Aurora A 激酶抑制剂Alisertib 诱导结直肠癌细胞自噬的作用及与p53表达间的关系
2021-03-26任宝军剧永乐封静陈淑香刘家旋罗振涛王家智杨帆李德智董博业耿岩
任宝军,剧永乐,封静,陈淑香,刘家旋,罗振涛,王家智,杨帆,李德智,董博业,耿岩*
在世界范围内结直肠癌(CRC)是最常见的恶性肿瘤之一,发病率居第三位,死亡率居第二位[1],尤其晚期结直肠癌的治疗效果不佳,仍缺乏有效的靶向药物。Alisertib 是第二代Aurora A 激酶(AURKA)抑制剂,研究发现对胶质母细胞瘤、膀胱癌、胰腺癌、卵巢癌等多种肿瘤具有抑制细胞增殖和阻止细胞周期发展的作用[2-4]。但Alisertib对CRC 的诱导细胞自噬的作用尚不清楚。p53 是一种抑癌基因,在CRC 的肿瘤形成机制中发挥重要作用,其与Alisertib 的诱导细胞自噬的作用的关系值得研究。本研究应用Caco-2 和HT29 细胞进行实验,探讨ALS 对不同p53 表达状态的CRC 细胞的诱导细胞自噬的作用。
1 材料与方法
1.1 主要试剂和仪器
Alisertib、SB202190(选择性p38 MAPK 抑制剂和自噬诱导剂)购自美国Selleckchem公司;Cyto-ID®自噬检测试剂盒及膜联蛋白V:藻红蛋白(PE)凋亡检测试剂盒购自美国Enzo Life Sciences 公司;p53、p-PI3K(Tyr458)、PI3K、p-p38 MAPK(Thr180/Tyr182)、p38 MAPK、p-Akt(Ser473)、Akt、LC3-I/II、Western Blotting 底物购自美国Cell Signaling Technology 公司。凝胶成像系统4000R PRO(美国Kodak 公司),流式细胞仪(美国BD 公司),细胞培养箱(美国Thermo Fisher 公司)。
1.2 细胞培养
Caco-2 和HT29 细胞来自美国ATCC。用含10%热灭活的胎牛血清以及抗生素的DMEM 培养基,于5%CO2、37℃及95%空气的恒温细胞培养箱中进行细胞培养。
1.3 流式细胞术检测细胞自噬率、凋亡率
取对数生长期的Caco-2 和HT29 细胞按密度3×105及6×105个/培养皿接种于60 mm 培养皿,24 h后用 0.1、1、5 μmol/L Alisertib 处理细胞,或先用10 μmol/L SB202190 处理细胞,1 h 后再用 5 μmol/L Alisertib处理细胞。对照组为同浓度的二甲基亚砜(DMSO)。作用24 h 后按照Cyto-ID®自噬检测试剂盒或膜联蛋白V:藻红蛋白(PE)凋亡检测试剂盒说明书处理细胞,流式细胞仪检测自噬率、凋亡率。
1.4 Western Blotting 法检测细胞相关调节蛋白的表达
取对数生长期的Caco-2 和HT29 细胞按密度3×105及6×105个/培养皿接种于60 mm 培养皿,24 h后用0.1、1、5 μmol/L Alisertib 处理细胞,作用 48 h后按照Western Blotting 试剂盒说明书处理。
1.5 统计学方法
以β-actin 作为内参,应用Image Lab 软件对Wester Blotting 条带进行灰度值测量,测算各蛋白相对表达水平。余数据用均数±标准差(Mean±SD)表示。多重比较应用Prism 软件通过单因素方差分析(ANOVA),然后进行Tukey 的多重比较。以P<0.05 为差异有统计学意义。
2 结 果
2.1 Alisertib 诱导了 Caco-2 和 HT29 细胞发生细胞自噬
对照组Caco-2 细胞自噬率为8.1%,实验组0.1 μmol/L Alisertib组和1 μmol/L Alisertib组细胞自噬率增加,分别为26.4%和26.8%(P<0.01,图1)。对照组HT29 细胞自噬率为7.3%,实验组1 μmol/L Alisertib组和5 μmol/L Alisertib组细胞自噬率增加,分别为36.8%和42.6%(P<0.01,图1)。
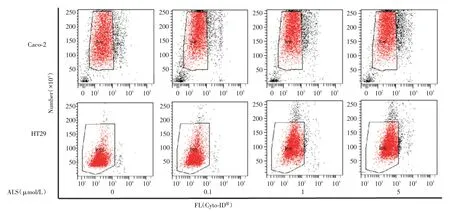
图1 ALS 诱导Caco-2 和HT29 细胞发生自噬 将Caco-2 和HT29 细胞暴露于0.1、1 和5 μmol/L ALS 24 h,然后进行流式细胞术分析。流式细胞斑点图显示通过Cyto-ID 染色的发生自噬的Caco-2 和HT29 细胞
2.2 Alisertib 调节 Caco-2 和 HT29 细胞中的 PI3K/Akt 和 p38 MAPK 信号通路
与对照组比较,1 μmol/L Alisertib组和5 μmol/L Alisertib 组 Caco-2 细胞的 p-PI3K/PI3K 的比值分别降低为对照组的62.6%和45.2%;0.1 μmol/L Alisertib组、1 μmol/L Alisertib 组和 5 μmol/L Alisertib 组Caco-2 细胞p-Akt/Akt 的比值分别降低为对照组的50.4%、30.1%和38.2%(P<0.01,图2);1 μmol/L Alisertib组和 5 μmol/L Alisertib 组 Caco-2 细胞的 p-p38/p38的比值分别降低为对照组的49.0%和30.4%(图2)。并且0.1 μmol/L ALS组、1 μmol/L ALS组和5 μmol/L ALS 组 Caco-2 细胞 LC3-Ⅱ/LC3-I 的比值较对照组分别升高67.3%、32.6%和46.8%(P<0.01,图2)。
在Caco-2 细胞中p53 蛋白表达缺失(图3)。而在 HT29 细胞中,1 μmol/L Alisertib 组和 5 μmol/L Alisertib 组p53 蛋白的表达水平升高为对照组的1.4 倍和1.5 倍(P<0.05,图3)。
与对照组比较,1 μmol/L Alisertib组和5 μmol/L Alisertib 组HT29细胞的p-PI3K/PI3K的比值分别降低为对照组的70.9%和66.3%(P<0.001,图3);0.1 μmol/L Alisertib 组、1 μmol/L Alisertib 组和5 μmol/L Alisertib 组 HT29 细胞的 p-Akt/Akt 的比值分别降低为对照组的81.4%、85.2%和27.2%(P<0.01,图3);0.1 μmol/L Alisertib组、1 μmol/L Alisertib组和5 μmol/L Alisertib 组 HT29 细胞的 p-p38/p38 的比值较对照组分别升高62.7%、2.1 倍和2.0 倍(P<0.05,图3);1 μmol/L ALS 组和 5 μmol/L ALS 组 HT29 细胞的LC3-Ⅱ/LC3-I 的比值较对照组分别升高78.5%和84.8%(P<0.01,图3)。
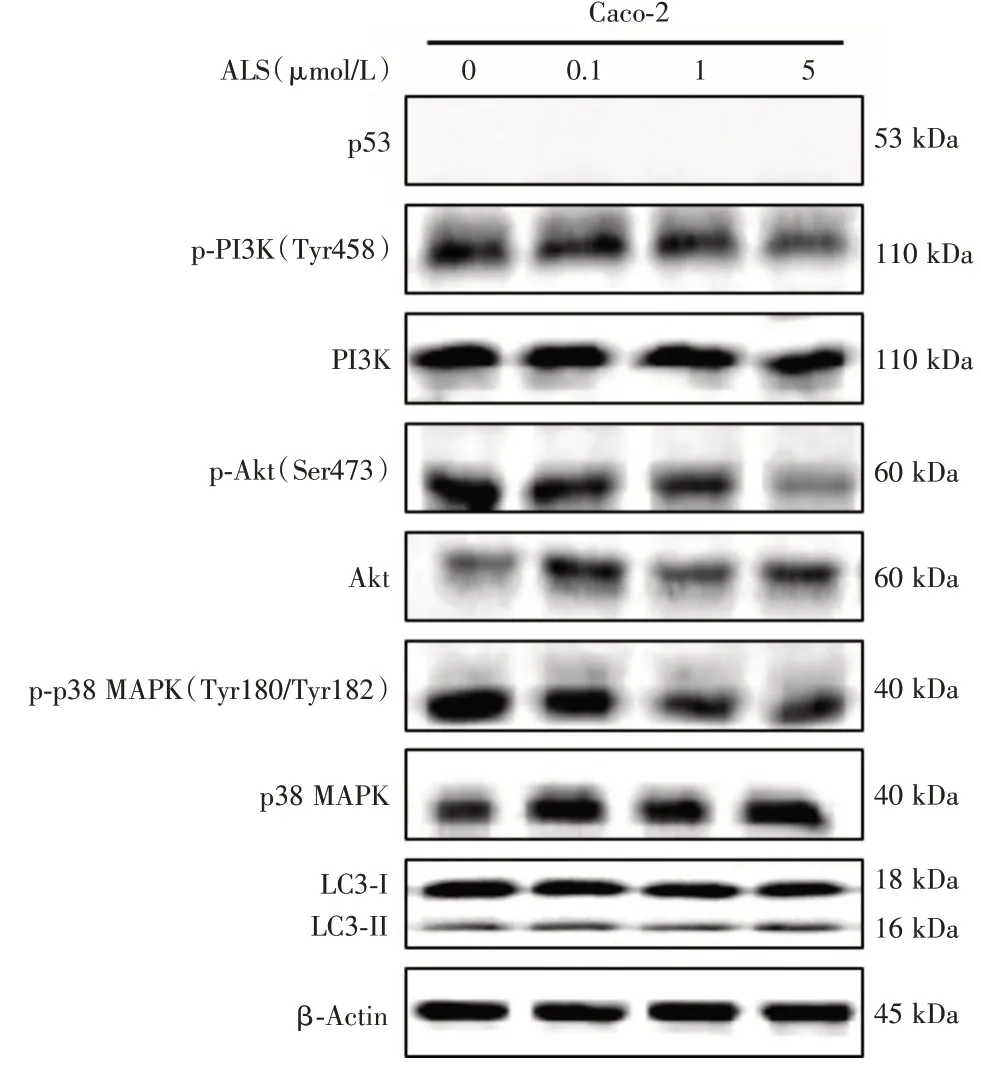
图2 ALS 对Caco-2 细胞中自噬通路上关键调节蛋白分子的表达水平的影响 ALS 处理Caco-2 细胞后通过Western Blotting 法测定 PI3K、Akt、p38 MAPK 蛋白表达水平及磷酸化水平,以及LC3-I 和LC3-II 的蛋白表达水平,上图为蛋白表达的代表性印迹条带
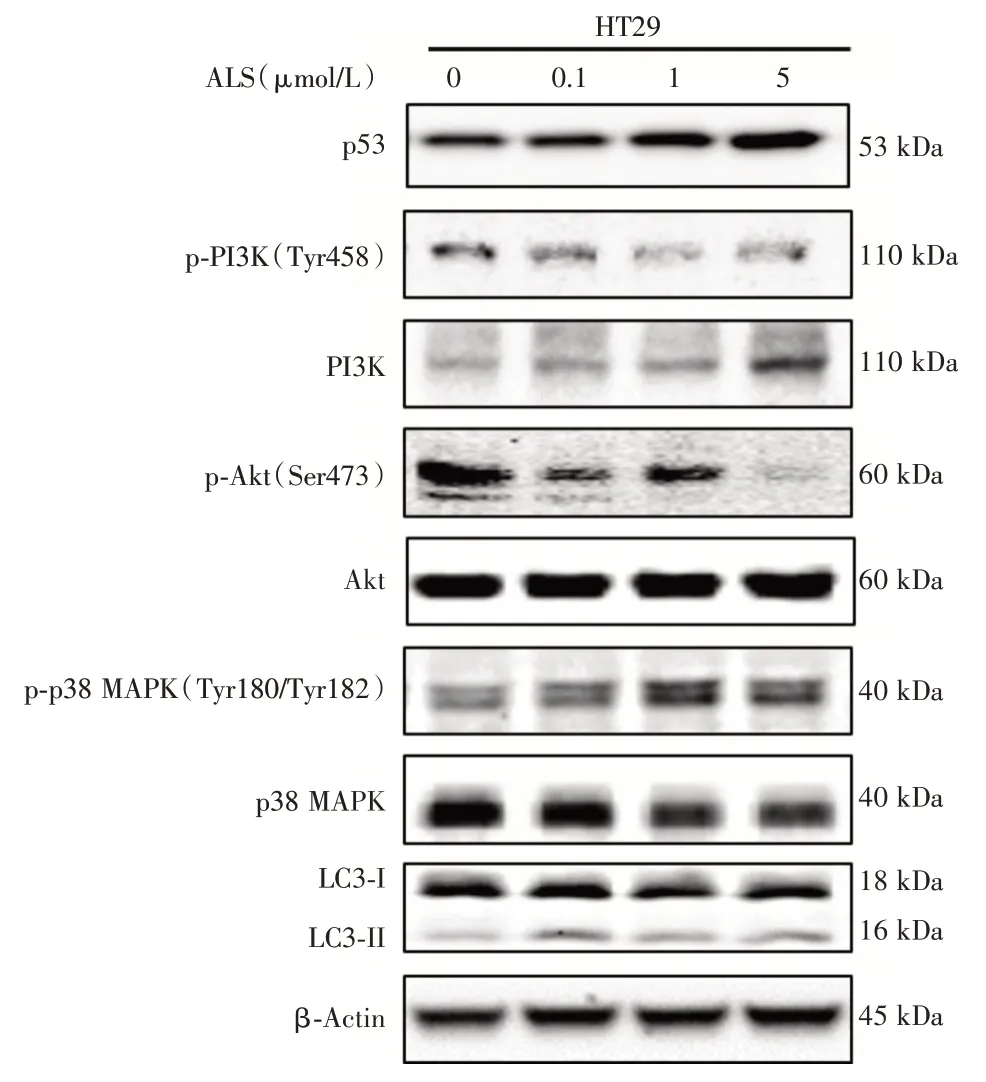
图3 ALS 对HT29 细胞中自噬通路上关键调节蛋白分子的表达水平的影响 ALS 处理HT29 细胞后通过Western Blotting 法测定 PI3K、Akt、p38 MAPK 蛋白表达水平及磷酸化水平,以及LC3-I 和LC3-II 的蛋白表达水平,上图为蛋白表达的代表性印迹条带
2.3 在HT29 和Caco-2 细胞中Alisertib 诱导的凋亡和自噬之间存在交互影响
在HT29和Caco-2细胞中,与5 μmol/L Alisertib组比较,10 μmol/L SB202190+5 μmol/L Alisertib 组细胞自噬率升高,分别较5 μmol/L Alisertib 组细胞自噬率升高88.8%和86.6%(P<0.001)。
在 HT29 细胞中,与 5 μmol/L Alisertib 组比较,10 μmol/L SB202190+5 μmol/L Alisertib 组细胞凋亡率升高64.7%(P<0.001)。而在Caco-2 细胞中,10 μmol/L SB202190+5 μmol/L Alisertib组较5 μmol/L Alisertib 组的细胞凋亡率无明显变化。
3 讨 论
结直肠癌现有的治疗措施包括外科手术、放疗、化疗等,但晚期结直肠癌患者总体治疗效果较差,亟需新的精准的靶向药物治疗。Alisertib(ALS)是选择性抑制Aurora A 激酶的小分子药物,对前列腺癌等实体瘤、白血病有很强的抗癌作用[5,6]。ALS是否引起CRC细胞自噬性死亡值得进一步研究。
自噬是一种细胞内的分解代谢的过程。损坏的、功能失调的或过剩的细胞器和蛋白被吞噬、重利用以维持细胞正常的代谢、生存和内稳定[7]。过度自噬可导致细胞死亡。在某些条件下,自噬通过增加程序性细胞死亡来抑制肿瘤形成[8]。有多个通路参与调节细胞自噬,包括正向调节通路(PI3K/Akt 和p38 MAPK 信号通路),发挥抑制自噬的作用,以及负向调节通路(AMPK 和p53 信号通路),发挥促进自噬的作用[9-11]。
本研究显示实验组0.1 μmol/L Alisertib 组和1 μmol/L Alisertib 组较对照组Caco-2细胞自噬率增加,分别为26.4%和26.8%,同样地,实验组1 μmol/L Alisertib 组和 5 μmol/L Alisertib 组较对照组 HT29细胞自噬率增加,分别为36.8%和42.6%。提示ALS诱导了Caco-2 和HT29 细胞发生自噬。进一步检测细胞自噬通路上的关键蛋白分子的表达情况,发现在分子水平ALS 引起Caco-2 和HT29 和细胞的 p-PI3K/PI3K、p-Akt/Akt 比值明显降低,LC3-II/LC3-I 的比值明显升高。这些结果表明引起自噬的潜在分子学机制可能为对PI3K/Akt/mTOR 信号的抑制。有趣的是,ALS 对p38 MAPK 信号通路的修饰作用有所不同,引起Caco-2 的p-p38/p38 的比值下降,而HT29 的p-p38/p38 比值升高。这一现象可能与p38 MAPK 有四个不同的亚型有关,包括p38 α、p38 β、p38γ和p38δ。研究显示在哺乳动物中,其上游调节因子及下游靶点随着细胞类型及刺激因子的不同而不同[12]。此外,HT29 细胞的p53 基因没有突变,而Caco-2 细胞的p53 基因有突变,并且不能表达p53 蛋白。这些特征可能引起两个细胞系的p38 MAPK 信号通路产生不同的变化。其他研究显示,在胃癌细胞[13]、胰腺癌细胞[3]、骨肉瘤细胞[14]、乳腺癌细胞[15]和卵巢癌细胞[4]中Alisertib 可能通过抑制PI3K/Akt/mTOR、p38 MAPK或Erk1/2 信号通路而激活AMPK 信号通路促进了细胞自噬。
细胞自噬和凋亡均是高度保守和严密调节的细胞过程,通常这两个生物学过程包含相似的调节通路,甚至共享相同的启动子和效应子。越来越多的证据表明,自噬和凋亡的交互作用可协同影响以使细胞选择二者中的某一途径,决定细胞的命运,以达到内部平衡。我们的研究通过应用SB202190(选择性p38 MAPK 抑制剂和自噬诱导剂)进行干预,结果表明自噬诱导剂SB202190 在Caco-2 和HT29 细胞中均增强了Alisertib 诱导的细胞自噬,但在HT29 细胞中明显增强了Alisertib 诱导的细胞凋亡,而在Caco-2 细胞中,细胞凋亡率无明显变化。这些对细胞自噬和凋亡的差异效应可能取决于细胞类型的差异。由此可以推测,某些重要的调节分子和信号通路协同介导了这一复杂的凋亡和自噬之间的交互作用,包括PI3K/Akt、p38 MAPK 信号通路及p53 是否表达。
综上所述,Alisertib 诱导了 Caco-2 和 HT29 细胞发生细胞自噬。此过程涉及了多个信号通路的参与,包括 PI3K/Akt、p38 MAPK 信号通路,并可能与p53 蛋白是否表达有关。Alisertib 可能代表了一类新的有潜力的治疗CRC 的靶向药物。
