川西北高原3 种禾本科牧草根系特征比较研究
2021-03-22刘斯莉王长庭张昌兵胡雷唐立涛潘攀
刘斯莉 ,王长庭 *,张昌兵 ,胡雷 ,唐立涛 ,潘攀
(1. 西南民族大学青藏高原研究院,四川成都610041;2. 四川省草原科学研究院,四川成都611731)
青藏高原位于亚洲中部,平均海拔超过4000 m,是地球上最为独特的生态单元,素有“地球第三极”之称,其对于欧亚大陆的径流调节、生物多样性保护以及维持碳收支平衡等方面发挥重要的生态安全屏障作用[1]。同时,青藏高原也是气候变化敏感区和生态脆弱带[1]。高寒草甸作为青藏高原的代表性植被类型,广泛分布于高原东部及其周围山地,其面积约占青藏高原的48%,是当地牧民生计与畜牧业发展的基础[1-2]。然而,随着气候变化及人类活动干扰的加剧,青藏高原高寒草甸生产力持续下降,约1/3 的高寒草甸生态系统面临着严峻的退化问题,严重制约了草地生态系统和高原畜牧业的可持续发展,对我国生态安全造成了较大威胁[3-4]。为了遏制高寒草甸生态系统进一步退化,国内外学者对退化高寒草甸的治理恢复进行了大量研究。减牧、灭鼠、施肥、补播、围栏封育以及人工草地建植等措施对退化高寒草地的改良和恢复具有积极作用[5-6]。其中,建植人工草地作为改善土壤质量和调节植物群落结构的重要管理措施,不仅能够加快植被的正向演替、提高草地生产力,还对维持高寒草甸生态系统各项生态过程和生态功能影响深远[4]。但目前针对建植人工草地的研究多集中于地上部分(地上生产力、物种多样性及群落结构)[7-9],对于植物地下部分特别是根系动态变化特征及空间分布格局的研究鲜见报道。
根系作为连接植物与土壤的动态界面,对植物生长、发育及繁殖具有重要意义。根系通过穿透和根际效应,帮助植物吸收营养物质,并将土壤养分产生的化学信号传递到植物地上部分,从而控制地上部分器官的生理活动[10]。因此,植物根系对于连接植物与土壤间的物质循环和能量流动过程具有关键作用[10-11]。其中根系特征是养分循环以及土壤形成等诸多生态系统过程的重要驱动因素,不仅决定着植物对土壤资源的利用效率,也反映了植物对环境的适应策略[12]。另外,根系的死亡与分解作为植物向土壤系统输入养分的主要途径[13],不仅影响植物地下生物量的分配及生态系统中的养分循环过程,还在一定尺度上反映气候、土壤和生物的总体特征[14]。但由于根系生长动态监测的复杂性与寿命估计的局限性,限制了对根系动态特征的深入了解[15]。
微根窗技术(minirhizotron method)[16]作为一种非破坏性的研究方法,克服了传统方法的缺陷,可在多个时段原位重复观测和记录根系生长、死亡的整个过程,为根系生产力、寿命及周转等方面的研究提供了便利。因此,本研究通过微根管技术原位监测青藏高原东缘人工草地建植中常用的3 种禾本科牧草紫羊茅(Festuca rubra)、垂穂披碱草(Elymus nutans)及老芒麦(Elymus sibiricus)单播草地的根系生长动态,并通过室内分析的方法,对不同栽培牧草的土壤理化性质进行了测定。旨在探讨:1)不同栽培牧草间根系现存量、生产量等动态变化特征及空间分布格局的变化;2)不同栽培牧草间根系周转对碳分配特征的影响机制。以期阐明不同栽培牧草在该区域高寒环境条件下根系的适应策略,为川西北退化草地生态系统恢复与治理提供理论依据。
1 材料与方法
1.1 研究区域概况
试验地设在四川省草原科学研究院牧草育种基地,该基地处于川西北阿坝州红原县境内(32°49′N,102°35′E),平均海拔3494 m,属大陆性高原寒温带半湿润季风气候,年均气温1.1 ℃,最热月(7 月)均温10.9 ℃,最冷月(1 月)均温-10.3 ℃,年日照时数2417.9 h,年降水量650~800 mm,无绝对无霜期,草甸类型主要是矮嵩草草甸,植被平均盖度80%以上,植被最高高度可达45~60 cm。土壤类型为亚高山草甸土[17]。
1.2 试验样地设计
于2015 年4 月上旬,清除试验地杂草,翻耕土地(深度20~30 cm),精细平整。5 月初开始建植,选用紫羊茅、垂穂披碱草和老芒麦进行单播,播种量为37.5 kg·hm-2,行距25 cm,播种后覆土1~2 cm;以150 kg·hm-2混合肥作基肥,每种牧草试验地为5 hm2。播种当年出苗后,清除杂草1~2 次;第2 年后,每年进行除草2~3 次,其余田间管理同当地大田管理一致[18]。在每个试验地内随机选择3 个面积为20 m×20 m 的平行样地,共计9 个样地。于2015 年9 月在每个样地内,随机选取1 个地上植被相对均匀的区域,安装1 根聚碳酸酯透明材料微根管(长100 cm,内径 5 cm,外径 6 cm),微根管与地面夹角为30°,每个样方埋入 1 根,共计9 根微根管[17]。
1.3 样品采集与处理
1.3.1 土壤样品采集与测定 于2016 年8 月中旬,在每个样地内随机选取1 个50 cm×50 cm 的样方,采用内径5 cm 的土钻在每个样方内按“S”形分两个土层(0~10 cm 和10~20 cm)取5 钻土样,分别混合为1 个土壤样品,并做好标记。待样品风干后,分别过1.00 和0.15 mm 筛用于测定土壤理化性质。其中,采用杜马斯燃烧法测定全碳(total carbon,TC)和全氮(total nitrogen,TN)含量[19],采用钼锑比色法测定全磷(total phosphorus,TP)含量,采用丘林法测定土壤有机质(soil organic matter,SOM)含量,采用电极法测定pH[20],采用烘干法测定含水量(soil water content,SWC)[21]。
1.3.2 根系图片采集与处理 于2016 年5-9 月对试验区的微根管进行连续取样。首先拆开微根管外包裹的塑料袋,打开顶端盖子,使微根管内部的湿气充分与外部气流交换,让管子内外的温差平衡,然后用探杆检查管子底部是否有积水,并擦拭管子内壁水汽,标记取样的初始位置后,将探头缓慢放入微根管内,连接电脑采用CI-600根系扫描仪(CID Bio-Science,Camas,WA,USA)进行图像采集。每根微根管按垂直土层深度分为0~10 cm 和10~20 cm 土层分别采集图像数据,每次测定结束后,用黑色塑料袋将微根管及裸露部分包裹严密,每间隔15 d 采集1 次,共取10 次图像,总共追踪到8837 条根系。
采用WinRHZIO Tron MF(CID Bio-Science,Camas,WA,USA)软件对采集的图像进行分析处理,测定每张图像中根系长度及根尖数等各形态指标,通过图像中根系的颜色来区分活根和死根,将白色和褐色的根定义为活根,黑色的根定义为死根。每次图片处理后导入上次数据,识别上次的根系,同时记录新生根系,每条根都有独立的编号,完整记录所有根系从起始到死亡的长度变化[17]。
由根系图像分析得到根系的长度,对t时不存在而在t+1 时出现的所有新根根长求和,加上在t时原有根系上延长的长度和来估算根系生产量;利用t和t+1 间隔时死亡和消失掉的根长及在t+1 时出现的死根根长的总和估算根系死亡量;根系现存量则用每一时刻的活根长度来估算[22-23]。利用如下公式计算:

式中:L(mm)为微根管中观察到的根系长度;A(cm2)为观测窗面积(图像大小)[23],本研究中A取 400 cm2。
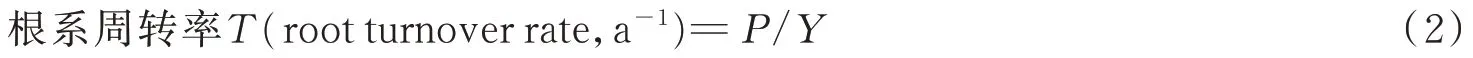
式中:P表示根系年生产量;Y表示根系年均现存量。
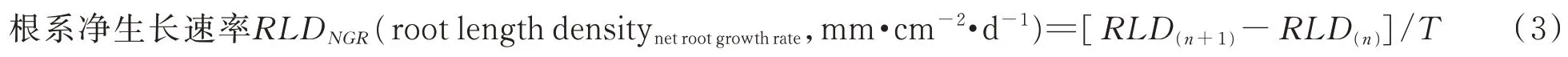
式中:RLD(n+1)和RLD(n)分别为第n+1 与n次观测中的单位面积根系现存量,T为相邻两次观测的间隔天数。RLDNGR为正值表示根系的生长速率大于死亡速率,即正生长;RLDNGR为负值表示根系负生长;RLDNGR为0 表示根系零生长[24-25]。
1.4 数据统计与分析
采用SPSS 22.0 软件对数据进行统计分析,Origin 8.5 作图。利用Kaplan-Meier 方法计算根系存活率,并估计根系平均寿命。对土壤(理化性质、生态化学计量比)和植物根系特征等数据进行单因素方差分析(One-Way ANOVA);采用最小显著极差法(LSD)进行不同处理之间的显著性检验(P<0.05),不同土层间的差异则采用独立样本t检验;采用双因素方差(Two-Way ANOVA)分析不同牧草种类、土层及其相互作用对根系特征的影响。应用Fuzzy 数学隶属函数对3 种牧草的根系特征进行综合分析。采用Canoco 4.5 对根系特征和环境因子间的对应关系进行冗余分析(redundancy analysis,RDA)。利用Amos 23.0 构建根系特征和环境因子间的结构方程模型(structural equation model,SEM)。
2 结果与分析
2.1 土壤理化性质
由表1 可知,0~10 cm 土层,老芒麦土壤TC、TN 和SOM 含量高于垂穗披碱草及紫羊茅,但差异不显著;老芒麦土壤C/P、N/P 及SWC 显著高于紫羊茅(P<0.05),其土壤pH 显著低于垂穗披碱草和紫羊茅(P<0.05)。10~20 cm 土层,老芒麦土壤TC 和TN 均显著高于垂穗披碱草及紫羊茅(P<0.05),垂穂披碱草土壤TP、SOM 显著低于紫羊茅和老芒麦(P<0.05)。此外,3 种栽培牧草 0~10 cm 土层TC、TN、C/N、SOM 及SWC 均高于 10~20 cm,10~20 cm 土层pH 高于0~10 cm,但差异不显著。

表1 3 种栽培牧草土壤理化特征Table 1 Soil physicochemical property of three cultivated herbage species
2.2 根系季节动态特征
2.2.1 根尖数 3 种栽培牧草根尖数随生长季逐渐增加,并在7 月中旬至8 月初达到生长峰值,随后逐渐降低。其中,0~10 cm 土层,老芒麦及垂穂披碱草根尖数峰值均出现于8 月1 日,分别为554.67 和470.00 根,而紫羊茅根尖数峰值出现于7 月17 日为268.50 根。10~20 cm 土层,紫羊茅、垂穂披碱草及老芒麦根尖数峰值分别出现于 6 月 1 日为 242.00 根、7 月 17 日为 344.33 根及 8 月 1 日为 298.67 根(P<0.05,图 1)。
2.2.2 根系现存量、根系生产量、根系死亡量及其季节动态 3 种栽培牧草0~10 cm 和10~20 cm 土层根系现存量季节变化差异显著。其中,垂穗披碱草和老芒麦0~10 cm 和10~20 cm 土层根系现存量随生长季逐渐增加,并在8 月中旬基本保持稳定。紫羊茅0~10 cm 和10~20 cm 土层根系现存量均于7-8 月逐渐降低(图2A,B),此外,紫羊茅根系现存量低于垂穗披碱草和老芒麦(图2C)。
3 种栽培牧草 0~10 cm 和 10~20 cm 土层根系生产量在生长季整体表现为降低的趋势。其中,0~10 cm 土层,垂穗披碱草根系生产量在生长季波动较大,7-8 月出现两次峰值(图2D)。而 10~20 cm 土层,紫羊茅和垂穂披碱草根系生产量均表现出随生长季先增后减的趋势(图2E)。另外,3 种栽培牧草0~10 cm根系生产量差异不明显,而10~20 cm 紫羊茅根系生产量显著高于垂穂披碱草及老芒麦(P<0.05,图2F)。

图1 3 种栽培牧草的根尖数动态变化Fig. 1 Seasonal dynamics of root tips of three cultivated herbage species
3 种栽培牧草 0~10 cm 和 10~20 cm 土层根系死亡量在生长季整体表现为递增趋势。其中,0~10 cm土层,3 种栽培牧草根系死亡量季节动态大致趋同,且三者均于8 月中旬出现死亡高峰(图2G);10~20 cm土层,紫羊茅根系死亡量波动较大,且生长季内每月死亡量均高于垂穂披碱草和老芒麦(图2H)。此外,3 种栽培牧草0~10 cm 根系死亡量无显著差异,而10~20 cm紫羊茅根系死亡量显著高于垂穂披碱草及老芒麦(P<0.05);紫羊茅根系死亡量随土层加深显著增加,而垂穂披碱草和老芒麦则显著降低(P<0.05,图2I)。
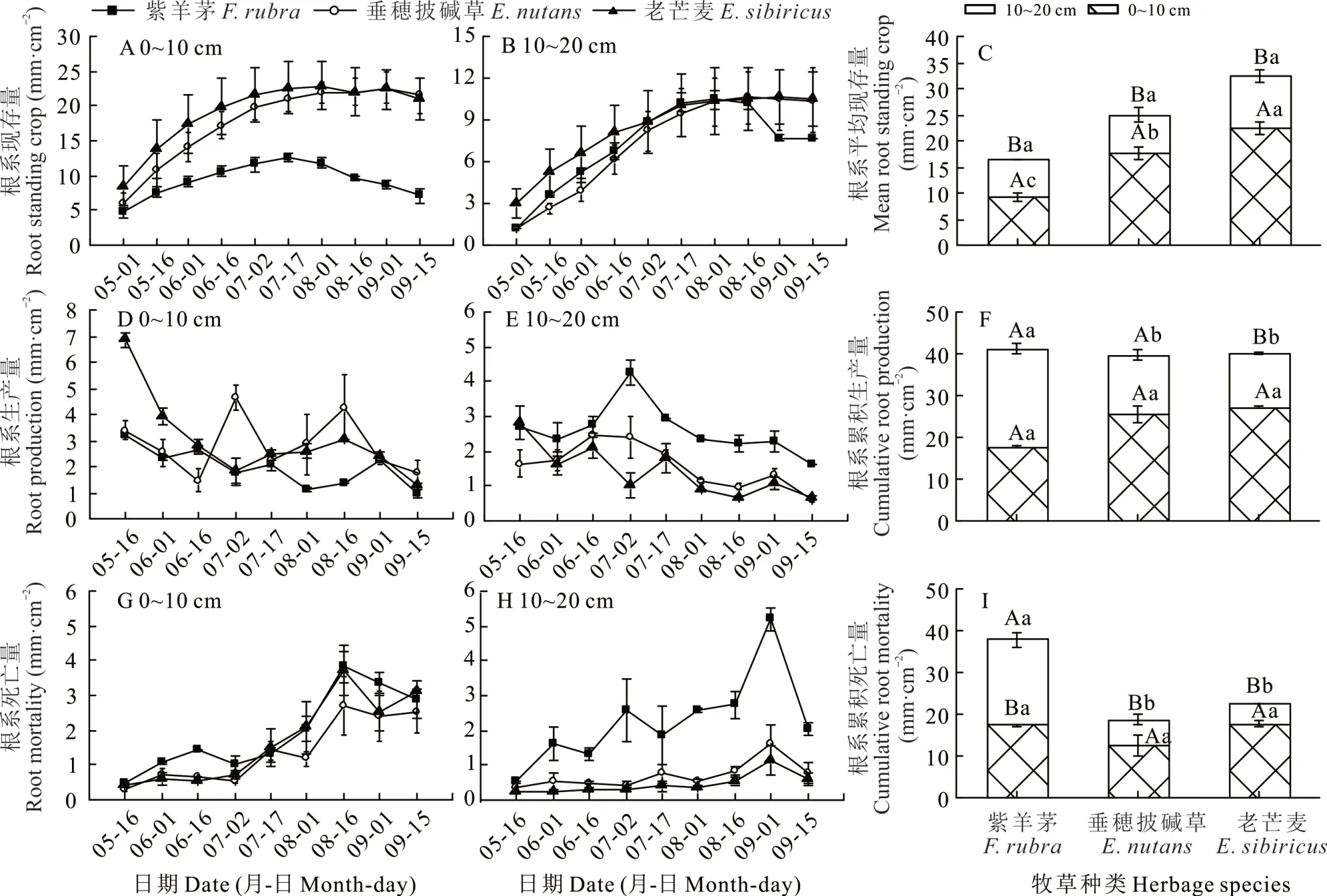
图2 3 种栽培牧草根系生长季节动态Fig.2 Seasonal dynamics of root growth of three cultivated herbage species
2.2.3 根系净生长速率 在0~10 cm 和10~20 cm 土层中,3 种栽培牧草根系净生长速率随生长季呈现波动式下降趋势,于5-6 月达到正生长高峰,8-9 月达到负生长高峰。0~10 cm 土层,3 种牧草同期达到生长峰值(5月16 日),而10~20 cm 垂穂披碱草正生长峰值出现时间最晚(6 月16 日)。生长季内0~10 cm 根系净生长速率高于 10~20 cm 土层,但更早进入负增长阶段(8 月 1 日,图3)。

图3 3 种栽培牧草根系净生长速率的差异Fig.3 Differences in RLDNGR of three cultivated herbage species
2.3 根系寿命与周转率
由图4 可见,在0~10 cm 和10~20 cm 土层中,紫羊茅的根系周转率均显著高于老芒麦及垂穂披碱草,而老芒麦及垂穂披碱草根系寿命显著高于紫羊茅(P<0.05)。此外,3 种栽培牧草的根系周转率均随土层加深而增加,0~10 cm 紫羊茅及垂穂披碱草根系寿命显著高于10~20 cm,老芒麦则相反(P<0.05)。

图4 3 种栽培牧草根系周转率及根系寿命的差异Fig.4 Differences in root turnover rate and root longevity of three cultivated herbage species
2.4 根系特征综合评判
应用模糊数学中的综合隶属函数对3 种栽培牧草根系特征进行了综合评价,结果显示:0~10 cm 土层,表现为老芒麦>垂穂披碱草>紫羊茅;10~20 cm 土层,表现为老芒麦>紫羊茅>垂穂披碱草(表2)。
2.5 根系特征与土壤环境因子的相关性分析
2.5.1 根系特征与土壤环境因子的冗余分析(RDA) 图5 显示,第一、二排序轴累计解释率分别为56.1%、27.7%,根据图中实线与虚线的夹角可以看出根尖数、根系现存量、根系生产量与TC、TN、C/N、C/P、N/P、SWC、SOM 正相关,与 TP、pH 负相关;根系死亡量与 C/P、N/P、SWC、SOM 正相关,与 TC、TN、TP、pH 负相关;根系周转率与 TP、pH 正相关,与 TC、TN、C/N、C/P、N/P、SWC、SOM 负相关;根系寿命与TC、TN、TP、C/N、C/P、N/P、SWC 正相关。

表2 3 种栽培牧草各项根系生长指标隶属函数值及评判结果Table 2 Function value of subordination and result of comprehensive judgment on the root characteristics to three cultivated herbage species
2.5.2 根系特征与环境因子的结构方程模型分析(SEM) 构建结构方程模型,分析根系特征与土壤环境因子之间的相互影响关系。建模时对土壤生态化学计量比(C/N、C/P、N/P)、土壤理化性质(pH、SWC、SOM)及根系动态特征(根系现存量、根系生产量、根系死亡量)进行降维处理。提取第一维数据,解释率分别为67.26%、69.37%、68.89%。结果显示,牧草种类直接影响根尖数量和根系特征(生产量、死亡量和现存量),导致根系周转率发生显著变化,同时牧草种类差异也能够直接改变土壤化学计量比,进而影响根系特征。土层深度直接改变了土壤化学计量比和土壤理化性质,土壤养分条件的差异改变了3 种牧草根尖数和根系特征,促进了根系周转率,此外,土层深度对根系周转率也具有直接促进作用(图6)。

图5 根系特征与土壤环境因子的RDA 分析Fig. 5 Redundancy analysis for root characteristics and soil environmental factors
3 讨论
3.1 3 种栽培牧草的土壤环境比较

图6 根系特征与环境因子结构方程模型Fig.6 The structural equation model of root characteristics and environmental factor
土壤作为生态系统中营养物质循环、水分平衡、凋落物分解等生态过程的参与者与载体,其养分状况是评价退化生态系统功能恢复与维系的关键指标,对植物生长与群落演替发挥重要作用[26]。而建植人工草地,相当于引入新的植被,其恢复作用可有效增加地表凋落物和地下根系残体及其分泌物的输入量,进而改善土壤理化性质[27]。本研究显示,3 种栽培牧草中,老芒麦土壤TN 含量较高,这可能是因为老芒麦土壤含水量较高(表1),促使土壤微生物活性增强,进而通过加快土壤养分的矿化速度来提高植物对铵和硝酸盐的有效利用率,导致更高的净初级生产力,从而提高其土壤TN 含量[25]。而老芒麦土壤较高的N 含量,一定程度上说明其土壤可利用N 含量较高[28],当土壤N 有效性提高后,不仅可以促进地上部分获取更多的N,从而提高叶面积、增加光合产物,而且可将更多的C 输送到根部,提高根系产量,促使根系的N 含量增加,而组织N 含量与呼吸速率呈显著正相关,从而导致根系呼吸强度增大[29]。也有研究表明,土壤N 含量较高能提高土壤中阳离子浓度及P 的有效性,进而促进植物对P 更有效的吸收[30],使得植物营养需求与养分供应之间保持稳态,从而有利于生态系统中P 的转化与循环,最终促进了C、N 循环平衡[30]。另外,土壤肥力较高不但有利于演替后续物种的生长和繁殖,还能促进群落演替过程,进而加快人工草地的修复进程[31],土壤SOM 的分解提升了老芒麦土壤养分含量的整体水平[32],进而改善了老芒麦栽培草地的土壤肥力。本研究还发现,3 种栽培牧草0~10 cm 土层pH 均低于10~20 cm,原因是随着地上植被生长、死亡,在地表不断累积的凋落物,被分解转化为有机质和其他养分,这些养分为土壤微生物活动提供了有利条件,加快对腐殖质的分解,促使地表产生更多有机酸,从而降低表层土壤pH[33]。另外,土壤微生物的硝化作用属于好氧过程,多发生于通气条件较好的表层土壤中,其直接产物NH4+和H+也进一步增加了表层土的酸度[29]。
3.2 3 种栽培牧草根系特征的季节动态
气温和降水通过影响根系的呼吸速率以及对水分和矿质元素的吸收等生理过程,进而影响根系的生长发育,因此根系特征会随季节变化呈现规律性变化[34-35]。本研究发现,5 月初3 种栽培牧草净生长速率出现正生长高峰,这可能是因为生长季初期,高寒草甸开始返青,地上部生长发育缓慢,大部分光合作用产物储存于地下,有利于根系的发育和生长,地下生物量增高明显[36]。而10~20 cm 土层垂穂披碱草净生长速率正生长高峰期迟于其他两种栽培牧草,这可能是因为青藏高原地下生物量对P 元素变化比较敏感[17],而10~20 cm 土层垂穂披碱草土壤TP 含量又较低,导致植物对P 的利用率降低,从而限制了根系生长。其次,本研究发现3 种栽培牧草根系现存量7-8 月达到高峰,这可能是因为6 月底根系分布层完全解冻,伴随气温回暖,植物光合作用增强[37],同时,该区域7-8 月,水热条件较好,促进有机物分解和矿化,植物生产力和基质利用率提高,并促进土壤养分移动和根系吸收,使得植物各项生理特征渐趋成熟,根系实现大量生长[28,38]。其中,老芒麦和垂穂披碱草生长季后期的根系现存量比前期增加了2 倍左右(图2A),这是由于生长季初期,营养体萌发消耗了大量地下累积的物质,8 月前形成的物质大多用于建造营养体,当营养器官生长减缓时(生长季后期),植物才迅速在地下累积约五成以上的物质,以供来年营养体萌发[39]。此外,一定范围内,温度的升高有利于根系的生长发育,而低温胁迫则导致根系生长受阻[40]。8 月气温回升,土壤微生物活性增强,老化死亡根系数量增多,微生物分解作用加快,根系累积速率渐缓[41],而趋近9 月根系死亡量逐渐升至观测期内最高,这可能与土壤温度降低,植物受到低温胁迫有关[40]。另外,一定范围内,土层越深,土壤温度和水分对气温和降水变化响应的波动越小[35]。因此,生长季后期,对植物根系生长不利的水热组合条件大致呈现“先表层—后深层”的规律,从而导致表层根系更早进入负增长状态。
3.3 3 种栽培牧草的根系特征及分布格局
植物根系作为生态系统碳分配及碳循环的关键环节,对生态系统功能的发挥及地下微环境的变化具有重要作用,其分布格局及现存量决定着植被对地下水分和养分的利用效率,同时分布格局及现存量也受到外界环境因素(增温降水等)的影响[17,42]。本研究发现,老芒麦及垂穂披碱草的根系生长具有明显的“表聚性”,这是由于高寒地区0~10 cm 土层具有较好的保水及保肥能力,植物为了获取更多的养分和水分,满足其生长需求,便将根系集中分布于此层[43]。另外,根系呼吸需要氧气,表层土壤通气状况较好,植物将根系集中分布于此层以获得更多氧气,使表层根际形成微域“氧化圈”,这不仅缓解了一些还原性物质对根系的毒害作用,同时也促进了硝化作用等好氧过程的发生,对根际氮循环意义重大[44]。而紫羊茅根系生产与死亡多发生于10~20 cm 土层,这可能与其土壤TN 含量较低有关。有研究发现,在低N 胁迫下,植物主要采用伸长根系作为适应策略,以获得更多的土壤空间和氮资源[45]。另有研究发现,根系长度不仅能反映植物根系生长状况以及在土壤中的拓殖能力,还能反映根系对土壤空间异质性的适应能力[46]。本研究中老芒麦的根尖数量及现存量较高,表明其根系密度较大,根系对土壤营养吸收利用率较高,说明老芒麦具有更强的抵御外界不利因素的能力。
植物根系的动态变化不仅体现在现存量、生产量及死亡量上,同时也体现在根系周转率和寿命等方面。根系周转作为生态系统营养动力学和储量的重要组成部分,其向地下生态系统输入的有机物达到输入总量的50%左右[17]。而根系寿命作为根系周转的重要内在机制,与根系周转率负相关,即根系寿命越长,根系周转越慢[47],其对C 的消耗也越小。本研究表明,随土壤N 有效性的提高,老芒麦根系现存量增加,根系周转率呈降低趋势。这与Burton 等[48]研究结果相似,其主要机理是,1)根据投入与收益原则,土壤有效N 增加,植物延长根系寿命的收益(吸收养分)高于维系根系生长的成本(消耗C),故根系采取长时间占据养分空间以吸收更多养分的生长策略。2)根据根系碳汇强度理论[48],假设土壤N 有效性增强,则将促进根系对养分的吸收效率,促使根系成为一个相对较强的“碳汇”,这些C 主要用于维持根系呼吸作用、养分吸收效能以及同化速率,故根系具有相对充足的C 以延长寿命,由此根系周转率减慢。
3.4 根系特征与环境因子的关系
植物根系可通过自身构型的可塑性对植物生长状况及外界环境的变化(土壤养分、水分、温度等)做出适应性调整,并表现出不同的特征[46]。因此,根系特征可作为植物响应环境条件变化的指示参数,反映土壤中物质与能量吸收利用的程度及转化率[49]。本研究SEM 分析表明,土壤理化性质及生态化学计量比直接影响根系动态特征,并对根系的生长具有直接的促进作用,这是因为植物根系与土壤之间的接触面积大,在植物与土壤进行频繁的物质能量交换时,彼此强烈影响,因而土壤作为植物的一个重要生态因子,土壤条件(土壤理化与生物学特性等)可影响植物地上和地下部分的生长与产量。另外,牧草种类通过土壤生态化学计量比间接影响根系动态特征,这是由于3 种牧草种类存在种间差异,导致不同植物物种对土壤养分的吸收利用方式及对土壤条件的适应性不同[49],而根系在土壤营养含量等环境梯度及物种间互作机制下具有高度的形态可塑性[12],这直接影响植物根系的生长、发育、分布及生产力,从而导致3 种牧草根系动态特征存在明显差异。
本研究还发现,土层深度通过影响土壤理化性质间接使得根系生物量在剖面上呈现出“倒金字塔”分布。这是由于随土层加深,土壤温度、养分、含水量及通气量等环境条件趋于恶劣,植物则会为了适应生态环境的变化采用减少根系生产以降低自身养分消耗的策略,从而抑制了10~20 cm 土层的根系生物量[37,50]。其次,土层深度与根系周转率呈正相关,这归因于禾本科牧草是须根系植物,0~10 cm 土层中根系现存量虽多,但大多由固定和支撑植物的不定根构成,其对土壤养分的吸收和利用能力较低[51-52],故其周转率相对较慢,而10~20 cm 土层的根系多由不定根上生成的侧根构成,其主要功能是吸收水分、养分等必要的营养物质[53],故其生产效率较高,周转率较快。
4 结论
3 种禾本科牧草根系的变化具有明显的季节特征,根系现存量随生长季显著增加,在8 月中旬趋于稳定,而根系死亡量随生长季波动式增加,在生长季末期(9 月初)达到峰值。且3 种禾本科10~20 cm 的根系周转率高于0~10 cm,土层深度是影响根系周转率的主要因子。其次,土壤条件对根系生长具有直接的促进作用,较高的土壤TN 含量可以提高根系现存量。另外,老芒麦和垂穂披碱草的根尖数、根系现存量较高,根系寿命长,周转率高,紫羊茅根系寿命较短,根系周转快。因此,3 种禾本科栽培牧草的根系特征具有差异性,是植物根系在适应高寒环境中权衡碳分配、碳消耗采取的生态适应策略。
