A New Species of the Gekko japonicus Group (Squamata:Gekkonidae) from Southwest China
2021-04-02YinmengHOUShengchaoSHIGangWANGGuochengSHUPuyangZHENGYinQIGaohuiLIUJianpingJIANGandFengXIE
Yinmeng HOU ,Shengchao SHI ,Gang WANG ,Guocheng SHU ,Puyang ZHENG ,Yin QI ,Gaohui LIU,Jianping JIANG and Feng XIE*
1 CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization and Ecological Restoration Biodiversity Conservation Key Laboratory of Sichuan Province,Chengdu Institute of Biology,Chinese Academy of Sciences,Chengdu 610041,Sichuan,China
2 Key Laboratory of Bio-Resource and Eco-Environment of the Ministry of Education,College of Life Sciences,Sichuan University,Chengdu 610065,Sichuan,China
3 College of Chemistry and Life Science,Chengdu Normal University,Chengdu 611130,Sichuan,China
4 College of Chemistry and Life Science,Institute of Ecological Environment Protection,Chengdu 611130,Sichuan,China
5 Yibin University,Yibin 644000,Sichuan,China
6 University of Chinese Academy of Sciences,Beijing 100049,China
7 Chinese Research Academy of Environmental Sciences,Beijing 100012,China
Abstract A new species of the Gekko(Squamata:Gekkonidae)is described from the border of Sichuan and Yunnan Province,southwest China,based on distinct morphological and molecular features.Gekko jinjiangensis sp.nov.is distinguished from congeners by a combination of the following characters:size small (SVL 50.2-61.6 mm,n=13);nares in contact with rostral;interorbital scales between anterior corners of the eyes 20-24;ventral scales between mental and cloacal slit 146-169;midbody scale rows 111-149;ventral scale rows 31-47;subdigital lamellae on first toe 8-11,on fourth toe 11-15;no webbing in the fingers and toes;with tubercles on uapper surface of fore and hind limbs;precloacal pores 4-5 in males;postcloacal unilateral tubercles 1-2;dorsal surface of body with 8-9 large greyish brown markings between nape and sacrum.In molecular analyses,the new species is sister to G.scabridus,but separated from them by approximately 9.9%-12.2% in genetic divergence as shown by a fragment of the partial mitochondrial ND2 gene.The new species is the highest Gekko with elevation range from 2000 to 2476 m.Further surveys are recommended to better understand the occurrence and population status of the new species.
Keywords Gekko jinjiangensis sp.nov.,Gekko japonicusgroup,molecular phylogenetic analysis,morphology,Jinsha River
1.Introduction
The genusGekko
Laurenti,1768 belongs to Gekkonidae Gray,1825 of Sauria (Lacertilia),distributed across tropical Asia and western Oceania,from India eastwards to Melanesia.The genus currently contains 77 species,of which 19 species are known from China (Uetz,2020).Woodet al
.(2020) presented a new classification emphasizing the most inclusive,original generic name(Gekko
) for these~60 taxa,arranged into seven subgenera:SubgenusGekko,Japonigekko,Ptychozoon,Rhacogekko,Lomatodactylus,Balawangekko,Archipelagekko
.From the phylogenetic relationship,the subgenusJaponigekko
is equivalent in content to theGekko japonicus
Group (Woodet al
.,2020).G.japonicus
group is the most diverse group in the genusGekko
,which have a wide distribution in eastern Asia,from China eastwards to Japan,southwards to Vietnam and Laos(Rösleret al
.,2011;Luuet al.
,2015).The members of this group are characterized by a moderate size;nare usually in contact with rostral;two or three nasals;0-21 rows of dorsal tubercles;0-32 precloacal pores;1-4 postcloacal tubercles;the webbing between fingers and toes weakly to extensively developed;lateral folds without tuber.TheGekko japonicus
groupcurrently contains 27 described species (Luuet al
.,2017;Woodet al
.,2020).The Jinsha River basin locates in the Mountains of Southwest China,which is one of biodiversity hotspots and one of the most concentrated and vulnerable areas of the forest ecosystem along the Yangtze River.During the field surveys in 2019 in the Jinsha River region,near the border of Sichuan and Yunnan Province (Prov.),China,a series of gekkonid specimens were collected in the middle reaches of the Jinsha River.Morphologically,these specimens can be assigned to theGekko japonicus
group based on the following features:size small;nare in contact with rostral;postcloacal tubercles present;no webbing between fingers and toes;lateral folds without tubercles;subcaudals enlarged;dorsum with bands (Rösleret al
.,2011;Nguyenet al
.,2013).Molecular data showed that the specimens from Sichuan and Yunnan Province were clustered in the same clade withGekko scabridus
,the specimens are monophyly and compose the sister species toG.scabridus
.And,the molecular divergence calculated using data from a fragment of the mitochondrial NADH dehydrogenase subunit 2 (ND2) gene between the two species is approximately 9.9%-12.2%,it is much higher than the threshold interspecific separation ofG.japonicus
group inGekko
species from ND2,which is approximately 6.6% (Luuet al
.,2015,2017).From the morphological comparisons and molecular phylogenetic analyses,we indicate that the specimens should represent an undescribed species.2.Materials and Methods
2.1.Sample preparation
Thirteen adult individuals (9 females,4 males) of the undescribed species were collected in August and September,2019 in the Jinsha River region,between the border of Sichuan and Yunnan Prov.,China (Table 1).For comparisons,some specimens of congeneric species were also collected,G.scabridus
from Panzhihua (Yanbian),Sichuan Prov.(The collection site is located between the two type locations Yongren and Miyi).After taking photographs,the individuals were euthanized using isoflurane,and the specimens were then fixed with 10% formalin and preserved in 75% ethanol.Liver tissue samples were taken and preserved separately in 95%ethanol prior to fixation.Specimens were deposited in Chengdu Institute of Biology,Chinese Academy of Sciences (CIB,CAS).2.2.Molecular data and phylogenetic analyses
Genomic DNA from each specimen was extracted using a TIANamp Genomic DNA Kit by TIANGEN (BEIJING) BIOTECH,China.The fragments of NADH dehydrogenase subunit 2 (ND2) gene was amplified using the primer pairs L4437b (Maceyet al
.,1997)and ND2r102 (Greenbaumet al
.,2007).PCR amplifications were performed in a reaction volume of 25 ul.Fragments were amplified under the following conditions:for ND2,an initial denaturing step at 95 °C for 2 min;32 cycles of denaturing at 95°C for 35 s,annealing at 50 °C for 35s and extending at 72 °C for 95 s.Sequencing was conducted using an ABI3730 automated DNA sequencer in Shanghai DNA BioTechnologies Co.,Ltd.(Shanghai,China).New sequences were deposited in GenBank(for GenBank accession numbers see Table 1).For molecular analyses,the available sequence data for all related species of the genus were downloaded from GenBank.For phylogenetic analyses,the available sequence data ofLepidodactylus moestus
(Peters,1867) was downloaded,and used as outgroup (All GenBank accession number see Table 1).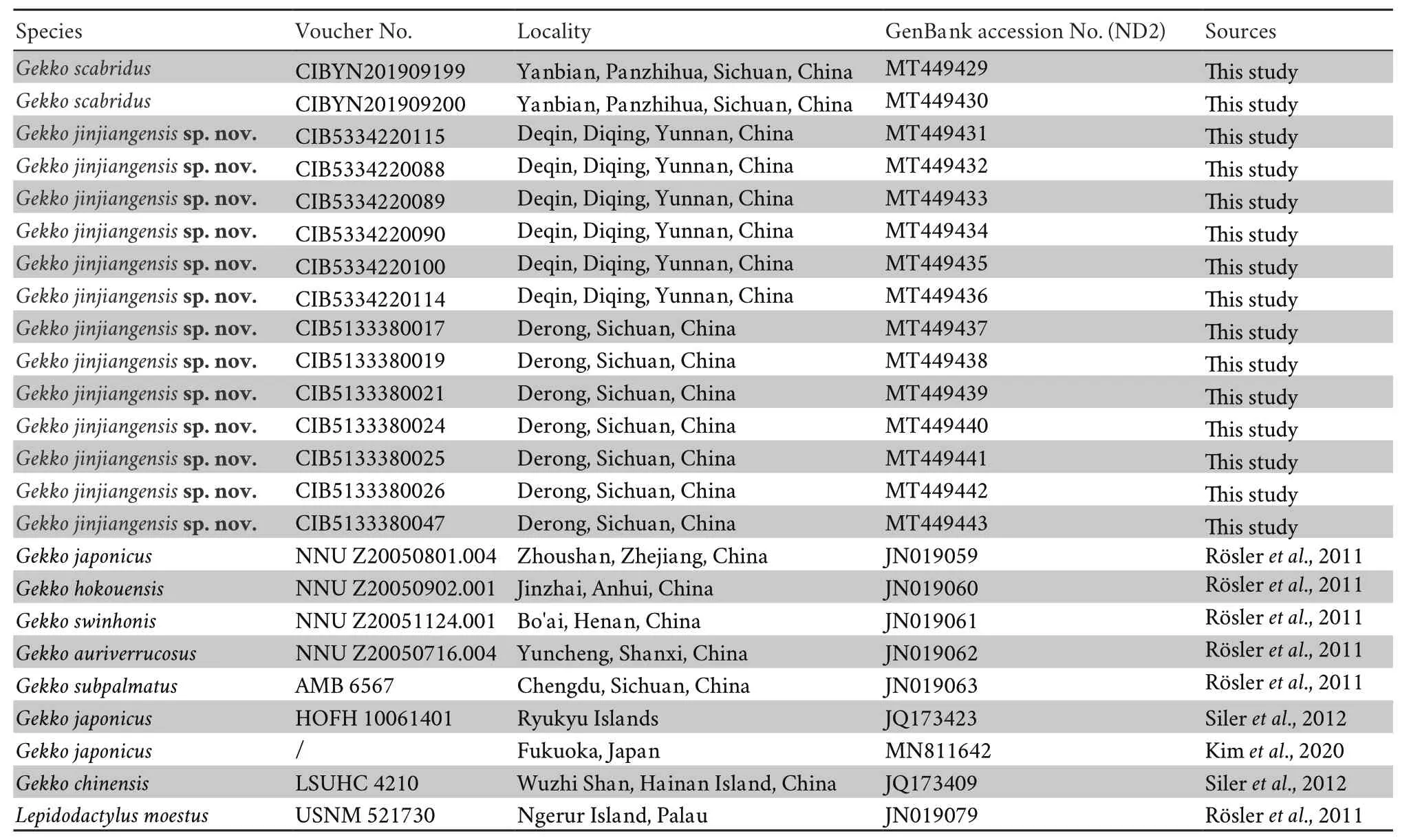
Table 1 Samples used in the molecular analyses,including their GenBank accession No.,Voucher No.and locality.”/”means no available data.
Sequences were assembled and aligned using the Clustalw options in BioEdit v.7.0.9.0 (Hall,1999) with default settings.Alignments were checked by eye and modified manually if necessary.Phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian Inference (BI)methods,implemented in PHYML Online execution (Guindonet al
.,2010) and MrBayes 3.2.2 (Ronquistet al
.,2012),respectively.The maximum likelihood (ML) analyses in PhyML software,with NNI search,automatic model selection by SMS (Smart Model Selection),under a GTR+G+I substitution model.The reliability of the phylogenies was estimated with the approximate likelihood-ratio (aLRT) SH-like test (Anisimova and Gascuel,2006).In BI analyses,two runs each with four Markov chains were run for 20 million iterations with sampling every 1000 generations.The first 25% of generations were removed as the“burn-in”stage followed by calculation of Bayesian posterior probabilities and the 50% majority-rule consensus of the post burn-in trees sampled at stationarity.Trees were visualized with FigTree v1.4.2 program (Rambaut,2012).Genetic distance was calculated with the pairwise uncorrectedp
-distance model betweenGekko
species on ND2 gene using MEGA v.7 (Kumaret al
.,2016).2.3.Morphological comparisons
A total of 13 adult specimens (9 females and 4 males) representing the type series of undescribed species (for voucher information see Table 1).The terminology and methods followed Rösleret al.
(Rösleret al
.,2011).Sex was determined by presence of precloacal pores.Morphological data of theG.scabridus
were obtained from specimens near type locality (see Table 1) and literatures (Zhouet al
.,1982;Zhaoet al
.,1999).For other species,data cited from literatures (Stejneger,1901;Zhouet al.
,1982;Song,1985;Zhaoet al
.,1999;Goris and Maeda,2004;Rösleret al
.,2004;Todaet al
.,2008;Zhou and Wang,2008;Rösleret al
.,2010;Phung and Ziegler,2011;Nguyenet al
.,2013;Luuet al
.,2014;Ngoet al
.,2015;Yang,2015;Luuet al
.,2015,2017).2.4.Morphological characters
Measurements were taken with digital calipers to the nearest 0.1 mm.In total,32 morphometric characters of adult specimen were measured.The following abbreviations were used (Table 2):3.Results
3.1.Phylogenetic analyses
Aligned sequences matrix of ND2 contained 552 bp.ML and BI trees presented similar topology(Figure 1).TheG.japonicus
group was strongly supported asa monophyletic group.All 13 specimens of the new species forms an independent clade,and nested in theG.japonicus
group,sister toG.scabridus.
Uncorrected pairwise divergences within these specimens range from 0% to 3.6% (Table 3).The uncorrectedp
-distance between the undescribed species and other known species from theG.japonicus
group range from 9.9% (G.scabridus
) to 27.0% (G.auriverrucosus
andG.chinensis
),which is among inter-species level (Luuet al
.,2015,2017).Herein,we describe these specimens as a new species.
Table 2 Morphological characters and abbreviations used in this study.Bilateral scale counts were given as left/right.
3.2.Morphological analyses
Taxonomic accounts
sp.nov.HOU,SHI,WANG,JIANG and XIE
Gekko japonicus
Yang and Rao,2008.Amphibian and reptilia of Yunnan.177-181.Holotype
CIB5334220115 (Figure 2),adult male,from BaiSheng Village,Benzilan Town,(28.23463° N,99.30227° E,2045 m a.s.l.),Deqin County,Yunnan Prov.,China,collected by Shengchao SHI,Jianyi FENG,Lei FU on 25 August 2019.Paratype
Twelve adults (9 females and 3 males),collected from the two places.Specimen CIB5334220088-90,CIB5334220100,CIB5334220114 were collected from Benzilan Town (Diqing Tibetan Autonomous Prefecture),Yunnan Prov.by Shengchao SHI,Jianyi FENG,Lei FU on 27 August,2019.CIB5133380017,CIB5133380019,CIB5133380021,CIB5133380024-26,CIB5133380047 were collected from Guxue Town (Derong County 28.424694° N,99.263363° E,2114 m a.s.l.),Sichuan Prov.by Gang WANG,Yinmeng HOU,Fuxuan LIANG,Yusong TANG on 26 August,2019.Etymology
The specific epithet jinjiangensis refers to the type locality of the species,were collected in the middle reaches of the Jinsha River across the West of Sichuan and Yunnan Province,China.Diagnosis
Gekko jinjiangensis
sp.nov.
can be distinguished from its congeners by a combination of following characters:size small (SVL 50.2-61.6 mm,n
=13);nares in contact with rostral;interorbital scales between anterior corners of the eyes 20-24;ventral scales between mental and cloacal slit 146-169;midbody scale rows 111-149;ventral scale rows 31-47;subdigital lamellae on first toe 8-11,on fourth toe 11-15;no webbing in the fingers and toes;with tubercles on upper surface of fore and hind limbs;precloacal pores 4-5 in males and absent in the females;postcloacal unilateral tubercles 1-2;dorsal surface of body with 8-9 large greyish brown markings between nape and sacrum.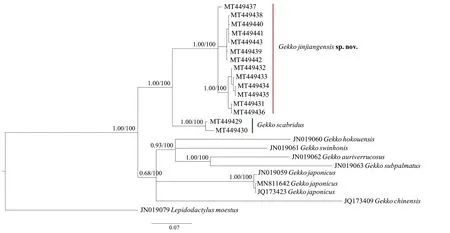
Figure 1 Phylogram based on the Bayesian analysis.Number above branches are Bayesian posteriorprobabilities (> 50%) and maximum likelihood bootstrap values,respectively.
Figure 2 Holotype (CIB5334220115,male) of Gekko jinjiangensissp.nov.A:Lateral view of head.B:dorsal view of snout.C:ventral view of chin.D:ventral view of left hand.E:ventral view of right hand.F:dorsal view of middle body.G:ventral view of precloacal region,showing five precloacal pores.H:lateral view of basal tail.
Description of holotype
Adult male,SVL 54.7 mm;tail partially regenerated,TaL 46.3 mm (regeneration tail);head length longer than width (HL/HW=1.20),slightly depressed(HH/HL=0.46),distinct from neck;rostral irregular polygon,wider than highth (RW/RH=1.59) and relatively longer than mental (RW/MW=1.17),rostral groove present;rostral in contact with nostril,first supralabial and nasorostral;nostril suborbicular,touching rostral,first supralabial and three nasals (nasorostral,supranasal,postnasal);nasorostral enlarge in contact each other;supranasal equal to postnasal;internasal oval;snout moderate (SE/HL=0.39),larger than eye diameter(SE/ED=1.43);snout region medially concave from interorbital region to rostral;granular scales on anteriodorsal surface on head larger than those on posterior surface;preorbitals 12/11(L/R);interorbitals 21;eye relatively large (ED/HL=0.27);pupil vertical with crenulated margin;ear opening oval,obliquely oriented,smaller than eye (EAD/ED=0.35);mental pentagons,equal in width and length (MW/ML=1.0);two enlarged postmentals,hexagonal,longer than width;postmentals in contact with mental and first infralabials anteriorly and six gular scales posteriorly;supralabial to midpoint of orbit 9/9(supralabial to angle of jaw 11/11);infralabials to midpoint of orbit 7/6 (infralabials to angle of jaws 9/8).Body slender,trunk relative long (AG/SVL=0.40),dorsoventrally depressed in cross section;dorsal scales on body smooth,round or oval,granular,juxtaposed;dorsal tubercles 3-4 times the size of dorsal scales,smooth,round to oval,convex,surrounded by 8-10 dorsal scales;dorsal tubercles extending from occiput region to base of tail;tubercles in 16 regular rows at midbody;ventrolateral fold weakly developed,without tubercles;43 ventral scales at midbody;ventral scales on abdomen much larger than dorsal or lateral scales,becoming smaller in chest and gular regions,flatted,subimbricated.Slightly enlarged precloacal scales bearing a continuous row of 5 precloacal pores;scales on dorsal surface of limbs larger than dorsum,smooth,round,or oval,granular,juxtaposed;scales on palms and soles smooth,round,or oval,granular,juxtaposed;scales on dorsal of digits,anterior surfaces of forearm,thigh,flatted,subimbricated.
Forelimbs and hindlimbs well developed,tubercles on fore and hind limbs are present,moderately long,slender;forearm and tibia moderately long;digits moderately expanded,both first finger and first toe,clawless,all remaining digits clawed;finger and toe webbing absent;subdigital lamellae,unnotched,undivided:9-10-10-11-9 (left manus),10-8-10-12-10 (right manus),11-11-11-13-12 (left pes),11-10-12-12-13 (right pes).Relative length of fingers:IV > III > V > II > I;relative length of toes:IV > III > V >II > I.
Tail regenerated;swollen at base;two postcloacal tubercle scales present on each lateral side;tail suddenly tapering from the basal swelling,slightly depressed,oval in section,covered above with small scales;tubercles on dorsal tail present only at base,similar with those on dorsal body,no large sharp tubercles present in rows on tail;dorsal scales of tail,smooth,oval,juxataposed to weakly subimbricated in regular transverserows;ventral scales much larger than dorsal,smooth,and imbricated,with enlarged subcaudal plates.
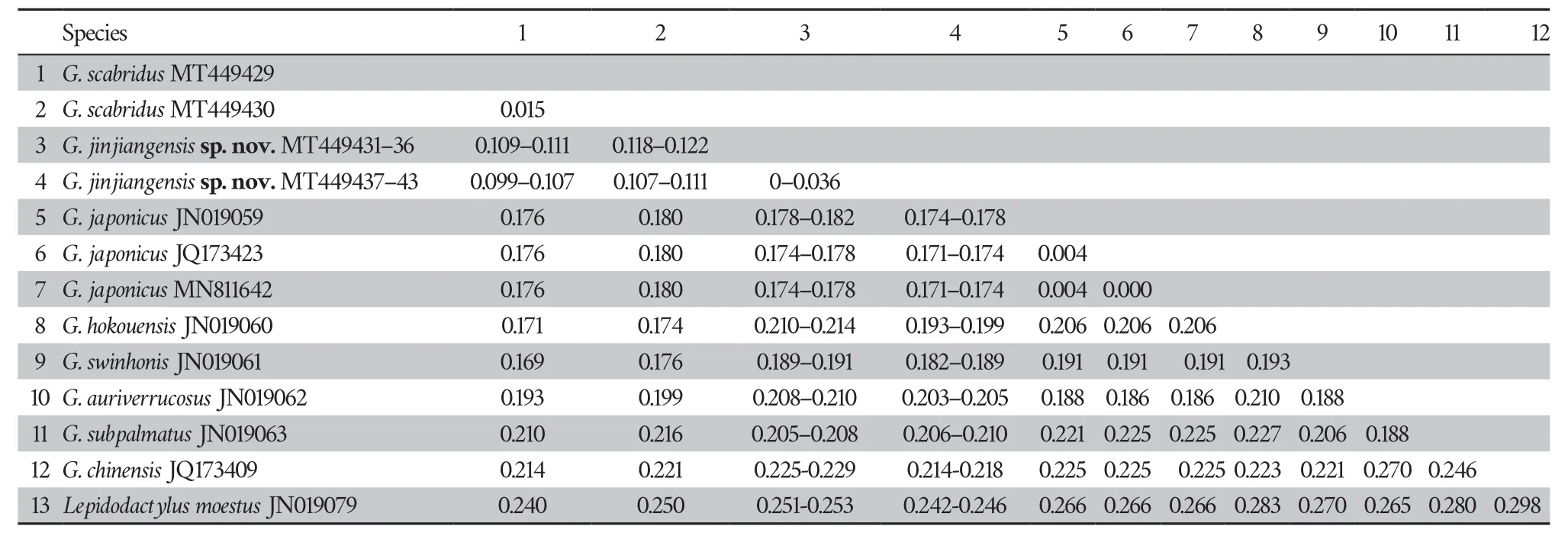
Table 3 Mean (min-max) uncorrected pairwise distances (%) within and between species of the Gekko japonicus group based on ND2 sequences.
Color of the holotype in life (Figure 2)
In life,dorsal surfaces of head,neck and body light grey,scattered with 8 large light and dark brown wide irregular W’-shaped patches from neck to the swollen section of tail;dorsal surfaces of limbs,flesh red.Dorsal part of limbs mottled with small,light blotches irregularly scattered;dorsal tail greyish,with single dark bands,regenerated portion yellow-brown without band;ventral skin creamy-yellow,ventral tail and regenerated tail creamy-white.Iris sliver with minute,dark brown reticulations,pupil solid black with a sliver edge.Color of holotype in preservative
In preservative (fixed in formalin,stored in 75% ethanol for 4 months),dorsal and lateral surfaces of head,body and limbs creamy-white with distinct dark grey flecks;tail similar to dorsal ground color with light grey bands and scatted small dark flecks;ventral body white,limbs,precloacal regions and tail creamy with minute dark grey pigment speckles on gular and limbs regions (Figure 3).
Figure 3 Holotype (CIB5334220115,male) of Gekko jinjiangensis sp.nov in preservation.A:dorsal view.B:ventral view.
Variation
The differences between the measurement data and the scales of the 13 specimens are shown in the Table 4.Male paratypes (CIB5133380021,CIB5334220090,CIB5334220088)with 4 distinct precloacal pores,holotype (CIB5334220115) with 5;paratypes CIB5133380021,CIB5133380026,CIB5133380024,CIB5334220114,CIB5133380047,CIB5334220089,CIB5133380025,CIB5133380017,CIB5133380019,CIB5334220090 and CIB5334220088 with one postcloacal tubercle,while paratype CIB5334220100 with two postcloacal tubercles similar with the holotype.A small internasal present in paratypes CIB5133380021,CIB5133380026,CIB5334220089,CIB5133380025,CIB5133380017,CIB5334220088,CIB5334220115,but absent in paratypes CIB5133380024,CIB5334220114,CIB5133380047,CIB5334220100,CIB5133380019,CIB5334220090.Dorsal coloration varies between specimens,paratypes CIB5133380019 and CIB5334220100 (Figure 4) dark brown,and the remaining are light colored.Dorsal surface of body with 8-9 large greyish brown markings between nape and sacrum.Among them the holotype and paratypes CIB5133380019,CIB5133380026,CIB5133380047,CIB5334220088,CIB5334220089,CIB5334220114 have 8 large greyish brown markings;the remaining type specimens are 9.Original tails with about 9-10 light brown and faint white transverse bands (CIB5133380024,CIB5133380047 have 10,CIB5133380021,CIB5133380026,CIB5334220090 have 9).The holotype and CIB5334220088,CIB5334220114,CIB5133380017,CIB5133380019,CIB5133380025 have an obscure dusky band from nostril through eye to earopening,the remaining type specimens have a clear and dark band.Regenerated tails (CIB5334220089,CIB5133380025,CIB5133380017,CIB5334220100,CIB5133380019,CIB5334220114,CIB5334220088 and CIB5334220115) shorter tail than original tails,and the scales on the ventral part of these tails are also more irregular.
Figure 4 Living female paratype of Gekko jinjiangensis sp.nov..Dorsolateral view of CIB5334220100.
Distribution and natural history
Gekko jinjiangensis
sp.nov.
is currently known from the middle reaches of the Jinsha River,Sichuan and Yunnan Province,China at elevations between 2000 -2476 m a.s.l.The new species is nocturnal,inhabits shrubs or dry rocky cliffs in Jinsha River valley,and also found on walls of building (Figure 5).The breeding season is currently uncertain,no gravid female found during field work from late August 2019 and late July 2020.Comparison.
The undescribed species can clearly differs from the remaining species of theG.japonicus
group by a unique suite of features.The newGekko
species differs from the members of theG.japonicus
group are as follows (Table 5).The new species can be easily distinguished fromG.aaronbaueri
by having fewer supralabials and infralabials (7-10 versus 13 or 14 and 6-9 versus 10 or 11,respectively),fewer interorbital scales (20-24 versus 34-37),dorsal tubercle rows at midbody (12-16 versus absent),more scale rows around midbody (111-149 versus 98-104),fewer lamellae under first toe(8-11 versus 14-17),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks and bands pattern.
Figure 5 Habitat of Gekko jinjiangensis sp.nov.A:macrohabitat,dryhot river valley in Benzilan Town,Deqin County,Yunnan Province.B,C,D:microhabitats.B:holotype CIB5334220115 found on small bushes in the Jinsha River valley.C:one individual hide in rock stretches on cliff after disturbance.D:one individual found on slope with bushes.
The new species differs fromG.adleri
by having fewer interorbital scales (20-24 versus 27-36),more dorsal tubercle rows at midbody (12-16 versus 7-11),no toes webbing (versus present),tubercles on fore limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 17-21),and a different head markings pattern.The new species differs fromG.auriverrucosus
by having a nostril touching rostral (versus not touching in latter),by having fewer interorbital scales (20-24 versus 25),postmentals enlarged(versus not enlarged in latter),more lamellae under fourth toe(11-15 versus 6-8),fewer precloacal pores in males (4 or 5 versus 8-11).The new species can be easily distinguished fromG.bonkowskii
by having fewer supralabials and infralabials (7-10 versus 12-14 and 6-9 versus 10 or 11,respectively),fewer interorbital scales (20-24 versus 26-27),dorsal tubercle rows at midbody (12-16 versus absent),no toes webbing (versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 6),tubercles on dorsal surface of tail(present versus absent),and a different dorsal flecks and bands pattern.The new species differs fromG.canhi
by having fewer supralabials and infralabials (7-10 versus 14 and 6-9 versus 10-12,respectively),fewer interorbital scales (20-24 versus 49 or 50),fewer scale rows at midbody (111-149 versus 205-227),fewer ventral scale rows at midbody (31-47 versus 49-51),fewer lamellae under first toe (8-11 versus 13-16),tubercles on fore limbs (present versus absent),tubercles on dorsal surface of tail(present versus absent),and a different dorsal flecks pattern.The new species differs fromG.chinensis
by having fewer interorbital scales (20-24 versus 35-48),dorsal tubercle rows at midbody (12-16 versus 10),no toes webbing (versus present),tubercles on fore limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 17-27);and a different dorsal flecks and bands pattern.The new species differs fromG.hokouensis
by having fewer interorbital scales (20-24 versus 30-33),postmentals enlarged(versus not enlarged),tubercles on limbs (present versus absent);no back flecked or blotched (versus present),and a different dorsal flecks and bands pattern.The new species differs fromG.kwangsiensis
by having fewer infralabials (6-9 versus 11-13),fewer interorbitals (20-24 versus 29-31);more dorsal tubercle rows at midbody (12-16 versus 9-11),fewer scales in a line from mental to the front of cloacal slit (146-169 versus 185-208),no toes webbing (versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 9-11).The new species differs fromG.liboensis
by having fewer supralabials and infralabials (7-10 versus 12 and 6-9 versus 11,respectively),fewer interorbital scales (20-24 versus 40),postmentals enlarged (versus not enlarged),dorsal tubercle rows at midbody (12-16 versus 10),more lamellae under fourth toe(11-15 versus 9),tubercles on limbs (present versus absent).The new species can be easily distinguished fromG.melli
by having fewer interorbitals (20-24 versus 34-40);dorsal tubercle rows at midbody (12-16 versus absent),postmentals enlarged(versus not enlarged),fewer scales in a line from mental to the front of cloacal slit (146-169 versus 181-200),no toes webbing(versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 9-11);tubercles on dorsal surface of tail (present versus absent),and a different head markings pattern.The new species can be easily distinguished fromG.nadenensis
by having fewer supralabials and infralabials(7-10 versus 12-14 and 6-9 versus 10-12,respectively),fewer interorbitals (20-24 versus 28-30);fewer scales in a line from mental to the front of cloacal slit (146-169 versus 175-185),fewer lamellae under first toe (8-11 versus 13-15),no toes webbing(versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 6);tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks and bands pattern.The new species differs fromG.palmatus
by having fewer supralabials (7-10 versus 11-15),fewer interorbitals (20-24 versus 27-36);no toes webbing (versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 23-30),and a different dorsal flecks pattern.The new species can be easily distinguished fromG.
scientiadventura
by having fewer supralabials (7-10 versus 12-14),fewer interorbitals (20-24 versus 41-51),dorsal tubercle rows at midbody (12-16 versus absent),more scales in a line from mental to the front of cloacal slit (146-169 versus 118-140),fewer lamellae under first toe (8-11 versus 12-15);no toes webbing(versus present),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks and bands pattern.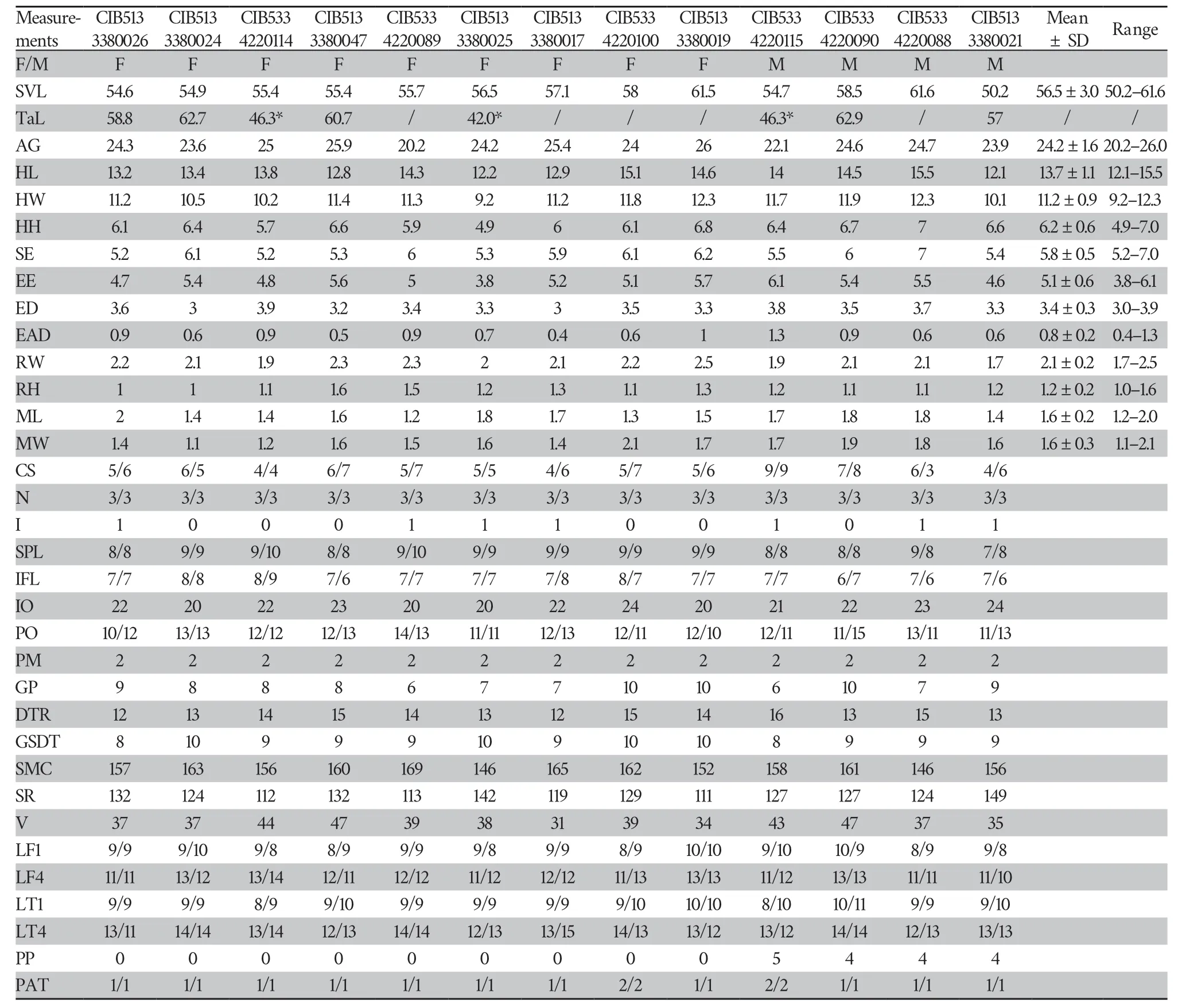
Table 4 The measurements (in mm) and morphological characters of the type series of Gekko jinjiangensissp.nov.(“F/M”means the gender female and male respectively.“*”means the length of regeneration tail;“/”means severed tail).
The new species differs fromG.subpalmatus
by having fewer interorbitals (20-24 versus 32),postmentals enlarged (versus not enlarged),fewer dorsal tubercle rows at midbody (12-16 versus absent),fewer ventral scale rows at midbody (31-47 versus 48),more lamellae under fourth toe (11-15 versus 7-10),no toes webbing (versus present),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks pattern.The new species differs fromG.swinhonis
by having postmentals enlarged (versus not enlarged),more dorsal tubercle rows at midbody (12-16 versus 6-8),more lamellae under fourth toe (11-15 versus 6-9),fewer precloacal pores in males (4 or 5 versus 7-9);and a different dorsal flecks and bands pattern.The new species can be easily distinguished fromG.sengchanthavongi
by having fewer interorbitals (20-24 versus 28-32),dorsal tubercle rows at midbody (12-16 versus absent),fewer scales in a line from mental to the front of cloacal slit(146-169 versus 175-184),no toes webbing (versus present),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks pattern.
G.shibatai
by having fewer infralabials (6-9 versus 10-14),fewer interorbitals (20-24 versus 37-52),postmentals enlarged (versus not enlarged),tubercles on limbs (present versus absent),more precloacal pores in males (4 or 5 versus 0-3),and a different dorsal flecks and bands pattern.The new species can be easily distinguished fromG.similignum
by having fewer supralabials and infralabials (7-10 versus 12-14 and 6-9 versus 11,respectively),fewer interorbitals(20-24 versus 46-48),postmentals enlarged (versus not enlarged),more dorsal tubercle rows at midbody (12-16 versus 11),no toes webbing (versus present),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 17),and a different dorsal flecks and bands pattern.The new species differs fromG.taibaiensis
by having fewer interorbitals (20-24 versus 28),more lamellae under first toe(11-15 versus 6 or 7),and more lamellae under fourth toe (11-15 versus 7 or 8),and a different dorsal flecks pattern.The new species differs fromG.tawaensis
by having fewer supralabials and infralabials (7-10 versus 15 and 6-9 versus 13,respectively),postmentals enlarged (versus not enlarged),fewer intersupranasals (0-1 versus 2-2),dorsal tubercle rows at midbody (12-16 versus absent),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks pattern.The new species differs fromG.thakhekensis
by having fewer supralabials and infralabials (7-10 versus 12-14 and 6-9 versus 10-11,respectively),dorsal tubercle rows at midbody (12-16 versus absent),no toes webbing (versus present),tubercles on limbs (present versus absent),tubercles on dorsal surface of tail(present versus absent),and a different dorsal flecks and bands pattern.The new species can be easily distinguished fromG.truongi
by having fewer supralabials and infralabials (7-10 versus 13-15 and 6-9 versus 11-13,respectively),fewer interorbitals(20-24 versus 45-48),dorsal tubercle rows at midbody (12-16 versus absent),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 10 or 11);tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks pattern.The new species differs fromG.vertebralis
by having fewer infralabials (6-9 versus 10-15),fewer interorbitals (20-24 versus 35-50),postmentals enlarged (versus not enlarged),tubercles on limbs (present versus absent),more precloacal pores in males(4 or 5 versus 0 or 1);tubercles on dorsal surface of tail (present versus absent),and a different dorsal flecks and bands pattern.The new species differs fromG.wenxianensis
by having fewer supralabials and infralabials (7-10 versus 12 and 6-9 versus 11,respectively),more dorsal tubercle rows at midbody (12-16 versus 10),more lamellae under first toe (11-15 versus 6),more lamellae under fourth toe (11-15 versus 9),tubercles on fore limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 6-8),and a different dorsal flecks and bands pattern.The new species differs fromG.yakuensis
by having fewer supralabials (7-10 versus 12-13),postmentals enlarged (versus not enlarged),tubercles on limbs (present versus absent),fewer precloacal pores in males (4 or 5 versus 6-8),and a different dorsal flecks and bands pattern.G.japonicus
is the species with the highest morphological similarity to the new species,but the phylogenetic relationships and genetic distance make it easy to distinguish between the two species (uncorrectedp
-distance 17.1%-18.2%).Morphologically,the new species differs fromG.japonicus
by having fewer interorbital scales (20-24 versus 32-35);no toes webbing (versus relative obvious);the tubercle scales on thigh distinct (versus generally without) (Tianet al
.,1986;Stejneger,1901).G.scabridus
is the species with the closest phylogenetic relationship and genetic distance to the new species.The two species are easily distinguished by their morphological features(Figure 6).The new species differs fromG.scabridus
by having fewer interorbitals (20-24 versus 30),fewer dorsal tubercles rows at midbody (12-16 versus 17-21),more lamellae under fourth toe (11-15 versus 7-9) (Figure 7),fewer precloacal pores in males (4 or 5 versus 10-15) (Figure 8),no toes webbing (versus relative obvious),large markings on the back (versus dark brown and mesh spots).
Figure 6 Holotype (CIB5334220115,male) of Gekko jinjiangensis sp.nov.in life and G.scabridus (CIBYN201909200,male) from Yanbian,Sichuan Province:A,C:dorsal view.B,D:ventral view.

Figure 7 Scalation on head,trunk,and digits of Gekko jinjiangensis sp.nov.(holotype CIB5334220115) and G.scabridus (CIBYN201909200).A:lateral view of head.B:dorsal view of snout.C:ventral view of chin of Gekko jinjiangensis sp.nov.D:lateral view of head.E:dorsal view of snout.F:ventral view of chin of G.scabridus.G:dorsal view of trunk.H:ventral view of left hand.I:ventral view of right foot of Gekko jinjiangensis sp.nov.J:dorsal view of trunk.K:ventral view of right hand.L:ventral view of left feet of G.scabridus.
4.Discussion
Yang and Rao (2008) recordedG.japonicus
from Benzilan of Deqin,Yunnan Prov.,which is the type location of the new species.This record should be referred to the new species as it matches the diagnosis of the new species morphologically.Besides,Yang and Rao (2008) noted that this species is common,andG.jinjiangensis
sp.nov.is common at its distribution area during our filed work last near two months (August 2019 and July 2020).Detailed distribution range,population size and states,and threats of the species is still not known enough,further surveys are recommended to better understand the distribution and population status of the species.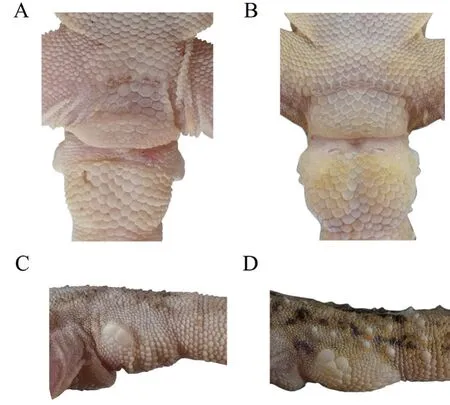
Figure 8 Male sexual character and basal tail of Gekko jinjiangensis sp.nov.(holotype CIB5334220115,male) and G.scabridus (CIBYN201909200,male).A:precloacal region of Gekko jinjiangensis sp.nov.B:precloacal region of G.scabridus.C:lateral view of basal tail of Gekko jinjiangensis sp.nov.D:lateral view of basal tail of G.scabridus.
The new species is the highest distributed species ofGekko
from the Qinghai-Tibet Plateau with a highest elevation range(2000 and 2476 m a.s.l.),the major habitat ofGekko jinjiangensis
sp.nov.is the dry-hot river valley in the middle of Jinsha River.We speculate that the reason for the radiation to the Tibetan Plateau is that the meridional dry hot river valley provides better hydrothermal conditions.The breeding period is presumed to be before July (May-June),as there are no females carrying eggs at July and August,but at the same time,more young individuals were found in the wild.During the field investigation,far more females were encountered than males.Two possible explanations to this phenomenon are possible high parthenogenesis rate and female biased sex determination during embryogenesis due to higher environmental temperature (Pieau,1996).At present,China (20 species) has the highest diversity ofGekko
in East and Southeast Asia,followed by the Philippines(17 species),Thailand (16 species) and Vietnam (14 species)(Uetz,2020).Among them,12 species are endemic to China.TheGekko
species of China belong to 4 gecko subgenera:Japonigekko
,Gekko,Ptychozoon
,Archipelagekko
(Woodet al.
,2020),mostly distributed in tropical and subtropical regions south of the Qinling Mountains,fewer in north regions of the temperate zone.The descriptions of the new species indicated that widespread species such as theG.japonicus
andG.swinhonis
may include cryptic species and need to be checked seriously.So,we suggest that a broader sampling strategy and molecular re-identification is necessary to clarify the taxonomy and phylogeny of Chinese geckos.Acknowledgements
This study was supported by the Second Tibetan Plateau Scientific Expedition and Research Program (STEP,2019QZKK05010503),the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment,China (2019HJ2096001006) to Feng XIE and Yin QI.We thank Jianyi FENG,Lei FU for their help in the field.We are also grateful to Mr.Xianguang GUO and Ke JIANG for their very helpful advices,and the local forestry administration for their support and help in our field work and issuing relevant permits.杂志排行

Asian Herpetological Research的其它文章
- An Integrative Taxonomy of Amphibians of Nepal:An Updated Status and Distribution
- Genetic Diversity and Population Structure of the Oriental Garden Lizard,Calotes versicolor Daudin,1802 (Squamata:Agamidae) along the Mekong River in Thailand and Lao PDR
- Species Diversity,Distribution,and Microhabitats of Anurans on Mt.Kalo-Kalo of the Mt.Kalatungan Range Natural Park,Bukidnon,Philippines
- Species Account of Anurans from the Western Slope of Mt.Kitanglad,Mindanao Island,Philippines
- Behavioral and Neurogenomic Responses to Acoustic and Visual Sexual Cues are Correlated in Female Torrent Frogs
- The Influence of Environmental Factors on the Behavior and Mortality Risk of Polypedates megacephalus Tadpoles