
Here, for the first time, human LS cultures (stages F0-F4) were successfully maintained and evaluated for 21 days allowing to explore molecular fibrogenesis in more detail including the role of important factors such as HCV infection, ethanol (EtOH), or steatosis, three of the main causes of liver injury in clinical practice.In addition, it was demonstrated that LS cultures are efficient instruments to study anti-fibrotic drugs and their combination.We obtained real-time reverse transcription-quantitative polymerase chain reaction analyses of the biomarkers (TGF-β1, procol1A1, MMP-2,MMP-9, Alpha smooth muscle actin, HSP47, and Vascular Endothelial Growth Factor)involved in molecular fibrogenesis, and estimation of anti-fibrotic drugs potency, in both non-fibrotic (F0-F1) and fibrotic livers samples (F2-F3, F4).Expression of the fibrosis biomarkers and the progression to steatosis (estimated by triglyceride production) increased with the addition of HCV and /or EtOH or palmitate.We observed a significant decrease in both of the expression of TGF-β1, and procollagen1A1 as well as in the production of triglycerides observed in a combined antifibrotic treatment applied to the F2-F4 LS cultures infected with HCV.
Research conclusions
The 3Dex vivoLS model provides hepatocyte-specific gene expression for 21 days, and effectively reproduces liver fibrogenesis related to HCV infection, EtOH, or lipids exposure, thus, mimicking human viral, alcoholic, and nonalcoholic steatohepatitis liver diseases.Our study is the proof of concept that this relatively easy model can be used to study human liver fibrogenesis of different origins and evaluate the potency of new anti-fibrotic therapies that are currently under development.In particular, this system might estimate unpredictable side effects when testing certain drug combinations.
Research perspectives
Using theex vivomodel of human liver slice culture, the perspectives would be to evaluate the potency of new anti-fibrotic therapies alone or in combination and to study the immune components of liver disease.
ACKNOWLEDGEMENTS
The authors are deeply indebted to the patients for their essential contribution to the work.We would like to acknowledge the members of the Departments of Digestive Surgery (Groupe Hospitalier La Pitié Salpétrière and Groupe Hospitalier Cochin, Assistance Publique–Hôpitaux de Paris (AP-HP), Paris, France) and the Department of Hepatology (Groupe Hospitalier Cochin, AP-HP, Paris, France), Dr.Jérôme Guéchot (Hôpital Saint-Antoine, Pôle de Biologie Médicale et Pathologie, AP-HP, Paris, France), Dr.Phuong Nhi Bories (Groupe Hospitalier Cochin, Service de Biochimie, AP-HP, Paris, France), the staff of Genomic (GENOM’IC) and Histology (HISTIM) facilities (Cochin Institute, Paris, France) for valuable assistance.We are grateful to Dr.Matthew Albert for the critical reading of the manuscript.Dr.Daria M.Kartasheva-Ebertz received a Ph.D.fellowship from AP-HP, Paris, France.
杂志排行

World Journal of Hepatology的其它文章
- Meeting report of the editorial board meeting for World Journal of Hepatology 2021
- Extrahepatic cholangiocarcinoma: Current status of endoscopic approach and additional therapies
- Production and activity of matrix metalloproteinases during liver fibrosis progression of chronic hepatitis C patients
- Awareness of non-alcoholic steatohepatitis and treatment guidelines: What are physicians telling us?
- Occult hepatitis C virus infection in the Middle East and Eastern Mediterranean countries: A systematic review and meta-analysis
- Two-stage hepatectomy with radioembolization for bilateral colorectal liver metastases: A case report