短波紫外线照射对‘金都’火龙果采后保鲜的影响
2021-04-06罗恩锡李真琴洪克前宋康华梁永康王俊宁
罗恩锡,李真琴,洪克前,宋康华,梁永康,王俊宁*
(1. 广东海洋大学滨海农业学院,广东 湛江 524088;2. 中国热带农业科学院南亚热带作物研究所海南省热带园艺产品采后生理与保鲜重点实验室,广东 湛江 524091)
火龙果(Hylocereus polyrhizus)又名红龙果、仙蜜果,为仙人掌科量天尺属(Hylocereus)植物。火龙果果实风味独特、营养价值高,是集观赏、花卉、保健功能于一体的新型绿色水果[1]。火龙果原产于热带地区,成熟于高温多雨的夏秋季节,果实本身水分含量高,新陈代谢旺盛,货架期短,采后常温下贮藏极易腐烂。炭疽病、黑斑病、果腐病是引起火龙果果实腐烂的主要病害。据统计,我国火龙果每年采后损失率为20%~25%[2]。目前,生产上多使用低温、防腐剂等处理延长火龙果藏期,虽然取得一定的保鲜效果,但仍有不少问题存在,如药剂残留、冷链不健全等影响果品安全的问题[3]。
短波紫外线(ultraviotet-cirradiation,UV-C)指波长在200~280 nm的紫外线,它能穿透微生物细胞膜,引起DNA链中相邻胸腺嘧啶和胞嘧啶之间发生交联,产生嘧啶聚体,引发突变,使细胞遗传物质的活性丧失,导致微生物失去繁殖能力或死亡[4—5]。UV-C作为一种杀菌消毒方法,被广泛用于食品、医疗卫生、水处理及其他工业领域。研究发现,适宜强度的紫外线照射能延缓果蔬呼吸峰的出现和降低呼吸代谢强度[6—7],抑制果蔬采后贮藏期间病害的发生,诱导果蔬采后产生抗病性,增强抗氧化能力[7—10],增加总酚含量,减少营养物质降解,改善果蔬品质[8,11—13],从而延缓果蔬成熟衰老和减少腐烂。与其他方法相比,UV-C照射具有杀菌效果好、处理时间短、不产生化学残留物、简便易行且经济有效等优点,是一种绿色环保型的果蔬采后贮藏保鲜方法[4],目前已应用于杧果[14]、荔枝[15]、苹果[16]、桃[7,17]等多种果蔬保鲜。但因果蔬的种类和品种不同,其携带的微生物种类和自身的抗病强弱不同,UV-C处理的适宜剂量也不同。因此,筛选不同种类果蔬的合适UV-C照射剂量,是其UV-C处理过程的关键。UV-C处理虽已在多种果蔬的贮藏保鲜中应用,但在火龙果贮藏中的应用少见报道。本试验以‘金都’火龙果(H. polyrhizus‘Jindu’)为材料,采用不同剂量 UV-C照射火龙果,探究UV-C处理对火龙果采后保鲜的影响及作用机理,并筛选出合适的UV-C照射剂量,为UV-C处理技术应用于火龙果保鲜提供理论依据。
1 材料与方法
1.1 材料与处理
以‘金都’火龙果成熟果实为材料,采收后挑选大小、颜色、成熟度相同,无病虫害和机械损伤的果实试验。用 UV-C(波长为 254 nm)照射,设置0.0 kJ·m-2、1.0 kJ·m-2、2.0 kJ·m-2、3.0 kJ·m-2和4.0 kJ·m-2等5个辐射剂量处理,根据UV-C照射时间用便携式数字辐射计量仪测量剂量,每个剂量处理30个果,重复3次。处理后将果实置于22±1 ℃、相对湿度85%~90%条件下贮藏,并于处理后第0、2、4、6、8天取样测定。
1.2 方法
以果实表面出现霉斑或者水渍为依据判断火龙果腐烂,腐烂率(%)=腐烂果实数/果实总数×100%。失水率(%)=(果实采收时的重量-每次果实取样时的重量)/果实采收时的重量×100%。可溶性固形物(TSS)含量采用数显手持折光仪测定。细胞膜透性(相对电导率)测定参考张昭其等[18]的方法。SOD、POD、CAT、PPO、β-1,3-葡聚糖酶、几丁质酶活性及 H2O2含量分别采用苏州科铭生物技术有限公司的相应试剂盒进行测定。
1.3 数据处理
采用 SPSS25进行数据分析,用 LSD法和Duncan新复极差法进行多重比较和显著性检验。
2 结果与分析
2.1 UV-C剂量对火龙果采后腐烂率、电导率和TSS的影响
由图1: A所示,贮藏期间不同剂量UV-C处理的火龙果腐烂率随贮藏时间延长总体呈上升趋势,其中对照组从贮藏第 4天起腐烂率迅速上升。各UV-C处理不同程度地降低贮藏4~8 d的火龙果腐烂率,其中贮藏4~6 d内,3.0 kJ·m-2UV-C处理的腐烂率最低,其次 2.0 kJ·m-2UV-C 和 1.0 kJ·m-2UV-C处理;在第8天,除对照外,火龙果腐烂率随UV-C剂量增大而增加。

图1 UV-C处理对火龙果腐烂率、相对电导率、TSS含量的影响Fig. 1 Effect of UV-C treatment on the decay rate,relative conductivity,TSS content in dragon fruits during storage
如图1: B所示,贮藏期间各个处理火龙果果皮的相对电导率随时间延长呈上升趋势,其中1.0 kJ·m-2和2.0 kJ·m-2UV-C处理极显著降低贮藏第8 天的电导率(P<0.01),而 3.0 kJ·m-2、4.0 kJ·m-2UV-C处理分别显著提高贮藏第2天和第6天的电导率(P<0.05),其他时期各处理与对照差异不明显。各处理间,低剂量UV-C处理电导率更低。
如图1: C所示,贮藏期间各处理的火龙果TSS含量缓慢上升后下降。与对照比较,1.0 kJ·m-2、3.0 kJ·m-2UV-C处理极显著提高贮藏第4天的果实TSS含量,但降低了贮藏第6~8天的TSS含量(P<0.01);2.0 kJ·m-2、4.0 kJ·m-2UV-C 处理极显著降低贮藏后期(第4~8天)的TSS含量;贮藏结束时,低剂量处理的果实TSS含量比高剂量的要高。可见,UV-C处理降低火龙果贮藏期间的TSS含量。
综合比较不同剂量 UV-C处理的火龙果电导率、TSS和腐烂率,表明1.0 kJ·m-2UV-C能更好地降低果实腐烂率和电导率,维持相对较高的TSS含量。
2.2 1.0 kJ·m-2 UV-C处理对火龙果失水率和H2O2含量的影响
随着贮藏时间延长,火龙果失水率不断增加(图2: A)。贮藏过程中,1.0 kJ·m-2UV-C处理的果实失水率与对照变化相似,也是不断增加。1.0 kJ·m-2UV-C处理的果实在贮藏第 2天失水率极显著低于对照(P<0.01),其他时期影响不大。
在火龙果贮藏期间,对照组果实H2O2含量迅速下降,随后变化缓慢(图2: B)。不过,1.0 kJ·m-2UV-C处理的果实H2O2含量在贮藏前4天变化不大,第4天后迅速下降。显著性分析表明,1.0 kJ·m-2UV-C处理与对照比较,除第6天外(显著降低H2O2含量,P<0.05),总体上显著提高火龙果贮藏期间的H2O2含量(P<0.05),其中贮藏第2天和第4天达到极显著水平(P<0.01)。
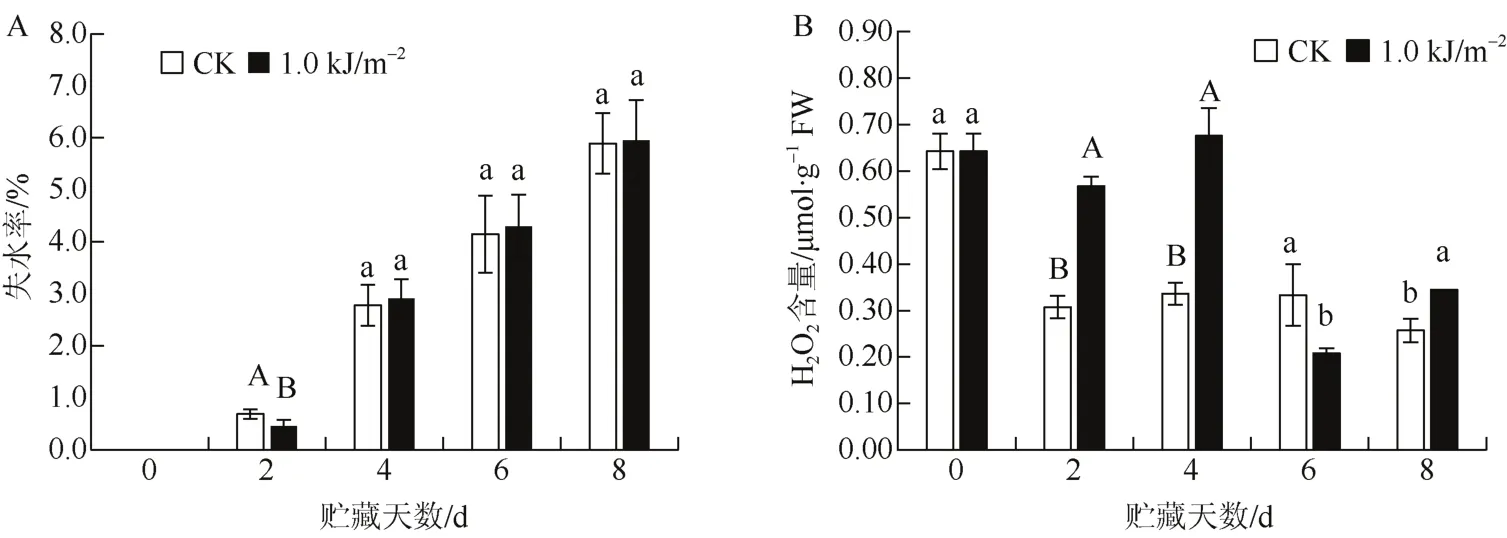
图2 UV-C处理对火龙果贮藏期失水率、H2O2含量的影响Fig. 2 Effect of UV-C treatment on water loss rate,H2O2 content in dragon fruits during storage
2.3 1.0 kJ·m-2 UV-C处理对防御酶活性的影响
如图3: A所示,对照组火龙果SOD活性在贮藏第2天略有下降,此后缓慢上升,第6天达最大后开始回落;而1.0 kJ·m-2UV-C处理的果实 SOD活性贮藏后迅速升高至最大,随后缓慢降低。与对照比较,1.0 kJ·m-2UV-C处理极显著提高贮藏期间火龙果SOD活性(P<0.01)。
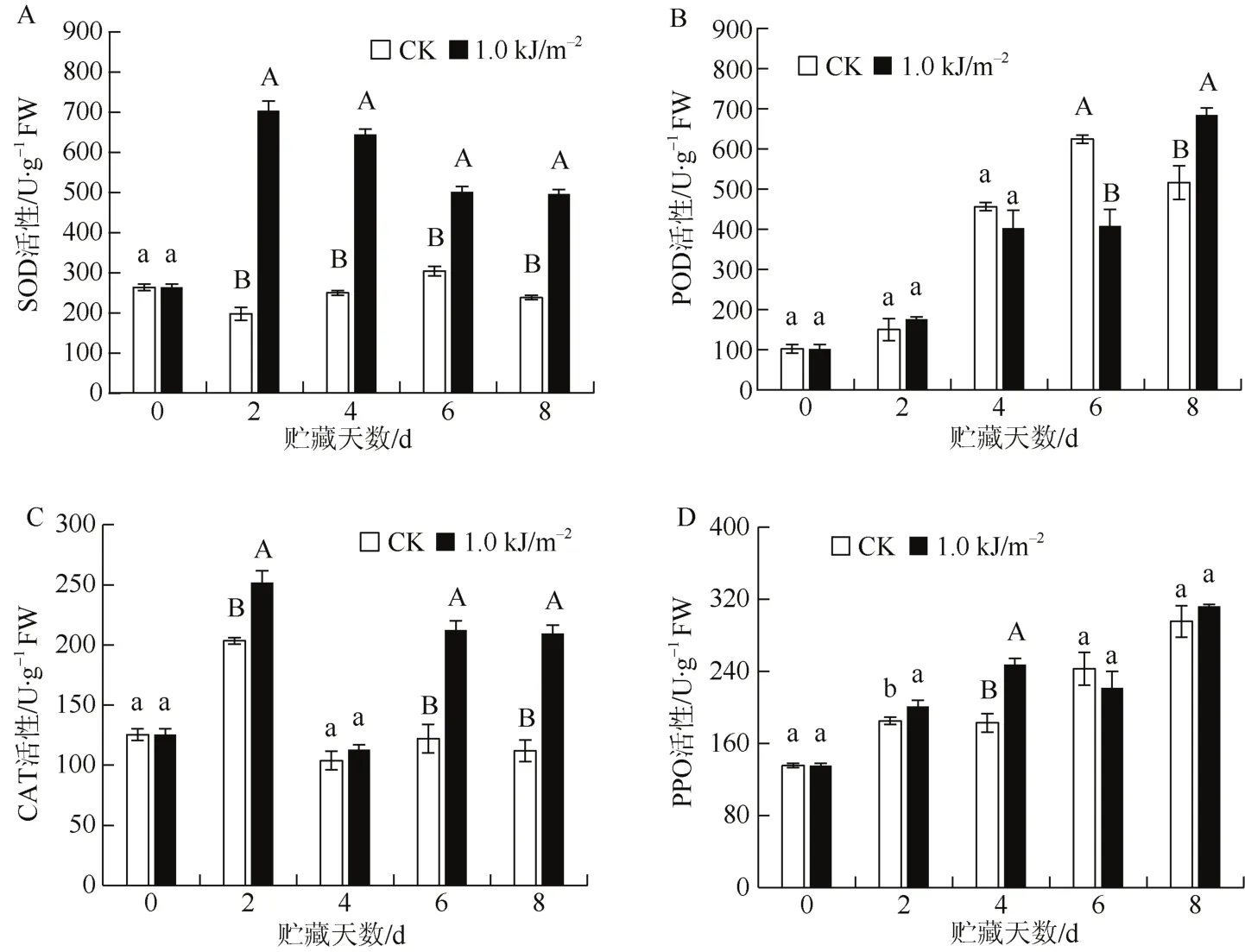
图3 UV-C处理对贮藏火龙果防御酶活性的影响Fig. 3 Effects of UV-C treatment on defensive enzymes activities in dragon fruits during storage
如图3: B所示,对照组的火龙果在贮藏前2天POD活性缓慢增加,之后快速增加,第6天达最大(693.33±11.55 U·g-1FW)后缓慢下降;1.0 kJ·m-2UV-C处理的果实POD酶活性在贮藏前4天与对照变化相似,之后变化缓慢,第 8天迅速增至最大(760.00±20.00 U·g-1FW)。1.0 kJ·m-2UV-C 处理火龙果贮藏第6天POD活性极显著低于对照(P<0.01),但贮藏第 8天的 POD活性极显著高于对照(P<0.01),其他时间与对照差异不明显。
由图3: C可见,贮藏期间火龙果CAT活性整体呈先升后降的趋势,于贮藏第 2天达到最大(203.04±2.83 U·g-1FW)。1.0 kJ·m-2UV-C 处理的火龙果 CAT活性在贮藏第 2天达最大(251.99±9.79 U·g-1FW)后,第4天有所回落,之后又快速上升后保持平稳。除贮藏第4天外,1.0 kJ·m-2UV-C处理的火龙果贮藏期间的CAT活性极显著高于对照(P<0.01)。可见,1.0 kJ·m-2UV-C处理能诱导火龙果果实抗氧化活性增强,促进果实抗衰老,起到保鲜作用。
图3: D显示,贮藏火龙果PPO活性总体呈不断上升的趋势,1.0 kJ·m-2UV-C处理的火龙果PPO活性变化趋势与对照相似,也呈整体上升趋势。1.0 kJ·m-2UV-C处理显著提高贮藏前4天的火龙果PPO活性(P<0.05),在第4天与对照差异达极显著水平(P<0.01)。可见,1.0 kJ·m-2UV-C处理能提高火龙果贮藏前期的抗病性。
2.4 1.0 kJ·m-2UV-C 处理对 β-1,3 葡聚糖酶和几丁质酶活性的影响
由图4: A看出,对照组火龙果β-1,3葡聚糖酶活性在贮藏前2天变化不大,此后开始升高,第4天达到高点后变化缓慢。火龙果经1.0 kJ·m-2UV-C处理后,其β-1,3葡聚糖酶活性缓慢上升,于贮藏第4天出现一个小的活性峰(114.82±4.83 mg·h-1·g-1FW),之后第 6天有所下降,但在第 8天迅速上升到最大(396.44±21.50 mg·h-1·g-1FW)。1.0 kJ·m-2UV-C 处理极显著降低了火龙果贮藏第6天的β-1,3葡聚糖酶活性活性(P<0.01),但提高了贮藏第8天的β-1,3葡聚糖酶活性(P<0.01),其他期间差异不明显。

图4 UV-C处理对贮藏火龙果β-1,3葡聚糖酶和几丁质酶活性的影响Fig. 4 Effects of UV-C treatment on β-1,3-glucanase and chitinase activities in dragon fruits during storage
几丁质酶可以降解病原真菌细胞壁中的几丁质,破坏细胞新物质的沉积致使病原体死亡,所产生的细胞壁碎片能诱导刺激寄主植物产生抗病反应,在植物抗病虫害的防御过程中发挥重要作用[19]。从图4: B中可以看出,贮藏期间火龙果的几丁质酶活性变化不大,但经1.0 kJ·m-2UV-C处理后,几丁质酶活性迅速增加,于贮藏第 2 天达到最大(16.67±2.11 mg·h-1·g-1FW)后开始下降,之后变化平缓。1.0 kJ·m-2UV-C处理显著提高火龙果贮藏期间的几丁质酶活性(P<0.05),在贮藏第6天达到极显著水平(P<0.01)。
3 讨论
UV-C处理可以杀灭果蔬表面的微生物,降低果蔬贮藏过程中的腐烂率。王焕宇等[10]发现,0.5~2.0 kJ·m-2剂量的UV-C处理均能抑制草莓贮藏过程中果实腐烂的发生,UV-C照射剂量越大,对果实腐烂的抑制效果越明显,并对果实可溶性固形物含量的影响不大。5.0 kJ·m-2UV-C处理能有效抑制蓝莓果实失重率和腐烂率的上升[20]。树莓[14]和杨梅[21]果实经过 UV-C处理,其腐烂率也低于对照。本研究发现,不同剂量 UV-C处理能降低贮藏期间火龙果果实的腐烂率,其中1.0 kJ·m-2的紫外线辐照效果最好,这与前人在草莓[10]、蓝莓[20]、杨梅[14]和树莓[21]上的研究结果一致。本研究还发现,UV-C处理降低火龙果TSS含量,低剂量UV-C处理降低火龙果贮藏后期的相对电导率,这与陈奕兆等[7]的研究结果不同。并且1.0 kJ·m-2UV-C处理对火龙果贮藏期间的失水率并无影响,这与UV-C在蓝莓[20]上能有效抑制果实失重率上升的结果不一致。
植物在逆境胁迫条件下,防御系统被激活,防御酶活性升高,还会诱导体内积累活性氧使植物发生抗病反应。参与植物防御的酶主要有PPO、SOD、CAT、POD等,它们在植物的抗病中,不仅单个防御酶起重要作用,而且彼此之间存在相互协调和交互作用[22]。杨华[23]等发现,0.9 kJ·m-2UV-C 处理提高了牛角椒贮藏期内果实SOD、POD和CAT活性,延缓果实采后衰老进程。在苹果上,7.5 kJ·m-2UV-C处理显著提高了果实整个贮藏期内的 PPO活性,SOD、POD活性也明显升高,提高其抗病性,有效抑制苹果青霉病和灰霉病的发生及扩展[8]。2.0 kJ·m-2UV-C处理提高草莓果实 POD、SOD 和CAT等抗氧化酶活性,促进果实贮藏前 3天 H2O2积累,但抑制贮藏后期H2O2含量上升,维持果实采后活性氧代谢平衡,延缓衰老[10]。赵静[9]研究发现,UV-C照射显著提高杧果果皮SOD活性,提升果皮内APX、POD和CAT等酶活性,降低H2O2累积。本研究发现,1.0 kJ·m-2UV-C处理极显著提高火龙果贮藏期间SOD和CAT活性(P<0.01)和贮藏第8天POD活性,这与草莓[10]、牛角椒[23]和杧果[9]上的研究结果一致;同时还显著提高火龙果贮藏前4天的PPO活性(P<0.05),这与杨梅[21]、苹果[8]的贮藏类似,但苹果经UV-C处理提高的是整个贮期PPO活性,可能不同种类的果实在抗病响应中各防御酶所起作用的阶段不同。
H2O2是活性氧(ROS)的重要成分,是植物-病原微生物互作系统中产生的主要 ROS形式。H2O2在植物应对逆境产生抗病防御反应中发挥重要作用[24]。本研究发现,1.0 kJ·m-2UV-C处理总体上显著提高火龙果贮藏期的 H2O2含量(贮藏第 6天除外),诱导果实产生抗病性,这与草莓的结果一致[10],但不同于杧果[9],说明UV-C处理对不同种类的果实H2O2含量影响不同。在贮藏第6天,1.0 kJ·m-2UV-C处理火龙果的POD活性和H2O2含量均极显著低于对照(P<0.01),这可能与该时期高的CAT活性足以清除体内积累H2O2,不需提高POD活性就能维持H2O2代谢平衡有关。
β-1,3-葡聚糖酶和几丁质酶是植物受到外界逆境刺激时产生的2种病程相关蛋白,二者在抑制真菌生长过程中具有协同作用[19]。王焕宇等[10]发现,2.0 kJ·m-2UV-C处理显著抑制草莓果实贮藏期间灰霉病病斑的扩展,增强果实PAL、PPO、β-1,3葡聚糖酶和几丁质酶活性,同时诱导总酚大量积累,认为 UV-C处理能通过诱导草莓果实增强抗病性从而抑制其采后腐烂。在杨梅上,3.0 kJ·m-2UV-C处理通过提高PAL、POD、PPO、几丁质酶和β-1,3-葡聚糖酸酶等抗病相关酶活性来增强果实抗病性,防止桔青霉病原菌侵染,从而有效地抑制果实腐烂[20]。本研究发现,1.0 kJ·m-2UV-C处理显著提高火龙果贮藏前 6天的几丁质酶活性和贮藏第 8天的 β-1,3葡聚糖酶活性,说明1.0 kJ·m-2剂量UV-C处理能够通过诱导增强几丁质酶和β-1,3-葡聚糖酶的活性,在一定程度下提高火龙果果实的抗病能力,减少果实腐烂,这与王焕宇等[10]、喻譞等[21]的研究结果一致。另外,1.0 kJ·m-2UV-C处理火龙果的几丁质酶和β-1,3-葡聚糖酶活性在不同时期出现增强,这可能与二者在火龙果果实的采后抗病过程中具有互补、协同抗病的作用有关。
