急性高温胁迫对大口黑鲈“优鲈3号”肝脏凋亡相关酶活性和基因表达的影响
2021-04-07张佳佳周国勤王佩佩
陆 健,张佳佳,周国勤,王佩佩,庆 辉
(南京市水产科学研究所,南京 210036)
大口黑鲈(Micropterussalmoides),又名加州鲈,原产于北美洲的淡水河流和湖泊中,目前已在广东、江苏和浙江等地大规模养殖,成为中国南方地区淡水人工养殖的主养品种之一[1-3]。已有研究表明大口黑鲈的最适生长温度为20~30 ℃,且在超过36 ℃的水环境中无法生存[4]。但是近年来,由于全球气候日益恶化,夏季连续高温、极端天气频有发生,同时常伴有病害发生,这影响了鲈产业的可持续健康发展。据统计,2018年我国鲈的产量由2017年45万吨减少为42万吨[5]。大口黑鲈“优鲈3号”是2019 年经全国水产原种和良种审定委员会审定通过的新品种,具有生长速度快、饲料利用率高及适应摄食人工配合饲料等优点[6,7]。大口黑鲈“优鲈3号”作为水产领域的新星,目前已在两广和江浙地区广泛推广,将来还会迎来更加广阔的养殖前景。关于大口黑鲈“优鲈3号”的生物学特性以及抗逆性研究鲜有报道,因此有必要加强大口黑鲈“优鲈3号”的生存温度等相关生物学研究,探究其在高温胁迫下的生理变化机制,以及在高温等恶劣环境下的生存状态。
细胞凋亡(apoptosis)是一种高度调节的程序性细胞死亡[8,9]。研究认为细胞调亡包括内源途径(线粒体通路、内质网通路)和外源途径(死亡受体通路)[10-11]。但是所有通路的最终途径都是半胱氨酸蛋白酶(Caspase)的激活,所以Caspase家族在细胞凋亡中扮演着举足轻重的角色[12-15]。同时,肝脏是维持生命活动和物质代谢的重要器官,参与鱼类体内多种物质的合成、贮存、代谢、转化和分解[16]。因此,本研究以“优鲈3号”为研究对象,探究了其在极度高温胁迫下肝脏细胞凋亡相关酶活性和基因表达,旨在为 “优鲈3号”应对高温胁迫的机制和健康高效养殖提供基础数据,促进大口黑鲈养殖产业升级。
1 材料与方法
1.1 试验处理
试验所用大口黑鲈“优鲈3号”幼鱼,体质量(9.87±1.2)g,体长(7.9±0.4) cm,均来自江苏南京帅丰饲料有限公司,将试验用鱼暂养于有生物过滤水再循环系统(配备有冷却和加热功能,流速为5 L/min,光照时间为14 h)的水族箱中。每天投喂两次(8:00~9:00和20:00~21:00)江苏南京帅丰饲料有限公司大口黑鲈配合饲料。整个暂养和试验期间水中溶氧高于5 mg/L,氨氮与亚硝酸盐低于0.01 mg/L,试验前禁食24 h,试验期间不投喂。
将各组幼鱼在25 ℃水温下暂养15 d后,对照组水温保持在该温度下恒定。试验组按水温31、34、37 ℃设置,每个温度包括三个平行组,每个平行组30尾幼鱼,将暂养幼鱼迅速转入事先预调好水温的水循环缸中,并于1、3、6、12、24和48 h取6尾幼鱼的肝脏组织于液氮中速冻,然后转移至-80 ℃保存。预实验结果显示37 ℃组大口黑鲈“优鲈3号”的死亡率较高,48 h小于50%,则认为37 ℃为极限高温,测定在37 ℃下细胞凋亡相关酶活性和基因的表达。
1.2 酶活性测定
酶活性测定均采用南京建成生物工程研究所提供的试剂盒:Caspase-3(批号:G015-1-3),Caspase-9(批号:G018-1-3),准确称取50 mg肝组织,加入50 μL试剂盒中的裂解液,冰水浴条件下机械匀浆,2 500 r/min,离心10 min,取少量样品用Bradford法测蛋白浓度,之后测定Caspase-3和Caspase-9酶活性,取3次重复实验平均值。
1.3 组织RNA提取与cDNA合成
组织RNA的提取采用高纯总RNA快速抽提试剂盒(北京百泰克生物技术有限公司,RP1202),用1%琼脂糖凝胶电泳检测RNA的质量(电压120 V,电流165 mA,时间25 min),用Nanodrop 检测 RNA 的纯度(OD260/280)和浓度。根据HiScript®Reverse Transcriptase的说明(Vazyme,R123-01)进行第一链cDNA反转录。
1.4 荧光定量PCR
参考樊佳佳等[17]以18s基因为内参基因,Caspase-3和Caspase-9引物序列来自Yu等[18]参考斑马鱼(Daniorerio,NCBI:NC_007135.7)、尼罗罗非鱼(Oreochromisniloticus,NCBI:NC_031982.2)、斑点叉尾鮰(Ictaluruspunctatus,NCBI:NC_030438.1)、尖吻鲈(LatesCalcarifer,NCBI:NW_017363888.1)的BCL-2基因的 cDNA 序列,在同源保守区内用Premier 5.0软件设计基因特异性上下游引物,参考引物序列详见表1。用SYBR qPCR Master Mix(High ROX Premixed)试剂盒(Vazyme,Q341-02/03)在ABI荧光定量PCR仪扩增。反应体系:2 μL cDNA模板(5 ng/μL),4 μL的上、下游引物(10 μmol/L),10 μL 2×ChamQ SYBR qPCR Master Mix(High ROX Premixed)。扩增条件为95 ℃ 10 min; 40个循环的条件是95 ℃ 10 s,58 ℃ 60 s,熔解曲线60 ℃→95 ℃,每15 s升温0.3 ℃。每个样品重复3 次。

表1 引物信息Tab.1 The information of the primers
1.5 数据统计与分析
通过计算OD诱导剂/OD阴性对照的倍数来表示酶的相对活化程度,实时荧光定量PCR分析获得的实验结果数据采用2-ΔΔCt法计算目的基因的相对表达量。实验所得数据均用平均值±标准差的方式表示,所有数据均用 SPSS 19.0进行统计分析。利用单因素方差分析(One-way ANOVA)和LSD多重比较检验对数据处理和检验分析,P<0.05表示差异显著(* ),P<0.01表示差异极显著(**),用Origin 8.6 绘图。
2 结果与分析
2.1 极度高温胁迫对大口黑鲈“优鲈3号”肝脏凋亡相关酶活性的影响
在大口黑鲈“优鲈3号”肝脏中,相较于对照组,37 ℃高温胁迫后,Caspase3和Caspase9酶活均显著升高 (图1和图2),且随着胁迫时间的延长呈升高趋势,在48 h达到峰值,极显著高于对照组,Caspase3为对照组的5.05倍,Caspase9为对照组的4.82倍。
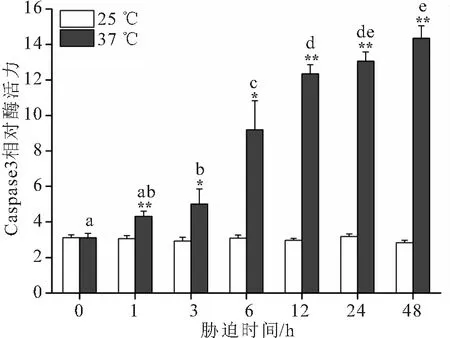
图1 高温胁迫对大口黑鲈“优鲈3号”肝脏Caspase 3活性的影响Fig.1 The effects of high-temperature stress on Caspase 3 activities in liver of M.salmoides “youlu No.3”

图2 高温胁迫对大口黑鲈“优鲈3号”肝脏Caspase 9活性的影响Fig.2 The effects of high-temperature stress on Caspase 9 activities in liver of M.salmoides “youlu No.3”
2.2 极度高温胁迫对大口黑鲈“优鲈3号”肝脏凋亡相关基因表达的影响
37 ℃高温胁迫下大口黑鲈“优鲈3号”肝脏Caspase3和Caspase9的表达量相较于对照组均呈现先升高再下降的趋势(图3和图4)。Caspase3从3 h时显著升高,在6 h的表达量达到峰值,为对照组的23.67倍,随之下降至24 h依旧显著高于对照组,48 h与对照组无显著性差异。Caspase 9的表达量相较于对照组从1 h时显著升高,在6 h的表达量达到峰值,为对照组的27.88倍,随之下降至24 h依旧显著高于对照组,48 h与对照组无显著性差异。极度高温胁迫下大口黑鲈“优鲈3号”肝脏BCL-2的表达量1 h达到峰值,显著高于对照组,之后显著下降至48 h时BCL-2的表达量仅为对照组的0.04倍 (图5)。
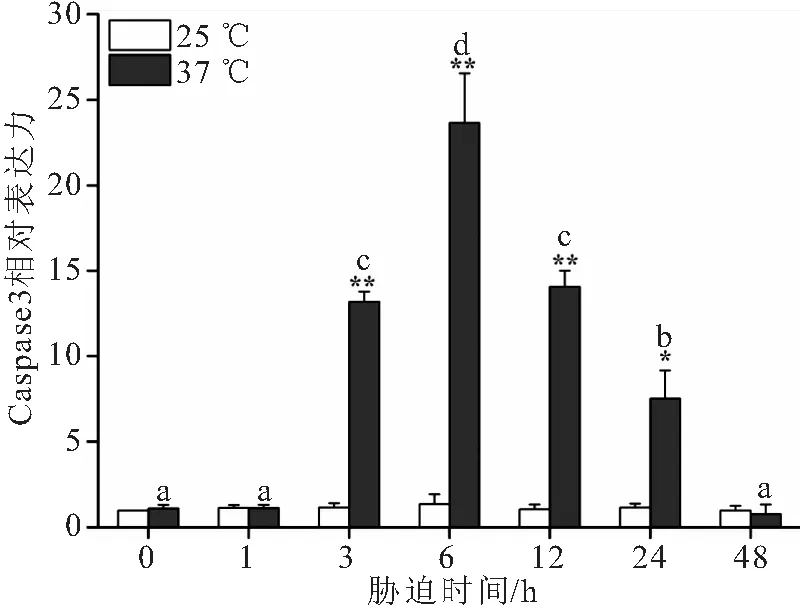
图3 温度胁迫对大口黑鲈“优鲈3号”肝脏Caspase3 mRNA 表达量的影响Fig.3 The effects of high-temperature stress on the relative expression of liver Caspase3 mRNA of M.salmoides “youlu No.3”
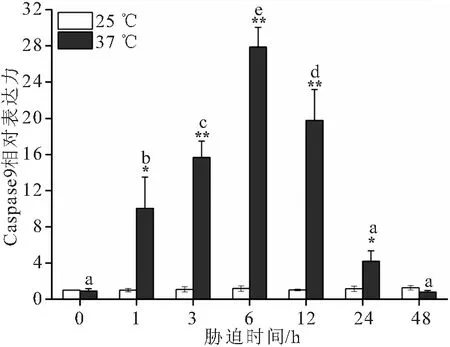
图4 温度胁迫对大口黑鲈“优鲈3号”肝脏Caspase9 mRNA 表达量的影响Fig.4 The effects of high-temperature stress on the relative expression of liver Caspase9 mRNA of M.salmoides “youlu No.3”

图5 温度胁迫对大口黑鲈“优鲈3号”肝脏BCL-2 mRNA 表达量的影响Fig.5 The effects of high-temperature stress on the relative expression of liver BCL-2 mRNA of M.salmoides “youlu No.3”
3 讨论
温度是影响鱼类生存的重要环境因子之一,主要对鱼类代谢反应起调控作用,进而能够影响鱼类的生理健康[19]。半胱氨酸蛋白酶(Caspase)家族是直接导致凋亡细胞解体的蛋白酶系统,在细胞凋亡机制中居核心地位[14]。本试验结果显示,作为凋亡的“起始者”[20],Caspase9 的表达量和酶活力在37 ℃高温胁迫后出现增加。作为凋亡的“执行者”[21],Caspase3在正常条件下是以休眠的酶原形式存在于正常细胞中,当其受到凋亡信号刺激后可以裂解特异性的蛋白质底物导致细胞凋亡,它的活化是凋亡进入不可逆阶段的标志[14,22]。本试验的结果显示,Caspase3的表达趋势与 Caspase9几乎是一致的,只是Caspase3显著变化的时间稍晚于Caspase9,表明起始子Caspases9 水解激活了处于细胞调亡关键地位的Caspase3,而使调亡开启。同样的,当斑马鱼[23]在39 ℃高温胁迫后,Caspase活性增加,相似的情况也在大西洋鲑鱼(Salmosalar)[24]、日本牙鲆(Paralichthysolivaceus)[25]、草鱼(Ctenopharyngodonidellus)[11]中发现,表明短期的急性升温应激会导致细胞的细胞膜和蛋白质受到损伤,触发激活凋亡相关系列基因的变化,从而诱导细胞发生凋亡,清除受损的细胞,来维持机体内环境的稳定[26]。但是随着胁迫时间的延长Caspase3和Caspase9均出现一定的下降,同时肝脏组织损伤较为明显,肝细胞排列无序、较多肝细胞均出现空泡化、细胞间界限杂乱模糊、细胞核萎缩,严重的甚至出现溶解,表现为细胞坏死[26],然而草鱼肾细胞在连续40高温胁迫24 h则表现出胞质空泡化、细胞凋亡率持续上升,表明细胞凋亡还在持续进行[11],造成这种情况的原因可能是连续的高温刺激致使大口黑鲈肝脏失去自我调节能力,体内平衡被打破,最终导致Caspase表达量降低,造成细胞凋亡的过程中Caspase和ATP的减少,从而发展成为不可逆转的细胞坏死,最后直至鱼体死亡[11]。值得注意的是,在高温胁迫后Caspase3和Caspase9在mRNA和酶活水平的变化并不一致,原因可能是长时间胁迫后导致总蛋白质浓度下降,从而使得基因表达下调[27]。另一方面,酶活属于蛋白质的水平,基因从mRNA 水平到蛋白质水平,需要经过转录和翻译两大程序,期间还有蛋白质的修饰等程序的启动,因此相对 mRNA 水平,基因在蛋白质水平的表达具有滞后性[28]。
BCL-2是BCL-2蛋白家族中主要的抑制调亡蛋白的代表,可以抑制细胞发生调亡提高细胞的存活率[11]。关于高温胁迫后鱼类BCL-2基因的研究报道较少,而在其他外源性刺激研究报道较多。在低氧4天(0.5 mg/L)的鲤(Cyprinuscarpio)的肝脏中检测到BCL-2的表达量增加[29],镉胁迫后草鱼肝脏BCL-2的表达量先升高再下降[30],我们的试验结果显示,BCL-2的表达量在37℃高温胁迫后1h出现峰值,表明BCL-2在抑制细胞凋亡过程中发挥作用。在胁迫6 h后BCL-2的表达量显著降低,原因可能是高温破坏了线粒体膜导致蛋白减少,细胞色素C释放到胞外激活开启了依赖于BAX的凋亡通路,使得BAX大量表达,致使BAX-BCL-2 异源二聚体被BAX-BAX同源二聚体取代,诱导细胞发生凋亡[31-32]。前期Caspase3和Caspase9基因表达量上升也可以印证这一点。同样的,高温诱导后尼罗罗非鱼性腺细胞抑凋亡基因BCL-2持续下调,且在诱导后18天仍然能检测到下调,说明抑凋亡基因可抑制细胞的凋亡[28]。另一方面,不少学者认为BCL-2可以通过发挥抗氧化作用来抑制凋亡的发生[11,12,33]。确实,我们的实验结果也表明,在高温胁迫后短时间内优鲈3号的SOD活力相较于对照组显著升高[34],这可能是机体在进行自我调节和自我保护来获得生存。当胁迫时间延长至48 h时,BCL-2的表达量仅为对照组的0.04倍,原因可能是胁迫时间较长,超过鱼体正常自我调节范围,大量的 ROS 在细胞内堆积,造成细胞氧化损伤最终导致BCL-2表达量显著降低,同时48 h时37 ℃组的存活率仅为50%也印证了这一点。因此,37 ℃可能已经成为大口黑鲈“优鲈3号”的极限温度,在日常养殖生产中,需要密切关注池塘温度的变化,减少温度应激,增加鱼体免疫力,同时做好应对高温天气的防护措施。
