油茶花芽分化形态结构及内源激素变化
2021-04-06李孟南戴秋月黄永芳杨运源
李孟南,戴秋月,黄永芳,杨运源,邓 征
(华南农业大学林学与风景园林学院,广东 广州 510642)
油茶(Camellia oleifera)是山茶科(Theaceae)山茶属(Camellia)植物,其特点是果实油脂含量较高,具有栽培经济价值,是我国特有的木本油料树种[1]。内源性植物激素是由植物自身代谢产生的一类具有高度生物活性的有机小分子化合物,激素信号往往通过调节基因表达水平影响成花[2]。研究表明,植物内源激素水平与花芽分化、成花有着十分紧密的联系[3]。其中,激素间存在协同或颉颃作用,因此激素的动态平衡对花芽分化有促进作用,有利于植物成花[3—6]。
目前,关于油茶花芽发育过程中内源激素与花芽分化的关系研究较少[7]。本研究测定 4个油茶无性系花芽分化期内源激素的动态变化,探讨内源激素与花芽分化的关系,为提高油茶花朵数、坐果率及油茶产量而采取田间管理措施提供理论基础。
1 材料与方法
1.1 材料
试材取自华南农业大学增城教学科研基地种植的5年生4个普通油茶无性系:岑软2号(C. oleifera‘Cenruan 2’)、岑软 3 号(C. oleifera‘Cenruan 3’)、湘林 8 号(C. oleifera‘Xianglin 8’)、粤华 5 号(C. oleifera‘Yuehua 5’,原为华南农业大学选育的阳春5号)。
1.2 方法
1.2.1 石蜡切片法观察花芽结构
选取树冠中上部外围不同方向、生长良好的花芽10~20个。4月至5月,每10 d取样一次;6月至10月,每5 d取样一次。花芽在FAA溶液固定24 h后转入70%乙醇,放入4 ℃冰箱保存备用。
参照《植物显微技术》石蜡切片法制片[8],0.5%苏木精染色,2%铁矾分色,中性树胶封片后,置于显微镜下观察并拍照。
1.2.2 酶联免疫吸附分析法测定内源激素
每品种选择3株长势一致、生长健壮、无病虫害的植株,每株选取不同方向、生长良好、中等长度的当年生枝条上的花芽5~10个。从5月10日开始,每隔20 d采样,直至10月20日。花芽取样后打包、标记迅速放入液氮中保存,并放入–80 ℃超低温冰箱保存待测。
根据《植物激素及其免疫检测技术》的方法,借助试剂盒通过酶联免疫吸附分析法测定材料中玉米素核苷(ZR)、脱落酸(ABA)、生长素(IAA)、赤霉素(GA)含量[9]。
1.2.3 数据处理
采用 Excel统计分析软件分析数据,Origin和Adobe Photoshop软件作图。
还可以使用创可贴疗法:对病斑纵划道,将蘸有EM菌剂,或枯草芽孢杆菌,或50倍液50%氯溴异氰尿酸的药棉或卫生纸敷于病斑及划口处,然后用薄膜包裹。
2 结果与分析
2.1 油茶4个无性系花芽分化时期及结构特征
对油茶4个无性系花芽发育的时间进程观察见表1,花芽在5~7月长度增加较快,表明花器正在形成。普通油茶花芽具鳞片5枚,先端浅裂;花萼3,复瓦状排列,具有金黄色绢毛;花瓣5,分离;雌蕊花柱先端3裂,少有4裂,子房上位,有毛,3~5室,每室有2~3颗胚珠;雄蕊多数,无毛,花丝分离,3~4轮。

表1 油茶4个无性系花芽分化各阶段的时间(月-日)Table 1 The stages of flower bud differentiation in 4 Camellia oleifera clones
2.1.1 前分化期
分前期和后期。前期花芽的生长点稍尖,外部形态与叶芽无明显区别。前分化期从4月上旬至 4月中旬,花芽分化约10 d (图1: A)。
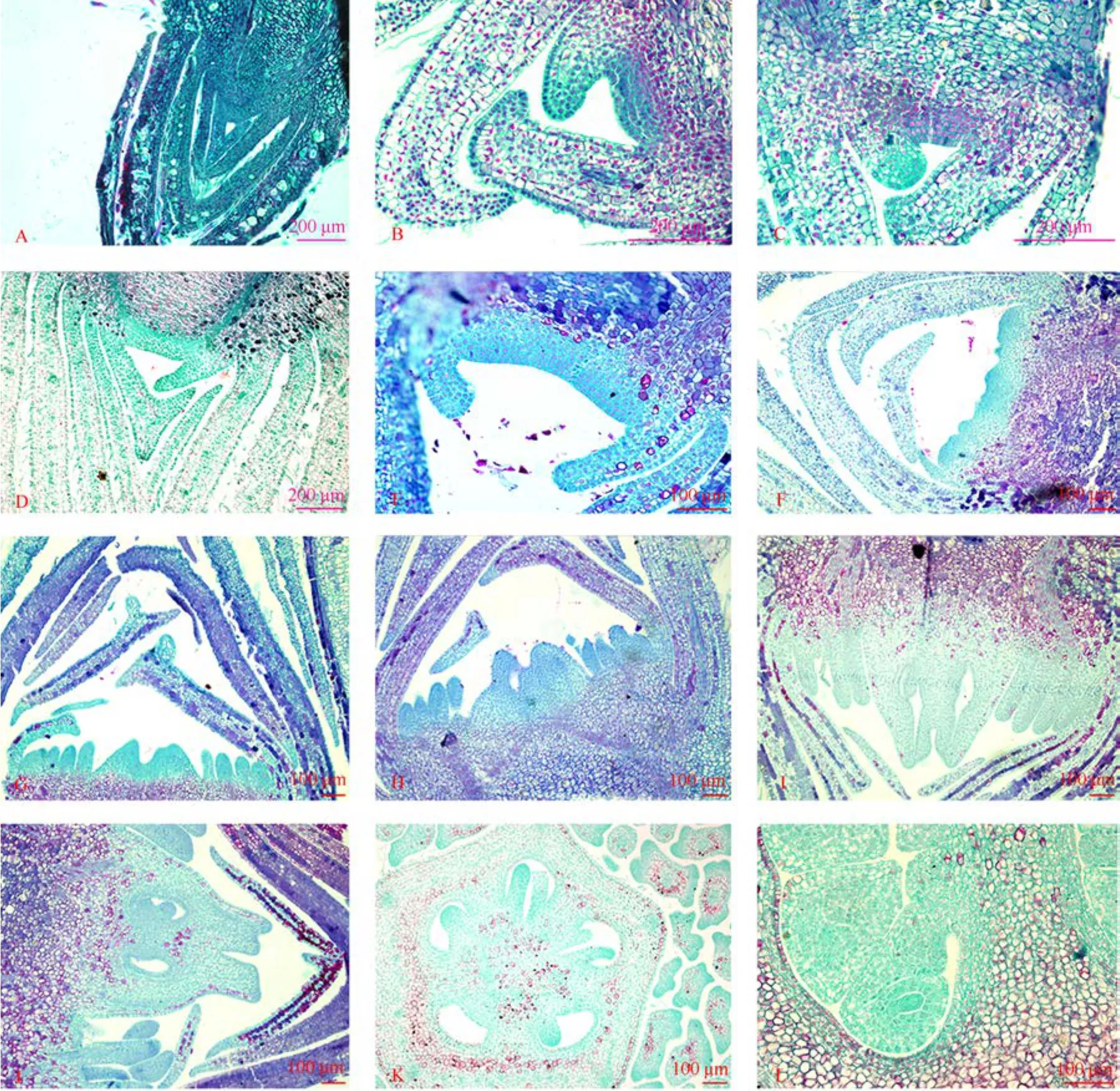
图1 油茶花芽分期形态变化Fig. 1 Morphological change of Camellia oleifera flower buds
2.1.2 萼片形成期
分前期与后期。前期萼片原基开始在生长点两侧出现,然后花萼原基向内延伸并弯曲;在萼片形成后期,生长点变扁平,而萼片继续伸长直至完全覆盖生长点,与此同时,花瓣原基出现在生长点上(图1: B~C)。这一阶段发生在4月中旬至5月中旬,持续约20 d 。
2.1.3 花瓣形成期
2.1.4 雌雄蕊形成期
分前期和后期。在花瓣形成后期,生长点变得更宽,略有凹陷,雌雄蕊原基同时出现在生长点上,雌蕊原基是中央3~5个较大的突起,雄蕊原基是周围一些小而互生的突起(图1: F~G)。在雌雄蕊形成后期,能明显地看到多层雄蕊围绕着中央雌蕊。通常雌雄蕊形成于6月中旬至6月下旬。
2.1.5 子房与花药形成期
在雌雄蕊形成后期,雌蕊下部膨大形成“V”型子房,雌蕊上部伸长,并靠拢形成柱头(图1: H)。此时花药开始在雄蕊中形成。子房与花药分化时间为6月下旬至7月上旬,历经10 d。
2.1.6 雌雄蕊成熟期
此时柱头继续伸长,子房膨大成囊状,3室,每室 2~3个胚珠。此时花药已全部形成(图1:I~L)。这个过程从 7月上旬到 7月下旬,历时约 20 d。
整个花芽期从4月上旬至7月,历时3~4月,花蕾期从7月至10月中旬,历时3个多月。油茶4个无性系花芽分化时间不一致。湘林8号分化快,粤华5号分化时间相对滞后,岑软2号、3号比湘林8号滞后10 d左右。
2.2 油茶4个无性系花芽激素含量动态变化
2.2.1 ZR含量
油茶花芽中 ZR相对含量较低,不同无性系花芽的ZR含量略有差异,各无性系的花芽ZR含量也随着时间变化而变化(图2: A)。湘林 8号和粤华 5号的ZR含量变化趋势一致,呈“升–降–升–降”变化趋势,在花芽期ZR含量先升后降,在现蕾期ZR含量略微上升后持续下降。湘林8号ZR含量最大值13.308 ng·g–1FW,出现在现蕾期中期,粤华5号最大值为14.916 ng·g–1FW,出现在现蕾期中期。岑软3号和岑软2号花芽ZR含量变化相似,基本呈“升–降–升–降–升” 趋势,总体含量呈上升趋势,在花芽期ZR含量先升后降,ZR含量在现蕾期中期先升后降,在现蕾期后期显著上升。其中岑软2号的ZR含量变化波动较大。岑软 3号最大值13.928 ng·g–1FW,出现在现蕾期中期。4个无性系的ZR含量无显著差异。

图2 油茶4个无性系花芽内源激素含量动态变化Fig. 2 The changes of endogenous hormones content in flower buds of 4 clones of Camellia oleifera
2.2.2 ABA含量
随着时间的变化,湘林8号和粤华5号花芽中ABA 含量出现“降–升–降–升”的变化趋势,总体上逐渐降低;岑软3号和岑软2号表现出“升–降–升”的变化趋势,在花芽分化前期,ABA含量升高迅速(图2: B)。4个无性系油茶的最高ABA含量均出现在萼片形成期,随后ABA含量均显著降低,并保持在较低水平,在花蕾期 ABA含量有所回升。ABA 含量的变化范围在 76.815 ng·g–1FW~137.648 ng·g–1FW。
2.2.3 IAA含量
油茶无性系花芽分化期与现蕾期IAA含量均呈先缓慢升高而后下降再回升的变化趋势,总体来看,在花芽分化期IAA含量是不断增加的,在现蕾中期IAA含量降低,至现蕾后期 IAA含量略有上升(图2: C)。其中湘林8号和粤华5号在花瓣形成期IAA含量下降,岑软2号和岑软3号在花瓣形成期IAA含量升高,直至现蕾中期IAA含量达最高值,临近开花时,IAA含量降低。IAA含量的变化范围在在 49.072~13ng·g–1FW。
2.2.4 GA含量
油茶花芽中GA含量相对较低,随花芽发育不断降低。花芽分化前期GA含量相对较高,在现蕾中期,GA含量降低一半(图2: D)。说明GA有抑制花芽分化的作用。湘林8号GA含量降低幅度最大,由花芽分化前期的最高值13.720 ng·g–1FW降至现蕾后期的最低值4.416 ng·g–1FW。粤华5号与岑软3号、岑软2号降幅近似。
3 结论与讨论
油茶花芽分化过程分为前分化期(10 d)、萼片形成期(20 d)、花瓣形成期(30 d)、雌雄蕊形成期(20 d)、子房与花药形成期(10 d)和雌雄蕊成熟期(20 d)等6个阶段[10—12],历时3~4个月。花芽在春梢褐化20 d左右开始分化,雌蕊成熟时间长,从7月上旬直至7月下旬,此时花器官原基雏形已形成,花蕾外部形态肉眼可辨。现蕾期从7月至10月中旬,此期间花丝伸长、药隔出现、柱头分裂、子房发育,10月中旬开始开花。油茶4个无性系的花芽分化时间不同,湘林8号分化时间早,分化快;粤华5号与湘林 8号分化起始时间相近,但雌雄蕊分化比湘林 8号慢5 d。岑软3号和岑软2号分化时间一致,均比湘林8号时间推迟15 d。
ZR作为一种细胞分裂素,具有促进细胞分裂、花芽分化和促进植物生长的作用,能够提高作物产量,增强植物抗逆性[13—14]。ZR含量一定程度上反映植物体内细胞分裂及代谢活动的强度。油茶花芽分化期间,花原基细胞迅速分裂成花萼、花瓣、雄蕊、雌蕊,此时 ZR含量不断增加,有利于花器官形成。至现蕾前期,雌蕊进入成熟期,ZR含量迅速下降,但随着胚珠的形成,ZR含量继续增加并保持在较高水平。油茶花芽分化中 ZR含量波动较大,与金花茶(Camellia petelotii)叶片[15]、油桐(Vernicia fordii)叶片[16]中ZR含量变化相似。其中,粤华5号和湘林8号的ZR含量变化趋势相近,岑软3号和岑软2号ZR含量变化趋势一致。这与不同油茶无性系之间花芽分化时间的早晚密切相关。
ABA对于成花的作用尚有争议,可能与植物种类有关。在苹果(Malus pumila)、梨(Pyrus)、李(Prunus salicina)中 ABA 对成花没有直接影响[17],但 ABA可明显抑制营养生长、诱导休眠,从而间接影响花芽孕育。油茶4个无性系的花芽ABA含量相对较高,在油茶花芽分化过程中ABA含量逐渐降低。花芽前分化期及萼片形成期,ABA含量升高迅速。油茶4个无性系的ABA含量均在萼片形成期达最高,并在花瓣形成期和雌雄蕊形成期出现不同程度的下降,而后保持在较稳定的水平。在花芽分化期和现蕾期,ABA与IAA存在明显的颉颃作用,即ABA含量上升,IAA含量下降。韦靖杰等[18]研究也表明,油茶现蕾期至谢花期花蕾、叶、枝条各器官中的IAA 和ABA均存在颉颃作用。
对不同植物的研究表明,低浓度IAA促进苹果[6]、刺梨(Rosa roxburghii)[19]花芽分化,而高浓度IAA对银杏(Ginkgo biloba)雌花芽分化有促进作用[20]。油茶 IAA 绝对含量高(49.072~135.622 ng·g–1FW),在花芽发育进程中均呈现出先缓慢升高后下降又回升的波动变化,整个花芽分化过程中IAA含量不断增加,临近开花时IAA含量降低,说明高浓度IAA促进油茶花蕾形成,低浓度IAA有利于开花。这与韦靖杰等[18]的研究相似,而喻雄等[21]研究表明,油茶‘铁城一号’IAA含量较低,这可能是品种不同或油茶大小年导致的。
一般植物花芽分化的先决条件是GA 含量下降[22]。与其他三种内源激素含量相比,GA含量相对较低(5.616~13.720 ng·g–1FW),随着花芽发育不断减少。在整个成花过程中缓慢下降,这说明GA抑制花芽分化,这与油茶‘铁城一号’研究结果一致[21]。
植物花芽分化受内源激素影响,同时各种内源激素相互颉颃、协调从而达到动态平衡,进而调控植物花芽分化[23]。油茶品种不同,其内源激素含量及变化也有差异。龙眼(Dimocarpus longan)内源激素在大小年变化不同[4],油茶大小年严重,其内源激素变化在大小年可能表现不同。此外,油茶成花及内源激素还与降水、气温、营养条件有较大关系[24]。研究表明,改变温度、控制土壤含水量、施肥、喷施外源激素、修剪等对花芽分化具有调控作用[25—28]。ZR促进细胞分化,有利于花器官的形成;IAA促进花芽分化;IAA与ABA之间存在颉颃作用。因此,在生产上可通过设置不同配比的ZR、IAA、ABA处理调节花芽分化期从而改变开花时间,降低落花落果率,提高结实率。
