光照对纤细裸藻生长及光合色素含量影响的研究
2021-03-24王祎哲姚奕炯王梓懿贾旭颖高金伟张文慧周文礼
王祎哲,姚奕炯,王梓懿,贾旭颖,高金伟,张文慧,邵 蓬,窦 勇,周文礼
(天津农学院 水产学院,天津市水产生态及养殖重点实验室,天津 300384)
纤细裸藻(Euglenagracilis),表质软,形状易变,直径20~100 μm,主要分布于淡水之中,是一种单细胞光合作用的裸藻[1],可以在低pH、高重金属等不良环境条件下生存,同时纤细裸藻具备植物和动物的特性,既可以在光照条件下进行光合自养,也可以在黑暗条件下通过补充碳源进行异养来维持生存[2]。纤细裸藻含有维生素、矿物质、不饱和脂肪酸、氨基酸等诸多营养元素[3],其中氨基酸种类达20种,包括亮氨酸、赖氨酸和色氨酸等9种人体所必需的氨基酸[4],其无细胞壁,所含的微量营养物质更易被人体消化和吸收[5],此外纤细裸藻作为水生生态系统中常见的初级生产者,可以产生大量的有益代谢物[6],其光合作用下特有的产物副淀粉(β-1,3-葡聚糖),结构类似于淀粉,具有抗感染[7]、抗病毒[8]、抗肿瘤[9]等多种医学价值,因此被广泛应用于食品、动物饲料、保健品和化妆品等领域[10-12]。在水产养殖中,纤细裸藻不仅可以作为水产动物的饵料,还具有良好的水质净化效果,与其他藻类相比,在废水中生长的裸藻具有更快的生长速率,同时带走水体中大量的碳、氮和磷[13],在缺氧和酸性条件下对水体中镉也具有良好的去除效果[14]。
藻类的生长状况与环境因子有很大关联,如温度、盐度、光照、pH均为影响藻类生长的关键环境因子[15],尤其在大规模培养过程中,无法保证藻体对光能的持续、高效利用,极易导致培养效率低、产品质量不稳定等问题。因此,优化光照条件、提高光能利用率是纤细裸藻产业化培养亟待解决的主要问题之一[16]。而光照作为复杂的一类环境因子,对微藻生长发育起主要影响的因素包括光照度、光照周期和光质[17-21]。已有研究证明,不同光照条件影响北方娄氏藻(Lauderiaborealis)的生长和光合色素含量,光照度2500~3500 lx时适宜北方娄氏藻生长[17];温度25 ℃、光照度6000 lx时适宜针叶蕨藻(Caulerpasertularioides)的生长[18];光照周期为16L∶8D时,对雨生红球藻(Haematococcuspluvialis)生长具有显著的促进作用[19]。蓝光促进浒苔(Ulvaprolifera)生长的同时也有利于其光合作用的进行,而绿光对其生长和光合作用均产生了明显的抑制作用[20];蓝光下蛋白核小球藻(Chlorellapyrenoidosa)不仅生长效果最佳,且促进了叶绿素b的合成[21]。这些试验结果表明,光是影响藻类生长的重要环境因子之一。但是目前关于不同光照条件对纤细裸藻生长及光合色素含量影响的研究报道较少。笔者以纤细裸藻为研究对象,探查光照度、光照周期和光质对其生长及光合色素含量的影响,以期获得纤细裸藻生长、光合色素积累的最佳光照条件,为探索纤细裸藻的高密度培养方式提供科学依据。
1 材料与方法
1.1 微藻及培养
试验用纤细裸藻来源于天津市水产生态及养殖重点实验室。基础培养基为蒸馏水配制的经120 ℃条件下灭菌20 min的AF-6培养基。取对数生长期生长良好的纤细裸藻按照V(藻液)∶V(培养基)=1∶1的比例在超净工作台内接种,在恒定培养箱内进行静置培养,设定培养箱温度为25 ℃,光照度为3000 lx,光照周期为12L∶12D,每日定时摇动至少3次,防止藻液贴壁或沉淀。
1.2 试验方法
1.2.1 光照度对纤细裸藻生长及色素含量影响
将150 mL密度为5×105个/mL的纤细裸藻藻液接种于150 mL的AF-6培养基中于500 mL锥形瓶中备用,利用培养箱设定不同光照度:0、1500、3000、4500 lx和6000 lx,光照周期为12L∶12D,光质为白光。于光照培养箱(AL-36美国珀尔瓦西)中静置培养,试验周期为10 d,每组设3个平行。试验期间每2 d定时取样10 mL,分别测定纤细裸藻细胞密度以及光合色素含量。
1.2.2 光照周期对纤细裸藻生长及色素含量影响
将150 mL密度为5×105个/mL的纤细裸藻藻液接种于150 mL的AF-6培养基中于500 mL锥形瓶中备用,利用培养箱设定不同光照周期:16L∶8D、14L∶10D、12L∶12D、10L∶14D和8L∶16D,光照度为3000 lx,光质为白光。于光照培养箱(AL-36美国珀尔瓦西)中静置培养,试验周期与测量指标同1.2.1。
1.2.3 光质对纤细裸藻生长及色素含量影响
将150 mL密度为5×105个/mL的纤细裸藻藻液接种于150 mL的AF-6培养基中于500 mL锥形瓶中备用,以5种颜色LED灯带作为光源,固定于培养箱内侧,设置不同光质:绿光(495~530 nm)、蓝光(450~480 nm)、红光(615~650 nm)、白光(450~465 nm)和黄光(580~595 nm),光照度为3000 lx,光照周期为12L∶12D。于光照培养箱(AL-36美国珀尔瓦西)中静置培养,试验周期与测量指标同1.2.1。
1.3 测定方法
1.3.1 生物量测定
藻细胞密度测定通过血球计数板计数获得。
藻细胞增长速率(R)按下式计算:
R/个·d-1=(lnn2-lnn1)/(t2-t1)
式中,t1为起始培养时间(d),t2为结束培养时间(d),n1为特定培养时间t1的细胞密度(个/mL),n2为特定培养时间t2的细胞密度(个/mL)。
1.3.2 色素含量测定
参考乙醇法[22]对藻类进行叶绿素a、叶绿素b、类胡萝卜素质量浓度测定。
取样品5 mL经5000 r/min离心10 min去掉上清液,加入5 mL 95%乙醇溶液后于4 ℃冷藏24 h后,经5000 r/min离心10 min,取上清液,测定各波长下吸光度。
ρa=13.95D665-6.88D649
ρb=24.96D649-7.32D665
ρc=(1000D470-2.05ρa-114.8ρb)/245
式中,ρa、ρb分别为叶绿素a和b的质量浓度(mg/L),ρc为类胡萝卜素的总质量浓度(mg/L);D665、D649和D470分别为叶绿体色素提取液在波长665 nm、649 nm和470 nm下的吸光度。
1.4 数据处理与统计分析
所有结果用平均值±标准差表示,使用SPSS 19.0进行单因素方差分析和Duncan多重比较分析,并用Origin 8.0软件作图。
2 结果与分析
2.1 光照度对纤细裸藻生长及色素含量影响
光照度对纤细裸藻细胞密度和增长速率的影响见图1、表1,光照度对纤细裸藻生长有显著影响(P<0.05)。

表1 光照度对纤细裸藻的增长速率影响Tab.1 Effect of light illuminance on growth rate of alga E.gracilis

图1 光照度对纤细裸藻细胞密度的影响Fig.1 Effect of light illuminance on cell density of alga E.gracilis不同小写字母表示同一时间各处理组间差异性显著(P<0.05),下同.Means with different letters are significant differences between treatment groups at the same time (P<0.05),et sequentia.
试验期间,随着培养时间的增加,各组细胞密度均呈上升趋势。其中3000 lx组藻细胞生长情况最好,始终呈持续稳定的生长趋势,尤其在试验中后期(4~10 d)其细胞密度均高于其他处理组,试验结束时,细胞密度达到最高,为5.18×105个/mL,增长速率达到0.162个/d。其次为4500、1500 lx组,试验结束时两处理组细胞密度分别为4.84×105个/mL和4.58×105个/mL,增长速率达到了0.156个/d和0.154个/d。而高光照度6000 lx组为3.81×105个/mL,增长速率为0.133个/d,显著低于以上各组(P<0.05)。试验期间,0 lx组的细胞密度始终显著低于其他组(P<0.05),至试验结束,细胞密度仅达到初始值的1.5倍。
光照度对纤细裸藻光合色素质量浓度的影响见图2,光照度对纤细裸藻叶绿素a、b和类胡萝卜素质量浓度均有显著影响(P<0.05)。

图2 光照度对纤细裸藻光合色素质量浓度的影响Fig.2 Effect of light illuminance on photosynthetic pigment contents in alga E.gracilisa.叶绿素a;b.叶绿素b;c.类胡萝卜素.a.chlorophyll a;b.chlorophyll b;c.carotenoid.
试验期间,0 lx组各色素质量浓度始终显著低于其他组(P<0.05);3000 lx组叶绿素a质量浓度始终高于其他组。光照组(1500~6000 lx)叶绿素a质量浓度呈明显升高趋势,至第10天,各组叶绿素a质量浓度均有所下降,但3000 lx组下降幅度较小,试验结束时其叶绿素a质量浓度显著高于其他组(P<0.05)。试验期间,除4500 lx组叶绿素b在第10天略有降低外,其余各组叶绿素b质量浓度均随培养时间的延长而升高。第10天,3000 lx组叶绿素b质量浓度最高,显著高于4500、6000 lx组(P<0.05)。试验期间,除0 lx黑暗组以外,其他光照组类胡萝卜素质量浓度均在第8天达到最高值,随后略有下降,第8天1500 lx组类胡萝卜素质量浓度最高,但与其他组无显著差异,而在第10天则是3000 lx组类胡萝卜素质量浓度最高,显著高于其他组(P<0.05)。
2.2 光照周期对纤细裸藻生长及色素质量浓度影响
光照周期对纤细裸藻细胞密度和增长速率的影响见图3、表2,光照周期对纤细裸藻生长有显著影响(P<0.05)。

图3 光照周期对纤细裸藻细胞密度的影响Fig.3 Effect of photoperiod on cell density of alga E.gracilis
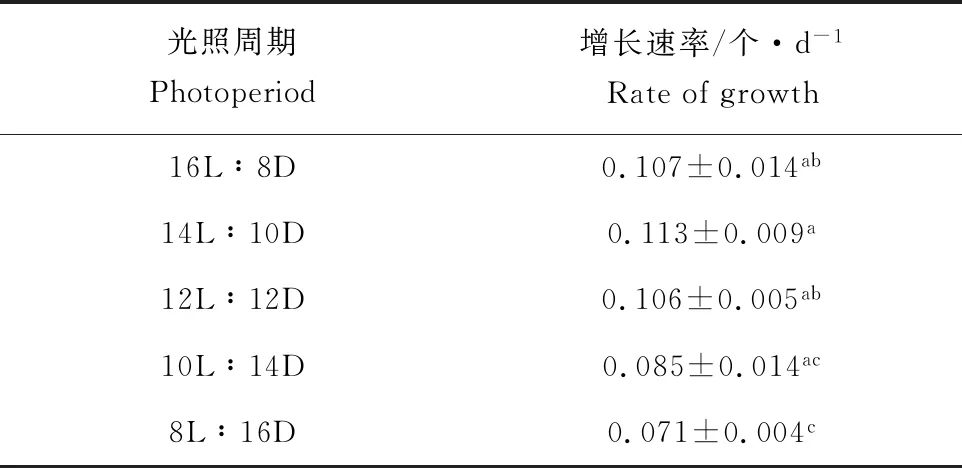
表2 光照周期对纤细裸藻增长速率的影响Tab.2 Effect of photoperiod on growth rate of alga E.gracilis
试验期间,纤细裸藻细胞密度呈增长趋势,各组前2 d并无显著性差异(P>0.05),第4天起至培养结束,长光照周期组(12L~16L)细胞密度均高于短光照周期组(8L~10L),并产生显著差异(P<0.05)。其中,14L∶10D组细胞密度始终高于其他组,至第10天,细胞密度达到最高,为4.57×105个/mL,增长速率达到0.113个/d,其次为12L∶12D和16L∶8D组,细胞密度分别为4.29×105个/mL和4.26×105个/mL,而8L∶16D组藻细胞生长缓慢,试验结束时细胞密度仅为起始密度的2.03倍,显著低于其他组(P<0.05)。
光照周期对纤细裸藻光合色素质量浓度的影响见图4,光照周期对纤细裸藻叶绿素a、b和类胡萝卜素质量浓度均有显著影响(P<0.05)。

图4 光照周期对纤细裸藻光合色素质量浓度的影响Fig.4 Effect of photoperiod on photosynthetic pigment contents of alga E.gracilisa.叶绿素a;b.叶绿素b;c.类胡萝卜素.a.chlorophyll a;b.chlorophyll b;c.carotenoid.
试验期间,纤细裸藻的叶绿素a整体呈增长趋势,第10天,14L∶10D组叶绿素a质量浓度达到最高,其次为12L∶12D、16L∶8D组,8L∶16D组整体增长缓慢,并在8 d后出现下降,叶绿素a的积累受到抑制;其余组叶绿素a质量浓度始终保持上升趋势。试验第4天时,14L∶10D、16L∶8D组显著高于其他组(P<0.05),直至第10天,14L∶10D组的叶绿素b质量浓度达到最高,8L∶16D组的叶绿素b质量浓度增长缓慢,显著低于其他处理组(P<0.05)。试验期间,14L∶10D组类胡萝卜素质量浓度保持持续增长,并在第10天时达到最大,其次为16L∶8D组,二者与其他处理组产生显著差异(P<0.05),8L∶16D组类胡萝卜素质量浓度始终处于缓慢增长,显著低于其他处理组(P<0.05)。
2.3 光质对纤细裸藻生长及色素质量浓度影响
光质对纤细裸藻细胞密度影响见图5、表3,光质对纤细裸藻生长有显著影响(P<0.05)。

表3 光质对纤细裸藻的增长速率影响Tab.3 Effect of light wavelength on growth rate of alga E.gracilis
试验前2 d,各试验组无显著差异(P>0.05);第4天起,白光组与蓝光组细胞密度显著高于其他处理组(P<0.05),第10天时,蓝光组细胞密度达到最高,为3.26×105个/mL,增长速率为0.079个/d,其次为白光组,细胞密度达到3.17×105个/mL,增长速率为0.077个/d,二者显著高于其他组(P<0.05),各组细胞密度大小依次为蓝光>白光>红光>绿光>黄光,黄光组显著低于其他组,不适宜纤细裸藻生长。

光质对纤细裸藻光合色素质量浓度的影响见图6,光质对纤细裸藻对纤细裸藻叶绿素a、b和类胡萝卜素质量浓度均有显著影响(P<0.05)。

图6 光质对纤细裸藻光合色素质量浓度的影响Fig.6 Effect of light wavelength on photosynthetic pigment contents of alga E.gracilisa.叶绿素a;b.叶绿素b;c.类胡萝卜素.a.chlorophyll a;b.chlorophyll b;c.carotenoid.
试验前4 d,各处理组间叶绿素a质量浓度无显著差异(P>0.05),第6天时,蓝光组叶绿素a质量浓度显著高于其他组(P<0.05),至试验结束,蓝光组与白光组叶绿素a质量浓度显著高于其他组(P<0.05)。试验期间,各处理组叶绿素b质量浓度前4 d无显著差异(P>0.05),第10天,蓝光组叶绿素b质量浓度达到最大,显著高于其他组(P<0.05)。试验第4天起,蓝光组与白光组类胡萝卜素质量浓度呈现良好的增长趋势,至试验结束,蓝光组类胡萝卜素质量浓度显著高于其他组(P<0.05)。
3 讨 论
3.1 光照度对纤细裸藻生长及色素含量的影响
光照是藻类生长发育的基本能量来源,直接影响藻类的生长、发育、代谢等生命活动[23],同时也作为直接作用因子影响藻类的色素形成以及光合效率[24-25]。藻类细胞体内所含的色素种类可依据在光合作用过程中的作用分为捕光色素和光保护色素,其中以叶绿素a、叶绿素b为代表的捕光色素可将吸收的光能有效地传递到相关的反应中心转变为化学能,而类胡萝卜素作为光保护色素将剩余能量吸收,避免膜体受伤,从而达到光保护作用[26-27]。因此,可通过光照条件下光合作用产生的光合色素含量的多少来判断藻类的生理状态[28]。
众多研究表明,藻类的生长速率随着光照度的增强而增加(从0至某一最适光照度),而当光照度超过藻类适宜范围时,藻细胞生长速率减弱甚至被抑制[29-30]。研究发现,凯氏小球藻(C.kessleri)在9600 lx光照度下细胞密度高于其他光照度组[0、1200、2400、4800、1600 lx],而光照度增至16 000 lx时,则出现光抑制现象,说明凯氏小球藻在低光强下处于光限制阶段,生物量随着光照度的增加而增加,再进一步增加光照度超过“光饱和点”时则出现光抑制现象[31]。本试验结果表明,光照度为0 lx时,细胞密度及生长速率均显著低于其他处理组,此时藻细胞的光合效率降低,进而影响了微藻的生物量,随着光照度的增加,至试验结束,3000 lx组细胞密度达到最高,其次为4500 lx组,可见,适当高光处理可以增加藻细胞对光的耐受能力[32],而当光照度超过“光饱和点”达到6000 lx时,纤细裸藻的生长反而受到抑制,可见,光照度适宜时,细胞利用光能合成有机物供养自己,一旦超过“光饱和点”,藻细胞对此表现出一定的生理抗性,相关光合色素结构受到刺激被破坏,抑制作用增强,从而影响藻类的生长[33-34]。
研究表明,光照度的不同会影响藻类色素的组成[35],北方娄氏藻[36]叶绿素a含量在低光照度下最高,光照度过高时,会引起光抑制现象,藻体内的色素体会被光解并褪色成黄白色,严重影响藻类生长及色素合成。对铜绿微囊藻(Microcystisaeruginosa)[37]的研究发现,光照度达到3200 lx时,其叶绿素荧光参数达到稳定状态,而当光照度升至25 600 lx时,其各荧光参数不再稳定,变化较大,表明此时铜绿微囊藻光合系统受到了影响[37]。本试验中,纤细裸藻色素含量在低、中光照度时(0、1500、3000 lx)随光照度的增加而增加,在高光照度时(4500、6000 lx)随光照度的增加而减少。3000 lx组叶绿素a含量在第8天达到峰值,达到光合作用的饱和光照度,之后随着光照度的继续増强,光合作用受到光抑制,叶绿素a含量减少。推测可能是由于纤细裸藻在较低光照度条件下依旧能合成细胞色素,但合成效率较低,含量较少,随光照度增加,细胞合成色素的能力增强,含量增多,对藻类生长起到促进作用;而当光照度过高时,细胞色素受光氧化破坏,色素含量降低进而影响藻类生长[38]。
3.2 光照周期对纤细裸藻生长及色素含量的影响
藻类的生长不仅与光照度有关,与光照周期也密切相关,光合作用分为光反应和暗反应两个阶段,只有确保藻细胞生长在适宜的光暗交替条件下才能使光合作用顺利进行,藻细胞得以良好生长[39]。研究表明,铜绿微囊藻[40]在光照周期为9L∶15D时的比增长率和最大细胞数均达到最大,当光照周期为12L∶12D,光照度为5000 lx时小球藻(Chlorellasp.)、湛江等鞭金藻(GY-H2Isochrysiszhangjiangensis)、青岛大扁藻(Platymonashelgolandicatsingtaoensis)和绿色杜氏藻(Dunaliellaviridis)的生长速率均达到最大[39],可见,并非一味延长光照时间更有利于藻类的生长,不同藻类对光照周期有不同的需求,光反应与暗反应时间达到良好的匹配,才能利于光合产物的合成[41],进而影响藻类的细胞密度。本试验结果表明,纤细裸藻在光照周期为8L∶16D时细胞密度显著低于其他组,可见光照时间过短,光合作用受到抑制,藻细胞停止制造有机物,对细胞密度产生影响。光照周期为14L∶10D时细胞密度显著高于其他处理组,且生长速率达到最大,可见延长光照2 h,采用光照周期为14L∶10D时更适合纤细裸藻的生物量积累。
研究表明,小球藻和斜生栅藻(Scenedesmusobliquus)在18L∶6D光照周期下,两种藻在单位水体内的叶绿素含量达到最高,而在0L∶24D条件下的叶绿素最低[42]。叶绿体中含有光合中间产物,其含量若超过一定水平,叶绿素合成受阻,从而引起叶绿素含量显著降低。所以适当地延长光照时间有利于叶绿素的合成,但对于叶绿素合成而言,存在一个最佳的光照时间,超过一定范围,光照时间增长反而抑制叶绿素的合成。由本试验得出结论,细胞色素含量在光照时间为14 h时达到最高,而在16 h时反而降低,此时光的透过率降低,光合作用减弱,培养液中的营养物质不足以满足藻细胞新陈代谢的需要,进而停止生长,叶绿素的合成减少,含量下降。说明纤细裸藻对色素合成最适的光照周期为14L∶10D,光照时间过长,叶绿素合成受阻,导致叶绿素含量降低。
3.3 光质对纤细裸藻生长及色素含量的影响
微藻的生长对光质的吸收具有选择性,光质通过影响藻体内光敏色素的感知性进而影响藻类的生长发育、光合作用和生理代谢[43]。研究表明,蓝光对于展枝马尾藻(Sargassumpatens)等大型藻类有显著促进生长的作用[44]。坛紫菜(Pyropiahaitanensis)在蓝光处理下自由丝状体的生长速率最快,而红光处理下生长速率最慢[45]。本试验结果表明,纤细裸藻在蓝光及白光下细胞密度显著高于其他组,其中蓝光效果更为显著,此种蓝光效应可以解释为颜色适应,它可以通过影响蛋白质的合成和酶的活性来帮助细胞在不同条件下生长,然后使细胞发生复杂的生理变化,分析原因可能与叶绿素在PSⅡ中的作用有关,该作用导致蓝光条件下叶绿素含量增加,从而保护叶绿素不被蓝光吸收和过滤而发生光氧化现象[46]。可见,纤细裸藻对不同光波长的反应存在差异,蓝光可以作为纤细裸藻生长的高效光质。
不同波长的光源不仅会直接影响藻类生长,也会影响其光合色素含量进而影响藻类光合作用效率[47]。对中华盒形藻(Biddulpphiasinensis)研究发现,蓝光显著促进叶绿素a的合成和积累[48]。蓝光对江蓠(Gracilaria)、坛紫菜和角叉菜(Chondrusocellatus)等大型海藻中叶绿素a合成有明显的促进作用[49-51],且对坛紫菜的类胡萝卜素合成也具有显著促进作用[52]。本试验结果与以上结果相似,蓝光条件下纤细裸藻叶绿素a和类胡萝卜素含量明显高于其他处理组,可见蓝光更有利于叶绿素a及类胡萝卜素的积累,提高了藻体的光合反应速率。而黄光与红光下叶绿素合成最少,这与Howell等[34]研究结果——螺旋藻在红光下培养时,其叶绿素含量最高,而在蓝光和绿光下培养,其叶绿素a的含量最低这一结果相反。产生这种差异可能是由于藻类光合色素系统可以吸收可见光内波段的光谱成分不同导致,螺旋藻属蓝藻门,蓝藻的藻胆素对可见光的吸收主要在红光区,因而光合效率较高,产生的光合产物多。
综上所述,光照对纤细裸藻生长及光合色素含量影响较大,光照度为3000 lx时利于纤细裸藻生长及光合色素积累,光照周期为14L∶10D时利于纤细裸藻生长及光合色素积累,蓝光利于纤细裸藻生长以及光合色素积累。下一阶段研究还有待于在单因素试验的基础上采取正交试验设计寻求一种适宜纤细裸藻生长及色素积累的复合光照条件,以期为纤细裸藻产业的发展提供理论依据及研究基础。
