樟叶越桔嫩枝内生真菌的植物病原菌拮抗活性
2021-03-23曾为林陈肖学王连春刘惠民
严 冬,曾为林,陈肖学,刘 云,王连春,刘惠民,赵 平,c
(西南林业大学 a.西南地区林业生物质资源高效利用国家林业和草原局重点实验室;b.西南山地森林资源保育与利用教育部重点实验室;c.云南森林资源培育与利用协同创新中心,云南 昆明 650233)
植物内生真菌是指生活于健康植物的各种组织和器官内部的真菌[1-2],普遍存在于植物根、茎、叶等不同部位的器官和组织细胞中,因宿主植物的种类及其生长环境等因素的差异,导致内生真菌种类繁多,具有极为丰富的物种多样性,并与宿主植物长期的生态系统演化进程中形成互惠共生关系,对植物生长发育、抵御胁迫等具有重要意义[3-5]。此外,研究还发现丛枝菌根(Arbuscular mycorrhiza,AM)真菌能够与植物根系形成有益共生体,“AM 真菌-根系-土壤-植物”之间形成的耦合作用可以调控植物生长及植被恢复与演替[6]。在已发现的内生真菌中大约有1/3 的真菌对植物病原真菌显示出较强的抑制活性,使得植物内生真菌成为生物防治中极具潜力的微生物农药、增产菌或生防载体菌的重要来源,具有重要的开发应用前景[7]。樟叶越桔Vaccinium dunalianum为杜鹃花科Ericaceae 越桔属Vaccinium多年生常绿灌木,主要分布在云南滇中高原至滇南和滇东南地区[8-9]。该植物全株均可药用,具有祛风除湿、舒筋活络等功效,其幼嫩叶芽经采摘、蒸熟、搓揉、阴干加工后在云南彝族民间又称“雀嘴茶”,据记载自明代开始就作为一种茶代用品饮用至今,是一种独具特色的民族药食兼用植物[10]。Zhao等[11]首次发现该植物富含具有黑色素生成抑制活性优于熊果苷的咖啡酰熊果苷类物质,可望作为天然美白剂的替代资源加以利用。目前樟叶越桔仍处于野生状态,多生于贫瘠的山坡灌丛或石灰山灌丛,随着雀嘴茶的关注度不断提高,致使樟叶越桔遭受破坏性采摘,其野生资源数量急剧减少。开展其内生真菌生物学功能等相关研究,有利于了解内生真菌的生物学功能及其与宿主植物之间的相关关系,为樟叶越桔的种质资源保护及其可持续利用提供科学依据。
有关越桔属植物的研究多集中于种质资源利用和果实品质方面[12-14],对其内生真菌的研究较少,且仅涉及菌根真菌的分离及其多样性研究[15-17]。如Richardson 等[18]发现来源于狭叶越桔V.angustifoliumAiton 嫩枝的炭角菌属Xylariasp.内生真菌具有生产抗真菌剂灰黄霉素(griseofulvin)的能力,樊苗苗等[19]和陈肖学等[20]发现樟叶越桔叶内生真菌烟曲霉Aspergillus fumigatus等10 株菌株对小麦雪腐病菌Typhla incarnata、三七丝核病菌Verticillium cinnabarium、灰葡萄孢Botrytis cinerea、腐皮镰孢Fusarium solani和芸苔链格孢Alternaria brassicicola等植物病原真菌具有较强的抑制活性。植物内生真菌在宿主植物中的分布呈现多样性的特点,同一植物不同组织部位中内生真菌的数量和种类也有较大差异[21],樟叶越桔叶和嫩枝中的内生真菌也不例外,但嫩枝中的内生真菌是否具有较强的植物病原菌拮抗活性尚未报道。本研究以禾谷镰孢Fusarium graminearum等5 种病原真菌和密粘褶菌Gloeophyllum trabeum等3 种木腐菌为指示菌,采用平板对峙法测定樟叶越桔嫩枝内生真菌对指示病原菌的拮抗活性,以期为樟叶越桔病害生防菌剂的进一步开发利用提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株
内生真菌:分离自云南省武定县樟叶越桔的健康嫩枝,经形态鉴别和分子鉴定后保存于西南林业大学西南山地森林资源保育与利用教育部重点实验室,菌株编号及其归属见表1。
病原真菌:3 种木腐菌购自中国林业微生物保藏管理中心,5 种植物病原真菌购自中科院微生物研究所菌种保藏中心,均保藏于西南林业大学西南山地森林资源保育与利用教育部重点实验室,菌株编号及其归属见表2。

表1 樟叶越桔嫩枝20 株内生真菌菌株归属Table 1 Strain attribution of 20 endophytic fungi from the twigs of V.dunalianum

表2 供试病原真菌信息Table 2 Information for the tested pathogenic fungi
1.1.2 培养基
PDA 培养基:蒸馏水1 000 mL、马铃薯200 g、蔗糖10 g、琼脂10 g,用于樟叶越桔嫩枝内生真菌的活化及培养。
1.2 试验方法
1.2.1 菌株培养
在超净工作台中将已灭菌(121℃,22 min)的PDA 培养基倒平板,每平板约20 mL 的培养基,待冷却凝固后用接种针挑取冷冻管里面保存的菌丝体接种于平板正中心位置,于28℃恒温培养箱中培养7 d 备用。
1.2.2 拮抗活性测试
采用五点对峙法[19,22],测试表1中所示的内生真菌对供试病原菌的拮抗作用。用打孔器分别在活化后的二代内生真菌和病原真菌的菌落边缘取带有菌丝的菌饼。将病原真菌的菌饼接入新的PDA 平板正中央,同时以病原菌为中心的十字架距离病原真菌饼的2.5 cm 处,4 个点上分别接入直径0.5 cm 的植物内生真菌菌饼。以只在平板中央接入的病原真菌作为空白对照组,每个试验重复3 次,28℃恒温培养,每隔24 h 观察1 次,从对峙培养的第3 d 起用十字交叉法测量抑菌带距离和菌落的直径,空白组长满时停止测量。
1.2.3 抑菌率计算
内生真菌拮抗供试病原真菌的抑菌率按如下公式进行计算。
抑菌率=(对照菌落直径-处理组菌落直径)/对照菌落直径×100%。
2 结果与分析
2.1 内生真菌对3 种木腐菌的拮抗作用
不同内生真菌对3 种木腐菌的拮抗效果差别较大,见表3。根据对病原菌的抑制率≥75%为强拮抗能力,50%≤抑制率<75%为较强拮抗能力,抑制率<50%为弱拮抗能力[19,20,22,23],20 株内生真菌中除季也蒙毕赤酵母(VDBF-18)对采绒革盖菌呈现弱拮抗能力外,其它内生真菌菌株的抑制率均>50%,表明樟叶越桔嫩枝内生真菌对采绒革盖菌、绵腐卧孔菌和密粘褶菌均显示出较强的拮抗作用。其中,菌株VDBF-13 和VDBF-25 对3 株木腐菌的抑菌率均大于70%,VDBF-25 对采绒革盖菌、绵腐卧孔菌和密粘褶菌的拮抗效果如图1a—c 所示,其中VDBF-25 的生长速度比采绒革盖菌快,能与病原真菌产生空间或营养竟争,有效地利用PDA 培养基的营养物质生长,占领病原菌的入侵位点而使病原菌不能正常的生长和繁殖。对绵腐卧孔菌呈现强拮抗作用的内生真菌共有7 株,其中拮抗最强的是VDBF-18(图1d),抑菌率高达88.9%,其次是VDBF-25、VDBF-13、VDBF-16、VDBF-23、VDBF-11 和VDBF-37,抑菌率分别为87.5%、84.5%、80.9%、78.4%、76.9%和75.3%。对密粘褶菌拮抗作用最强的是VDBF-6(图1e),其次是VDBF-27,抑菌率分别为81.1%和76.2%。对采绒革盖菌拮抗作用最强的是VDBF-2(图1f),其次是VDBF-25,抑菌率分别为77.0%和76.3%。

表3 内生真菌对3 种木腐菌的拮抗效果Table 3 Antagonistic effects of endophytic fungi on three wood-decaying fungi
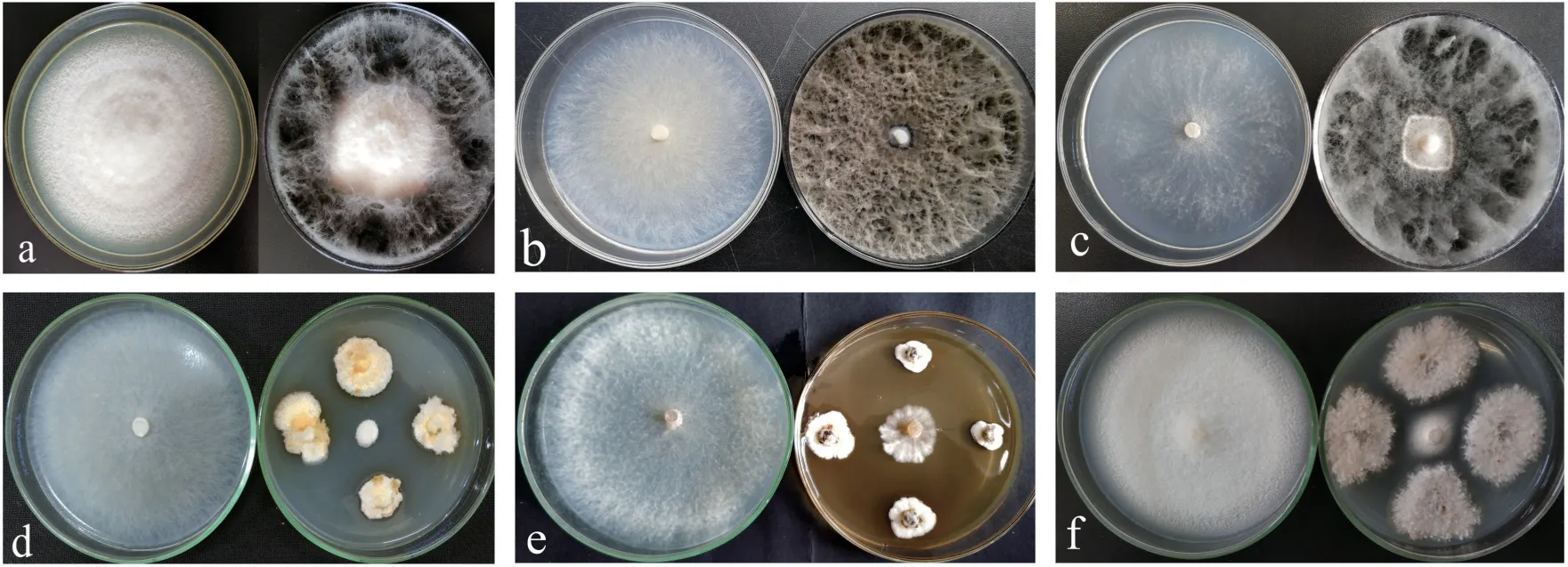
图1 部分内生真菌对3 种木腐菌的拮抗效果Fig.1 Antagonistic effects of some endophytic fungi on three wood-decaying fungi
2.2 内生真菌对5 种病原真菌的拮抗作用
从表4可以看出,20 株内生真菌中,对腐皮镰孢、禾谷镰孢、灰葡萄孢、尖孢镰孢和芸苔链格孢具有较强抑菌作用的活性菌株各有14、20、16、19 和19 株,分别占内生真菌总数的70%、100%、80%、95%和95%,表明樟叶越桔嫩枝内生真菌对5 种病原真菌具有广泛且较强的拮抗作用。菌株VDBF-6、VDBF-14、VDBF-21、VDBF-24、VDBF-25 和VDBF-27 除外,其余14株内生真菌对5 种病原真菌均呈现较强及以上拮抗能力,其中VDBF-2 对芸苔链格孢,VDBF-11和VDBF-42 对禾谷镰孢,VDBF-18 对灰葡萄孢和VDBF-37 对腐皮镰孢呈现强拮抗能力,抑菌率分别为81.0%、76.7%、77.8%、82.8%和75.2%。VDBF-6 菌株虽然对腐皮镰孢和尖孢镰孢显示无拮抗作用,对芸苔链格孢显示弱拮抗能力,但是其对禾谷镰孢和灰葡萄孢的拮抗作用最强,抑菌率分别高达90.8%和83.3%。VDBF-14 对腐皮镰孢和灰葡萄孢显示弱拮抗作用,但其对尖孢镰孢也呈现强拮抗作用,抑菌率为75.3%。VDBF-6 对灰葡萄孢(图2a),VDBF-2 分别对腐皮镰孢(图2b)、禾谷镰孢(图2c)、灰葡萄孢(图2d)、尖孢镰孢(图2e)和芸苔链格孢(图2f)的拮抗效果如图所示。
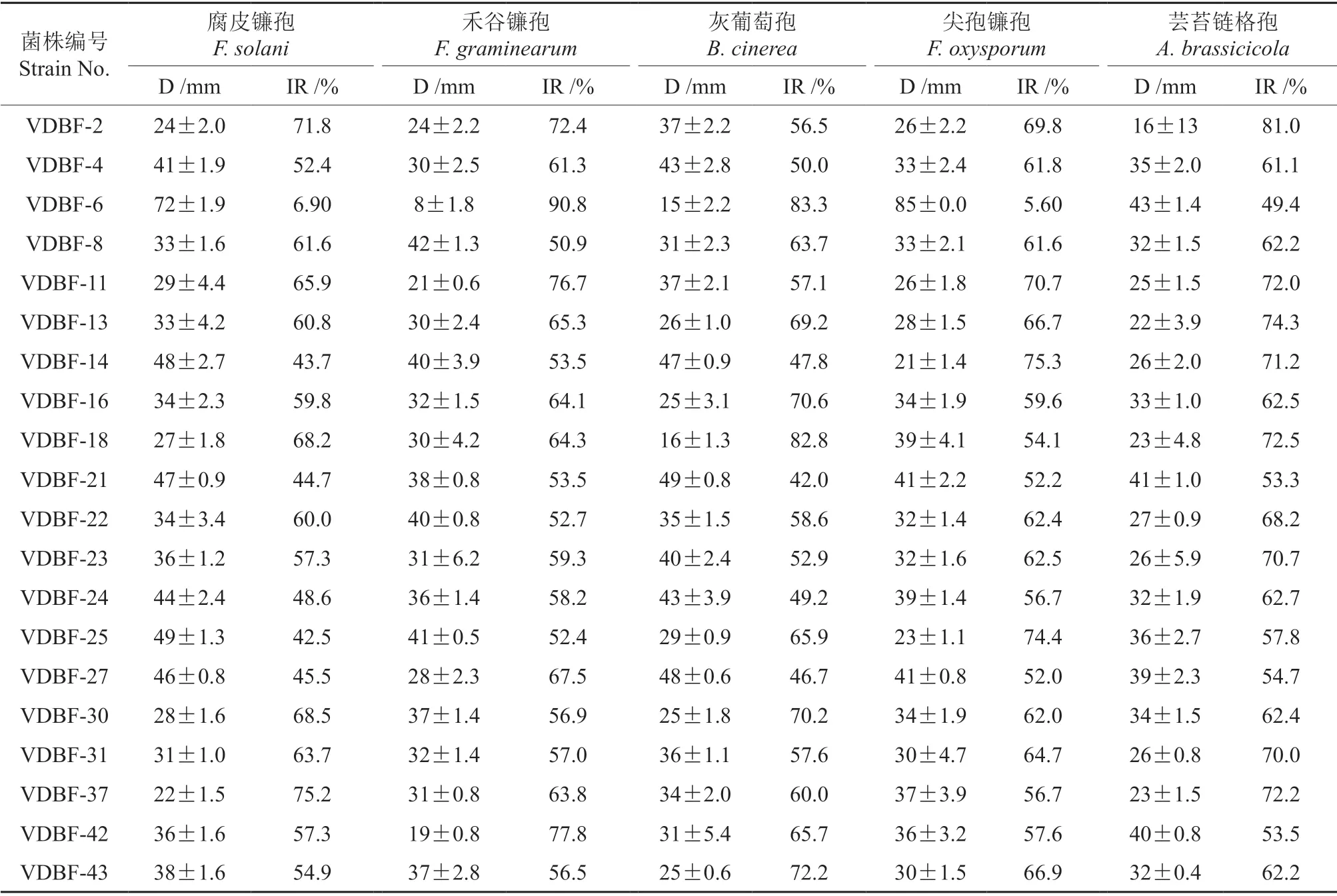
表4 内生真菌对5 种病原真菌的拮抗效果Table 4 Antagonistic effects of endophytic fungi on five pathogenic fungi
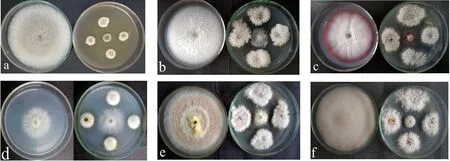
图2 部分内生真菌对5 种病原真菌的拮抗效果Fig.2 Antagonistic effects of some endophytic fungi on five pathogenic fungi
3 讨 论
大量研究显示,植物内生真菌对多种植物病原真菌具有专一拮抗或广谱抗性[24-26],其拮抗机制通常表现为竞争、重寄生、溶菌和抗生作用等多种方式[27]。试验结果表明,供试20 株樟叶越桔嫩枝内生真菌对3 种木腐菌和5 种植物病原真菌均具有较强的拮抗活性,但不同的内生真菌对不同病原真菌的拮抗作用表现出一定的选择性或专一性。如内生真菌生长速度较快,与病原真菌相互竞争营养、空间,导致病原真菌不能正常繁殖,有产生抑菌圈而使病原真菌菌丝生长受到抑制,也可能是内生真菌的自身代谢产物具有溶菌和抗生作用,而表现出较强的拮抗作用。樊苗苗等[19]和陈肖学等[20]发现34 株樟叶越桔叶内生真菌中,曲霉属Aspergillussp.、叶点霉属Phyllostictasp.和鹿角菌属Xylariasp.等10 株菌株对小麦雪腐病菌、三七丝核病菌、灰葡萄孢、腐皮镰孢和芸苔链格孢等植物病原真菌具有较强的抑制活性,与其相比,樟叶越桔嫩枝中的内生抑菌活性菌株种类显得更为丰富。陈敏等[28]报道氯化钙结合生防菌株季也蒙毕赤酵母(VDBF-18)对芒果采后炭疽病具有较好的抑制效果,无柄盘菌属Peziculasp.[29-30]、间座壳属Diaporthesp.[31-35]和Cytospora属[36-38]等内生真菌能够产生多种抑菌活性天然产物,而层出盘菌属菌株VDBF-6、Helotiales属菌株VDBF-16 和葡萄座腔菌属菌株VDBF-25 等的抑菌活性则未见报道。本次试验仅测定了樟叶越桔嫩枝内生真菌对3 种木腐菌和5 种植物病原真菌的拮抗活性,围绕上述强拮抗活性菌株,进一步深入开展其与宿主的互作关系、活性次生代谢产物及其抑菌机理等相关研究,将为植物病害生防菌剂的进一步开发利用提供理论依据。
4 结 论
供试20 株樟叶越桔嫩枝内生真菌对3 种木腐菌和5 种植物病原真菌均具有较强的拮抗活性,其中季也蒙毕赤酵母(VDBF-18)、Neofusicoccum vitifusiforme(VDBF-25)、Pezicula heterochroma(VDBF-13)、Helotialessp.(VDBF-16)、胶孢炭疽菌(Colletotrichum gloeosporioides,VDBF-23)、Diaporthe nobilis(VDBF-11) 和Diaporthe rudis(VDBF-37)等7 株内生真菌对绵腐卧孔菌呈现强的拮抗作用。有14 株内生真菌对5 种病原真菌均呈现较强及以上拮抗能力,其中Pezicula neosporulosa(VDBF-2)对芸苔链格孢,VDBF-11 和Cytosporasp.(VDBF-42)对禾谷镰孢,VDBF-18 对灰葡萄孢,VDBF-37 对腐皮镰孢,Proliferodiscussp.(VDBF-6)对禾谷镰孢和灰葡萄孢,分别呈现强拮抗能力。
