荒漠河岸多枝柽柳灌丛碳氮磷化学计量特征及其影响因素
2021-03-25张晓龙周继华来利明姜联合郑元润
张晓龙, 周继华, 来利明, 姜联合, 郑元润*
1.山西财经大学资源环境学院, 山西 太原 030006 2.中国科学院植物研究所, 北京 100093
生态化学计量学作为一门探索生态过程中能量和多重化学元素平衡的新兴学科,为探究植物生长和养分供应关系以及植物与环境之间化学元素的相互耦合性提供了一种综合方法[1-2]. 碳、氮、磷是植物生长发育和生理生态活动所需的重要元素,植物吸收氮元素、磷元素,同化碳元素,进而影响群落、生态系统的碳过程以及矿质元素的生物地球化学循环[3-4]. 相比于其他元素而言,碳、氮和磷元素的耦合作用更强,C∶N∶P计量比不仅能够反映植物的养分限制状况和适应策略,同时能反映出植物吸收氮和磷过程中的光合固碳能力[5-6]. 有研究表明,低C∶N和C∶P的植物倾向于采取高光合速率的竞争策略,而高C∶N和 C∶P的植物更倾向于采用低光合速率的强大防御生态策略[7-8]. 因此,基于生态化学计量特征的植物-环境关系研究,能更好地揭示不同环境植物群落养分获取及其对环境的适应机制[9]. 值得提出的是,尽管目前对森林、草原、荒漠和水生生态系统的植物化学计量特征进行了一些有意义的探讨[3,10-11],但对灌丛群落化学计量特征及其对环境梯度变化响应规律的研究仍较少,尤其缺乏对干旱区固沙灌丛的生态化学计量研究[12-13].
多枝柽柳(TamarixramosissimaLedeb.)具有很强的耐旱性和适应能力,在长期的植被-环境相互作用下发展成不同的群落类型,在维持当地生态系统稳定和提供关键生态系统服务方面发挥着独特的作用[14]. 有研究表明,在相对潮湿的河岸地带,多枝柽柳具有强烈的水分竞争和向土壤中分泌盐的能力,尤其会限制草本植物的生长,形成单优势种植物群落,从而降低当地生物多样性[15]. 然而,在相对干旱的荒漠地区,多枝柽柳通过根系的“提水作用”将水分从深层土壤和地下水输送到浅层土壤,水分和养分富集形成“沃岛效应”,通过“沃岛效应”为干旱荒漠地区动植物提供良好的栖息地,从而提高生物多样性[16]. 以往关于柽柳的研究多集中于植物群落生态学特性及其与环境因子关系方面,例如,地下水变化对柽柳群落多样性的影响[14,17]、土壤水分变化对柽柳群落特征的影响[18-19]、人工生态输水工程对荒漠河岸柽柳植被恢复的影响[20-21]以及柽柳沙包与环境因子之间的关系[22]等,而对柽柳群落的生态化学计量学研究比较缺乏[8,23]. 在当前环境条件下,尤其是在极端干旱的荒漠河岸地区,群落水平上柽柳群落碳氮磷化学计量及其相互作用有哪些特点?土壤水盐和养分等环境因子如何对柽柳群落碳氮磷化学计量特征产生影响?定量地揭示它们之间的相互作用关系将有助于深入认识荒漠河岸地区柽柳群落的养分限制状况和适应策略.
鉴于此,该文通过对黑河下游荒漠河岸地带多枝柽柳群落进行调查,分析群落水平下碳氮磷化学计量特征,探讨多枝柽柳群落碳氮磷化学计量格局与环境因子的关系,从而阐明多枝柽柳群落对极端干旱环境的适应性,以期为荒漠河岸柽柳植被恢复和柽柳群落多样性保护提供参考.
1 材料与方法
1.1 研究区概况
研究区域位于黑河下游荒漠河岸地带(42°06′N~42°07′N、101°00′E~101°03′E),地处中纬度温带大陆性干旱气候区,气候极端干旱[8]. 年均温8 ℃左右,多年平均降水量低于40 mm,75%以上的年降水量集中在7—8月,年蒸发量为 2 300~3 700 mm[14]. 下游荒漠河岸地带性土壤为灰棕漠土,两岸发育有林灌草甸土,由于成土过程受地下水影响较大,呈现一定的盐碱化[24]. 受河流补给地下水的影响,荒漠河岸林主要分布在河岸两边,以胡杨(Populuseuphratica)和柽柳(Tamarixramosissima)为主;外围戈壁荒漠以旱生和沙生类型的灌木为主,代表性植物有泡泡刺(Nitrariasphaerocarpa)、琵琶柴(Reaumuriasoongarica)、细枝盐爪爪(Kalidiumgracile)、膜果麻黄(Ephedraprzewalskii)等[25].
1.2 采样点设置与取样
2019年8月9—18日植物生长旺季,多枝柽柳处于夏花期(6—9月),在黑河下游乌兰图格沿河监测断面,沿垂直于河岸大致以500 m为间隔设置调查样地,共布设群落调查样地9个(见图1). 每个样地随机设置3个10 m×10 m的灌木样方,调查样方内所有物种,在灌木、草本层主要记录种类名称、株(丛)数、高度、冠幅、基径和盖度等群落特征;同时记录样地基本状况,包括样地经纬度、坡度、生境、地貌和土地利用等属性. 对于每个样方,每个物种地上生物量的测定采用样株收获法,每个物种取2~3株具有代表性的植株带回实验室,用毛刷刷净植株表面的尘土等杂质,于80 ℃恒温烘干至恒质量并称量,进而估算调查样方地上生物量. 获取生物量后,将植株粉碎后过0.149 mm筛,用于化学性质分析,植物化学性质采用质量百分比表示,植物TC、TN、TP含量测定参照ZHANG等[8]所述方法测定. 在与植物群落相对应的样地内,对其土壤理化性质(土壤含水量、容重、TC含量、TN含量、C∶N、速效磷含量、pH和电导率)进行测定,取样深度为50 cm,每个样地设3个重复,土壤理化属性测定参照ZHANG等[8]所述方法测定.

图1 研究区样地位置示意Fig.1 Locations of the sample plots
1.3 数据统计分析
植物群落TC、TN和TP含量是群落内物种TC、TN和TP含量的加权平均值[11-12].
群落TC含量(TC):
(1)
群落TN含量(TN):
(2)
群落TP含量(TP):
(3)
式中:S为物种数;Bi为物种i的相对地上生物量;Ci为物种i的TC含量,mgg;Ni为种物i的TN含量,mgg;Pi为物种i的TP含量,mgg.
采用SPSS 18.0软件进行数据统计分析,用单因素分析和LSD检验法对沿河不同样地多枝柽柳群落的盖度、生物量、TC、TN、TP含量及C∶N∶P计量比进行差异显著性检验(P<0.05),通过Pearson相关系数分析多枝柽柳群落TC、TN、TP含量及C∶N∶P计量比与土壤理化属性的关系(P<0.05). 为定量分析土壤因子对多枝柽柳群落碳氮磷化学计量特征的影响,采用排序法确定主要影响因子,DCA结果显示,所有排序轴梯度长度均小于3,因此采用冗余分析法(RDA)确定主要影响因子. 为避免冗余变量的影响,采用Monte Carlo检验(9 999次置换)检测多枝柽柳群落碳氮磷化学计量特征和土壤因子是否存在显著相关关系,排除影响不显著的变量(P>0.05). 采用偏冗余分析(pRDA)用于揭示土壤因子对多枝柽柳群落碳氮磷化学计量特征的单独影响与交互作用. 上述统计分析在CANOCO 5.0软件中完成[26].
2 结果与分析
2.1 荒漠河岸多枝柽柳群落基本特征
在沿河梯度上,多枝柽柳群落盖度(F=83.18,P<0.001)、地上生物量(F=63.52,P<0.001)在不同样地间均具有显著差异,群落盖度为16.19%~85.33%,地上生物量为161.49~2 812.81 gm2(见表1). 群落盖度和地上生物量随沿河距离的增加呈下降的变化趋势,地上生物量最大值出现在距河100 m处,而群落盖度最大值出现距河 1 300 m处(见表1).
2.2 荒漠河岸多枝柽柳群落碳氮磷化学计量特征
在沿河梯度上,多枝柽柳群落TC含量(F=10.05,P<0.001)、TN含量(F=12.24,P<0.001)、TP含量(F=44.73,P<0.001)、C∶N(F=12.90,P<0.001)、C∶P(F=9.65,P<0.001)和N∶P(F=5.11,P=0.002)在不同样地间均具有显著差异(见表2). 多枝柽柳群落TC含量为317.39~439.73 mgg,TN含量为21.27~43.61 mgg,TP含量为0.97~2.15 mgg,C∶N、C∶P和N∶P分别为9.08~16.52、182.07~410.06和14.55~27.20(见表2).
在沿河梯度上,多枝柽柳群落TC和TN含量均随沿河距离的增加呈显著下降的变化趋势,最大值出现在距河100 m处,分别为423.67和40.89 mgg;群落TP含量呈先下降后上升的变化趋势(见图2),最大值出现在距河300 m处,为2.09 mgg. C∶N随沿河距离的增加呈波动上升的变化趋势,最大值出现在距河 3 800 m处,为16.18;而C∶P和N∶P均大致呈先上升后下降的变化趋势,分别出现在距河 1 800 和 2 300 m处,分别为336.36和24.10(见图2).
2.3 多枝柽柳群落计量学特征与土壤因子的关系
在沿河梯度上,多枝柽柳群落TC含量、TN含量与土壤含水量、土壤TC含量、土壤TN含量和土壤速效磷含量均呈显著正相关,群落TP含量与土壤含水量、土壤TC含量、土壤TN含量和土壤pH均呈显著正相关,而群落TP含量与土壤电导率呈显著负相关(见表3). 群落C∶N与土壤容重、土壤C∶N均呈显著正相关,而与土壤含水量、土壤TC含量、土壤TN含量和土壤速效磷含量均呈显著负相关;群落C∶P与土壤电导率呈显著正相关,而与土壤含水量、土壤TC含量、土壤TN含量和土壤pH均呈显著负相关;群落N∶P 与土壤容重、土壤C∶N均呈显著负相关(见表3).
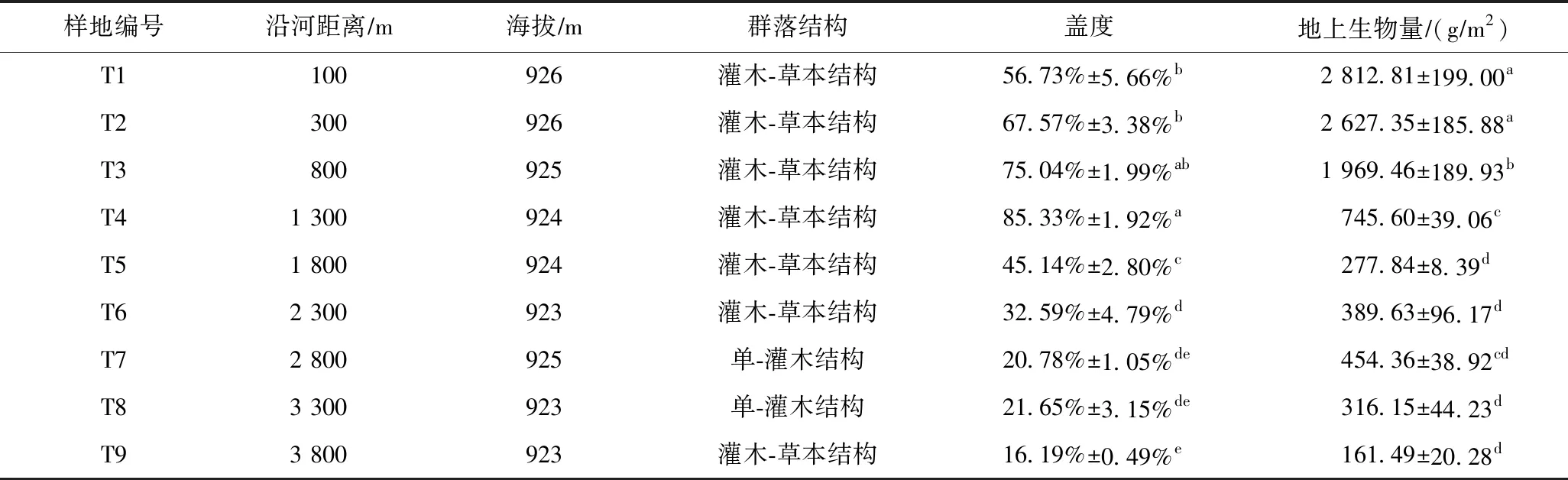
表1 黑河下游沿河多枝柽柳样地基本情况
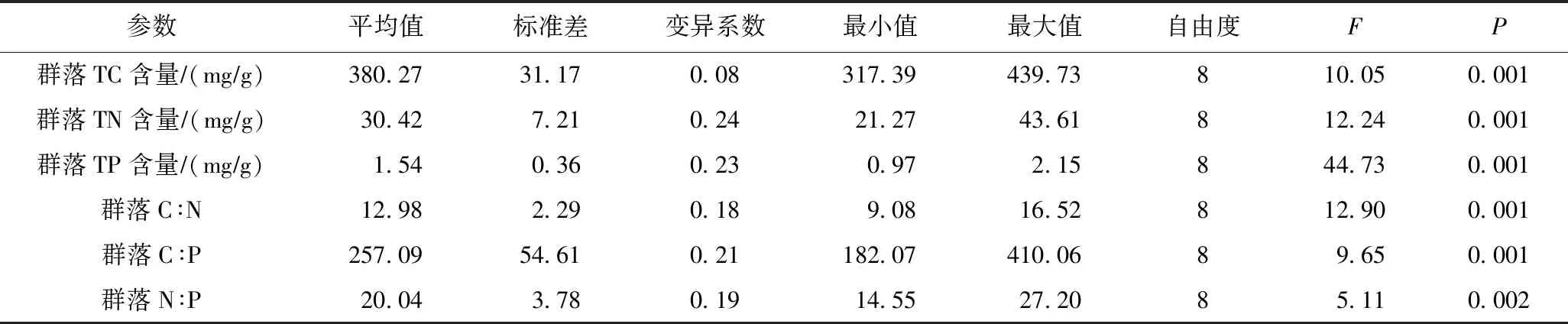
表2 荒漠河岸多枝柽柳样地间群落化学计量特征的方差分析
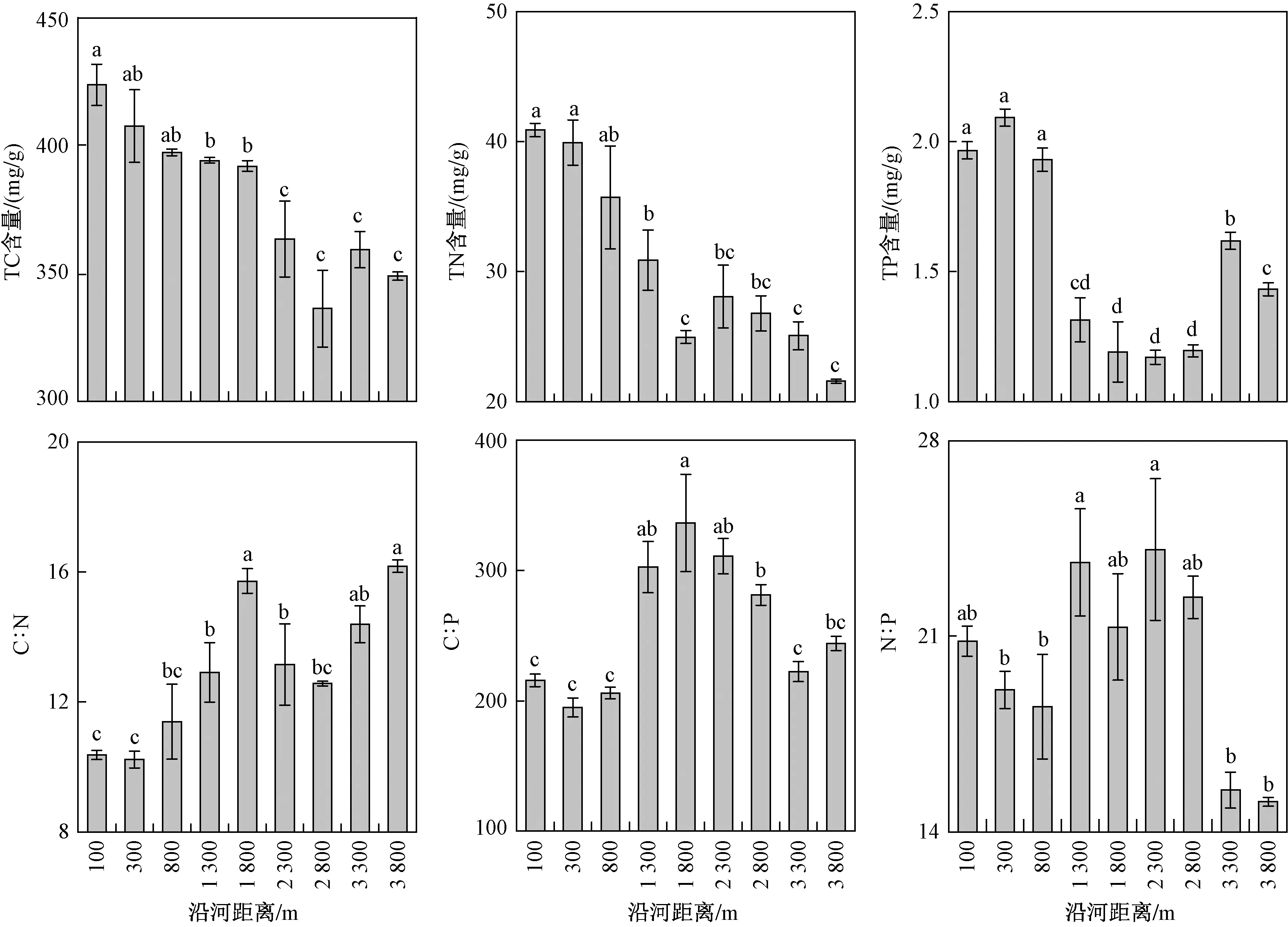
图2 多枝柽柳群落全碳、全氮、全磷及碳氮磷计量比随沿河梯度的变化Fig.2 Changes in the T. ramosissima community TC, TN, TP and C∶N∶P stoichiometric ratios along the river gradient
2.4 土壤因子对多枝柽柳群落计量学特征的影响
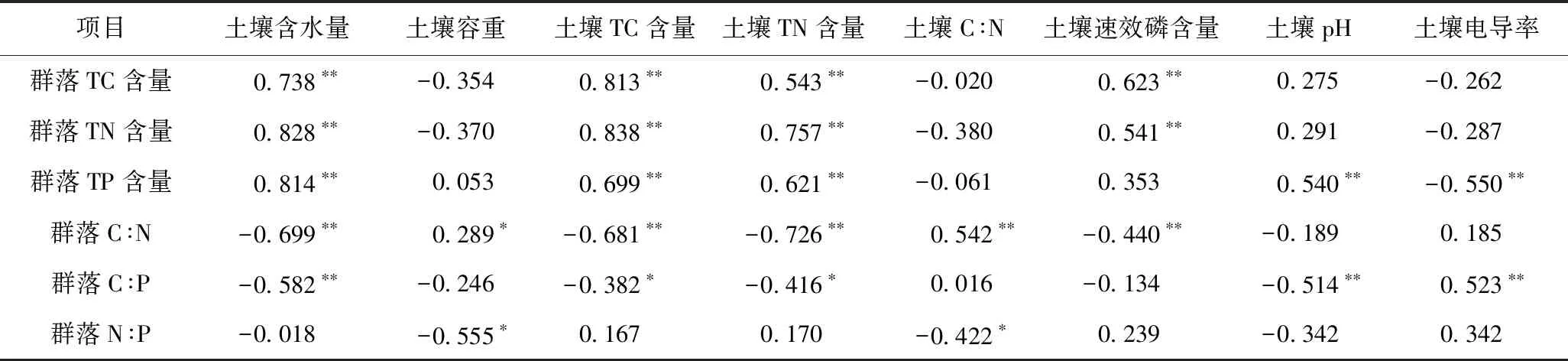
表3 多枝柽柳群落碳氮磷化学计量特征与土壤因子之间的相关关系
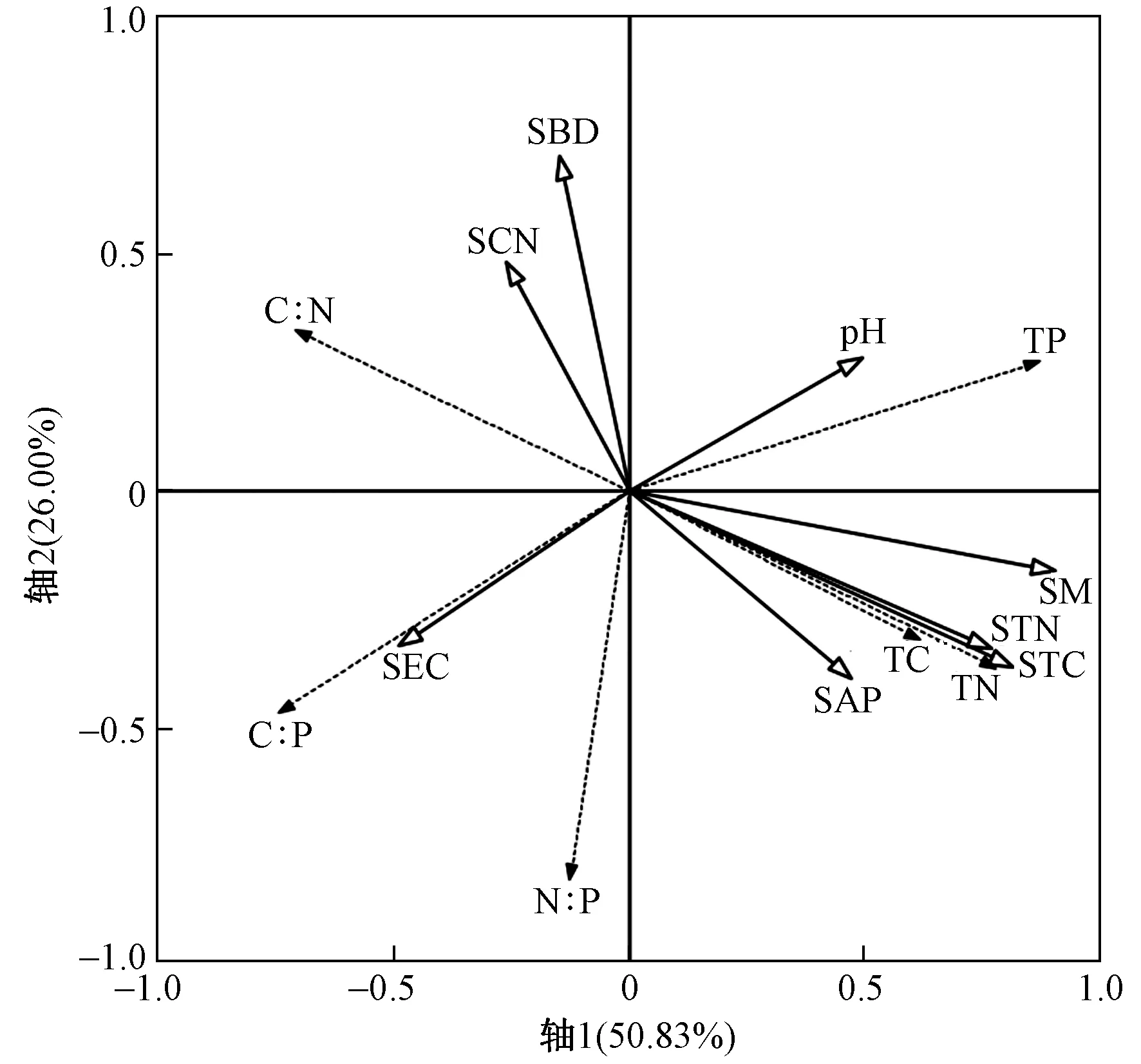
注: TC—群落TC含量;TN—群落TN含量;TP—群落TP含量;C∶N—群落C∶N;C∶P—群落C∶P;N∶P—群落N∶P;SCN—土壤C∶N;SBD—土壤容重;pH—土壤pH;SM—土壤含水量;STN—土壤TN含量;STC—土壤TC含量;SAP—土壤速效磷含量;SEC—土壤电导率.图3 多枝柽柳群落碳氮磷化学计量特征与土 壤因子RDA排序结果Fig.3 RDA ordination plot of the T. ramosissima community stoichiometric traits and soil properties
从RDA排序结果来看,前2个排序轴分别解释了多枝柽柳群落计量学特征变化的50.83%和26.00%(见图3). 排序轴1主要解释了土壤含水量(F=20.3,P<0.001)和土壤pH(F=3.7,P<0.05)对柽柳群落计量学特征变化的影响,排序轴2主要解释了土壤容重(F=9.0,P<0.001)对多枝柽柳群落计量学特征变化的影响(见图3). 该结果表明,对多枝柽柳群落计量学特征变化具有显著影响的土壤因子为土壤含水量、土壤容重和土壤pH. 偏冗余分析表明,土壤含水量、土壤容重和土壤pH共同解释了多枝柽柳群落计量学特征变化的57.7%,土壤含水量的单独解释率在总解释率中的占比(32.8%)最大,其次是土壤含水量和土壤pH的交互作用(12.1%)以及土壤容重的单独解释率(10.5%),而土壤pH的单独解释率相对较低(见图4).

图4 多枝柽柳群落碳氮磷化学计量 特征与土壤因子偏冗余分析结果Fig.4 Partial redundancy analysis of the T. ramosissima community stoichiometric traits and soil properties
3 讨论
3.1 群落水平上荒漠河岸多枝柽柳灌丛TC、TN、TP含量及其计量比
该研究聚焦于黑河下游极端干旱荒漠河岸地带(年降水量30~40 mm)沿河梯度上多枝柽柳群落的碳氮磷化学计量特征及其影响因素. 在群落水平上,该研究中多枝柽柳群落TC含量平均值为380.27 mgg,略高于黑河下游荒漠河岸地带和中下游戈壁荒漠地区物种水平的TC含量平均值(见表4),而多枝柽柳群落TC含量平均值显著低于黄土高原地区、全球尺度下物种水平的TC含量平均值[27-29]. 多枝柽柳群落TN含量平均值为30.42 mgg,高于黑河下游荒漠河岸地带和中下游戈壁荒漠地区物种水平的TN含量平均值(见表4),同时也高于黄土高原地区、全球尺度下物种水平的TN含量平均值[27-29]. 多枝柽柳群落TP含量平均值为1.54 mgg,高于黑河下游荒漠河岸地带物种水平的TP含量平均值(见表4),而略低于中下游戈壁荒漠地区、黄土高原地区、全球尺度下物种水平的TP含量平均值[27-29],这可能是导致该研究中N∶P较高的原因. 与全球尺度、区域尺度相比较,该研究中多枝柽柳群落具有TC含量低、TN含量高、N∶P高、TP含量相对稳定的特点. 此外,与黑河下游荒漠河岸地带和黑河中下游荒漠地区物种水平下的研究结果相比,黑河下游荒漠河岸地带群落水平碳氮磷化学计量参数变异系数相对较小(见表4),反映出多枝柽柳群落水平碳氮磷化学计量特征相对较高的内稳性.

表4 黑河中下游荒漠地区植物在物种和群落水平下碳氮磷化学计量特征
3.2 荒漠河岸沿河多枝柽柳群落碳氮磷化学计量变化特征
在沿河梯度上,多枝柽柳群落TC含量平均值相对较低,主要与该区域极端干旱和盐碱的环境有关,植物为应对干旱和盐碱胁迫,其自身代谢成本增加,光合速率受到抑制,从而使得多枝柽柳群落的固碳能力降低[31-32],这也可能是多枝柽柳群落TC含量随沿河距离的增加呈显著下降变化趋势的主要原因. 由于该地区河流水补给的地下水是植物和土壤的主要水分来源[24],随着沿河距离的增加,地下水埋深逐渐增加,土壤含水量逐渐降低,水分条件变差使得植物生产力下降,多枝柽柳群落TC含量降低[8,17]. 这和多枝柽柳群落TC含量与土壤含水量呈显著正相关的分析结果相符合,尤其是在土壤水分条件最好的样地T1,群落TC含量平均值为423.67 mgg,而在土壤水分条件最差的样地T9,群落TC含量平均值仅为348.96 mgg,这在一定程度上说明水分条件变化影响着荒漠植物群落碳含量的变化,较低的碳含量可能与极端干旱的环境有关[32]. 有研究表明,在自然条件下,植物叶片氮含量与土壤氮含量呈线性正相关[33],即使在半干旱-干旱地区的氮添加控制试验中,土壤无机氮含量的增加也会导致植物叶片氮含量的显著增加[34],这与该研究中多枝柽柳群落TN含量与土壤TN含量呈显著正相关的分析结果相符合. 该研究中群落相对较高的氮含量主要与优势种多枝柽柳和干旱盐碱生境相关. 在沿河梯度上,多枝柽柳是一种典型的内生固氮菌属灌木[35],此外,在盐碱环境下,荒漠植物可积累大量含氮物质,导致多枝柽柳群落具有相对较高的氮含量,相对较高的氮含量可能是荒漠植物对极端干旱和盐碱环境的适应结果[32,36].
磷元素被认为是中国陆地植物生长的主要限制性养分,植物磷含量低主要是由土壤磷含量较低引起的[37]. 该研究中多枝柽柳群落TP含量平均值(1.54 mgg)略高于全国陆地植物物种平均水平(1.46 mgg),且与植物吸收关系密切的土壤速效磷含量(4.48 mgkg)也高于全国平均水平(3.83 mgkg)[38],该研究中黑河下游荒漠河岸地带多枝柽柳群落TP含量可能是由于相对较高的土壤磷含量所致. 然而,该研究中多枝柽柳群落TP含量与土壤速效磷含量呈正相关(R=0.353),但不显著,这与该区域盐分胁迫有关. 有研究表明,在受盐胁迫土壤中,存在大量的Cl-、SO42-等阴离子,它们会与磷元素产生竞争效应,抑制植物对磷元素的吸收[39],这与该研究中多枝柽柳群落TP含量与土壤电导率呈显著负相关的分析结果相符合. 在沿河梯度上,在距河 1 800~2 800 m之间,土壤电导率为13.32~15.05 mScm,极端盐胁迫可能抑制植物对磷元素的吸收,这也可能是多枝柽柳群落TP含量呈先下降后上升的变化趋势的主要原因. 有研究表明,磷元素主要来源于土壤母质,干旱区降水过程对土壤的淋溶程度较低,相对于氮元素,土壤母质中磷含量相对丰富,使得氮元素更易成为限制性元素[40-41]. Güsewell等认为,在群落水平上N∶P 更能准确判断植物生长的养分限制[42],而笔者得到的柽柳群落N∶P平均值为20.04,相对较高的N∶P可能意味着该区域多枝柽柳群落在生长旺季氮过量而磷含量相对不足.
3.3 荒漠河岸沿河多枝柽柳群落碳氮磷化学计量格局形成的影响因素分析
在干旱区,尤其是极端干旱地区,水分条件是影响植物生长和分布的主要影响因子[17]. 该研究中多枝柽柳群落TC、TN、TP含量均与土壤含水量呈显著正相关,在黑河下游荒漠河岸地带,水分条件较好的近河地带,土壤水分和土壤养分含量相对较高,植物群落生长条件较好,植物群落通过水分-养分的耦合效应获取更多的养分[8,43];随着沿河垂直距离的增加,养分和水分条件变差,导致植物能获取的养分减少,使得植物TC、TN、TP含量显著降低[8]. 偏冗余分析结果表明,土壤含水量、土壤容重和土壤pH会对多枝柽柳群落碳氮磷化学计量特征产生显著影响,进一步说明多枝柽柳群落TC、TN、TP含量与水分条件有关. 水分条件较土壤pH的影响作用更为明显,可能是因为黑河下游极端干旱的环境导致水分条件更易成为荒漠植物生长的限制因素. 此外,土壤含水量和土壤pH的交互作用解释了多枝柽柳群落碳氮磷化学计量特征变化的12.1%,表明该区域土壤盐碱共同影响着多枝柽柳群落的生长. 考虑到植被与环境之间的复杂关系,在黑河下游荒漠河岸地带进行长期多尺度野外调查,或者控制试验可能更有利于进一步阐明多枝柽柳群落对土壤水盐和养分的响应.
4 结论
a) 黑河下游荒漠河岸地带多枝柽柳群落具有TC含量低、TN含量高、N∶P高、TP含量相对稳定的特点. 与黑河中下游地区荒漠植物物种水平相比,在群落水平上,荒漠河岸多枝柽柳灌丛碳氮磷化学计量特征的变异系数相对较小,内稳性较强. 群落水平N∶P 的分析表明,黑河下游荒漠河岸多枝柽柳群落在生长旺季受磷元素的限制程度较大.
b) 在沿河梯度上,多枝柽柳群落TC、TN、TP的含量以及C∶N、C∶P和N∶P在不同沿河距离样地内均具有显著差异(P<0.05). 随着沿河距离的增加,多枝柽柳群落TC、TN、TP含量和C∶N∶P均呈现出显著的变化趋势. 土壤含水量、土壤容重和土壤pH较好地解释了多枝柽柳群落碳氮磷化学计量特征的变化,共同解释了总变异的57.7%.
c) 在解释多枝柽柳群落碳氮磷化学计量特征的变化方面,土壤含水量以及土壤含水量和土壤pH交互作用的贡献率大于土壤pH,表明水分是该区域植物生长的主要限制因子,植物可能通过调节自身营养元素的比例来适应极端干旱盐碱的环境.
