南方红豆杉天然居群叶片的表型性状和氮磷化学计量特征的变异规律
2021-03-22罗芊芊周志春邓宗付喻龙华孙建军徐刚标
罗芊芊, 周志春, 邓宗付, 喻龙华, 孙建军, 徐刚标
(1. 中国林业科学研究院亚热带林业研究所 浙江省林木育种技术研究重点实验室, 浙江 杭州 311400;2. 中南林业科技大学林学院, 湖南 长沙 410004; 3. 中国林业科学研究院 亚热带林业实验中心, 江西 分宜 336600)
在长期的自然生长过程中,植物通过调节自身的资源配置适应特定的生境,在不同生境下演化出各自特有的生长特性和物质分配规律[1]。植物的表型性状反映了基因型对环境变化的适应性,分析植物表型性状变异对研究植物的环境适应性和演化有重要意义[2],是植物遗传多样性研究的重要内容之一[3],并广泛用于揭示植物天然居群的遗传变异规律及地理分布格局[4-5]。植物叶片形态与其营养元素组成等一系列生理生态因子密切相关[6],研究叶片氮磷化学计量特征有助于了解植物对外界环境的生长适应策略[7],其中,叶片氮磷比在指示生态系统限制性元素方面应用广泛[8]。
南方红豆杉〔Taxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li〕是集药用、材用和观赏于一体的珍稀濒危树种[9-11],主要分布于中国东部和南部地区,其中,浙江、湖南、江西、福建和广东等省的南方红豆杉天然资源蕴藏量较多。南方红豆杉幼树生长需要特定的温度、湿度和光照条件,但由于群落中乔木层郁闭度大、林内光照有限,导致南方红豆杉易出现小范围聚集分布现象,其天然居群的个体多以老龄为主、幼龄为辅,幼龄个体的数量和生长速度难以支撑居群的扩展进程[12-13]。迄今为止,研究者已对南方红豆杉的濒危机制[14]、生理生态特征[15-16]、基因流[17]、基因克隆[18]和遗传多样性[19]等方面进行了深入研究。谢伟东[19]对南岭山地南方红豆杉天然居群遗传多样性的研究结果表明:南方红豆杉不同居群间的分化程度较高,且叶片性状在不同居群间存在显著差异,这种差异可能与地理位置和气候等因子有关,加之居群间和居群内存在自然杂交,导致不同居群产生了不同程度的分化。
植物叶片的可塑性较大且对生境变化较敏感,生境的改变常导致叶片形态及氮磷化学计量特征的变化[20]。南方红豆杉为典型的阴性树种,对生境变化较为敏感。费永俊等[21]的观察结果表明:当南方红豆杉由高海拔山地引种至低海拔平原湖区后,一些个体的叶片和孢子形态发生了变化,但其化学计量特征是否存在显著差异仍没有明确的研究结论。Du等[22]对全球171个地点优势树种的叶片进行了研究,发现全球自然陆地生态系统(农田、城市和冰川除外)中有18%的区域受到较强的氮限制,而43%的区域受到较强的磷限制,其他39%的区域则受氮和磷的共同限制或氮与磷任一元素的微弱限制。总体上看,氮限制在苔原、北方针叶林、温带针叶林以及山地草原和灌丛较为普遍;磷限制在热带及亚热带森林,温带阔叶林,沙漠和地中海植被,热带、亚热带和温带草原,以及稀树草原和灌丛较为常见[22]。植物生长的限制性元素是否会随生长环境不同而改变?这种变化在不同居群间是否具有普遍性?这些疑问均有待进一步研究。
对南方红豆杉不同居群间叶片的表型性状及氮磷化学计量特征的变异进行研究,不仅可以揭示环境变化对南方红豆杉分布的影响,还可以为其生长的限制性元素研究提供参考依据,从而为南方红豆杉天然居群的繁衍和保护提供帮助。作者以中国东部和南部地区分布的南方红豆杉天然居群为研究对象,对叶片的表型性状和氮磷化学计量指标进行比较,从形态学、数量学和生态化学计量学等方面揭示叶片表型性状在居群间和居群内的变异状况以及叶片氮磷化学计量指标在居群间的差异,分析叶片的表型性状和氮磷化学计量指标以及地理-气候因子间的相关关系,以期为南方红豆杉种质资源遗传保育和开发利用提供基础研究数据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于中国东部和南部,包括浙江省的临安、丽水、江山和龙泉,湖南省的桑植、浏阳、新晃和桂阳,江西省的婺源、铜鼓、分宜和信丰,福建省的南平、明溪、福州和武平,以及广东省的乐昌和连州,共18个南方红豆杉天然居群。地理位置为东经109°05′~119°56′、北纬24°55′~30°01′,海拔300~990 m,供试各居群的地理-气候概况见表1,各居群的经度、纬度和海拔采用GPS记录,气象数据提取自DIVA-GIS的数据包。
研究区的地带性植被为常绿及落叶阔叶林,土壤多为红黄壤;居群内的主要伴生树种为毛竹〔Phyllostachysedulis(Carrière) J. Houzeau〕、杉木〔Cunninghamialanceolata(Lamb.) Hook.〕和枫香树(LiquidambarformosanaHance)等。
1.2 研究方法
1.2.1 样株选择和样品采集 于2018年6月至7月,在18个南方红豆杉天然居群中选择株龄80~100 a的成年个体为样株;为防止样株间存在半胞亲缘关系,样株间距均大于30 m。根据居群的规模确定样株数:若居群中成年植株数量少于或等于50株,全部作为样株;若居群中成年植株数量多于50株,则随机选择50株作为样株。18个居群共选择样株670株,各居群的样株数见表1。
在各样株冠层中部的向阳面,随机采集无病虫害的2年生叶片50 g,装入塑封袋并置于冰盒中,带回实验室,于4 ℃保存、备用。因各居群的样株分布较为分散,故未采集土样。
1.2.2 表型性状测定 每株随机选取90枚完整叶片,分成3组,记为3次重复。采用Epson Perfection LA2400扫描仪〔爱普生(中国)有限公司〕扫描叶片,并采用WinRHIZO软件测定叶片的叶长(叶基至叶尖的距离)、叶宽(叶片最宽处的宽度)、叶面积和叶周长,测量精度为0.01 cm;根据叶长和叶宽计算叶宽长比。
1.2.3 氮和磷含量测定 将扫描后的叶片以居群为单位混合,置于105 ℃下杀青30 min,然后于80 ℃烘干至恒质量;冷却后研磨,过2 mm筛,备用。
分别称取各居群叶片干粉3份,每份2.0 g,共计54份样品,采用凯氏定氮法[8]和电感耦合等离子体发射光谱法[23]检测叶片中的氮和磷含量;根据叶片的氮和磷含量计算氮磷比。
1.3 数据处理
采用EXCEL 2016软件对实验数据进行整理;参照文献[24],采用SAS 8.0软件中的GLM过程进行表型性状方差分析;参照文献[25]中的方法计算表型分化系数。
对叶片的表型性状和氮磷化学计量指标间以及叶片表型性状与地理-气候因子间的相关性进行分析,并依据叶片表型性状对18个居群进行聚类分析。采用SPSS 10.0软件进行多重比较(Duncan’s新复极差法)、相关性分析和聚类分析(中位数聚类法),并计算各性状的均值、标准差和变异系数。

表1 南方红豆杉18个天然居群的地理-气候概况和样株数
2 结果和分析
2.1 南方红豆杉各居群间叶片的表型性状和氮磷化学计量指标的差异
南方红豆杉18个天然居群叶片的表型性状和氮磷化学计量指标的比较结果见表2;各表型性状和氮磷化学计量指标的方差分析结果见表3;各表型性状的方差分量和表型分化系数见表4。
2.1.1 叶片表型性状的比较 由表2可见:南方红豆杉不同居群间叶片表型性状差异明显,叶长、叶宽、叶面积、叶周长和叶宽长比的变幅分别为2.32~3.00 cm、0.33~0.48 cm、0.64~1.12 cm2、5.43~7.03 cm和0.14~0.17,均值分别为2.62 cm、0.38 cm、0.81 cm2、6.11 cm和0.15;各性状的变异系数为12.79%~29.56%,其中,叶宽的变异系数最小,叶宽长比的变异系数最大。
在供试的南方红豆杉18个天然居群中,福建武平居群的叶长(3.00 cm)、叶宽(0.48 cm)、叶面积(1.12 cm2)和叶周长(7.03 cm)总体上显著(P<0.05)大于其他17个居群,该居群的叶宽长比(0.16)也较大;浙江龙泉居群的叶长和叶周长最小(分别为2.32和5.43 cm),浙江丽水居群的叶宽最小(0.33 cm),湖南桂阳居群的叶面积最小(0.64 cm2),浙江丽水、浙江江山、湖南浏阳、江西婺源、江西分宜和江西信丰居群的叶宽长比最小(均为0.14)。
方差分析结果(表3)显示:南方红豆杉的叶长、叶宽、叶面积、叶周长和叶宽长比在居群间和居群内的差异均达到极显著(P<0.01)水平。
2.1.2 叶片氮磷化学计量指标的比较 由表2还可见:南方红豆杉不同居群间叶片氮磷化学计量指标也存在明显差异,氮含量、磷含量和氮磷比的变幅分别为14.30~20.83 g·kg-1、0.96~1.61 g·kg-1和10.62~22.28,均值分别为17.81 g·kg-1、1.22 g·kg-1和14.94;各指标的变异系数为11.13%~20.90%,其中,氮含量的变异系数最小,氮磷比的变异系数最大。
在供试的南方红豆杉18个天然居群中,浙江临安、江西分宜、江西信丰和广东连州居群的叶片氮含量均在20.00 g·kg-1以上,且显著高于除江西婺源外的其他居群;而福建明溪居群的叶片氮含量最低(14.30 g·kg-1),且与其他居群差异显著。江西婺源居群的叶片磷含量最高(1.61 g·kg-1),且与其他居群差异显著;而江西分宜和福建武平居群的叶片磷含量最低(0.96 g·kg-1),且与多数居群差异显著。江西分宜居群的叶片氮磷比最大,且与其他居群差异显著;而浙江丽水居群的叶片氮磷比最小,且与多数居群差异显著。
方差分析结果(表3)显示:南方红豆杉的叶片氮含量、磷含量和氮磷计量比在居群间的差异均达到极显著水平。

表2 南方红豆杉18个天然居群间叶片的表型性状和氮磷化学计量指标的比较

表3 南方红豆杉天然居群叶片的表型性状和氮磷化学计量指标的方差分析结果1)
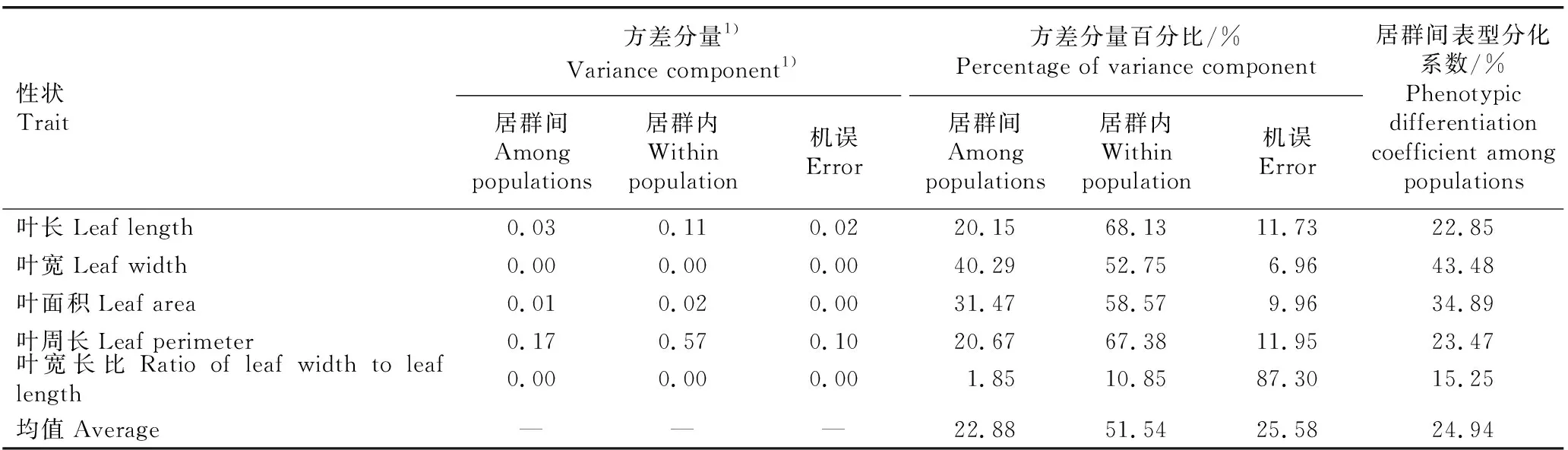
表4 南方红豆杉天然居群叶片表型性状的方差分量和表型分化系数
2.1.3 叶片表型性状的方差分量和表型分化系数
由表4可见:南方红豆杉天然居群叶片的5个表型性状在居群间和居群内的方差分量百分比均值分别为22.88%和51.54%,居群内的方差分量百分比明显大于居群间,说明南方红豆杉叶片表型性状的变异主要来源于居群内。
从叶片5个表型性状的居群间表型分化系数看,叶宽的居群间表型分化系数最大,为43.48%;而叶宽长比的居群间表型分化系数最小,仅为15.25%;各表型性状的居群间表型分化系数均值为24.94%,说明南方红豆杉天然居群叶片表型性状的变异约25%来自居群间。
2.2 南方红豆杉叶片的表型性状和氮磷化学计量指标以及地理-气候因子间的相关性
南方红豆杉天然居群叶片的表型性状和氮磷化学计量指标间的相关系数见表5;叶片表型性状与地理-气候因子间的相关系数见表6。
由表5可见:在南方红豆杉叶片的5个表型性状中,仅叶长与叶宽长比无显著相关性,其他表型性状间均极显著(P<0.01)或显著(P<0.05)正相关,说明叶片较长,叶片的宽度和面积也较大。在3个氮磷化学计量指标中,氮磷比与氮含量显著正相关,与磷含量极显著负相关,但氮含量与磷含量无显著相关性。在表型性状和氮磷化学计量指标间,仅氮磷比与叶长显著负相关,并与叶面积和叶周长显著正相关,氮含量和磷含量与各表型性状均无显著相关性,说明叶片氮磷比越大叶片越宽大。
由表6可见:叶片各表型性状与年均温、最冷月最低温、年均降水量、最湿月降水量和最干月降水量无显著相关性,与纬度和海拔也无显著相关性,但叶长、叶面积和叶周长与经度显著负相关,说明随经度增大叶片趋于短小;叶宽长比与最热月最高温极显著负相关,说明随最热月最高温增高叶片趋于细长。

表5 南方红豆杉天然居群叶片的表型性状和氮磷化学计量指标间的相关系数1)

表6 南方红豆杉天然居群叶片表型性状与地理-气候因子间的相关系数1)
2.3 南方红豆杉天然居群的聚类分析结果
基于南方红豆杉18个天然居群的5个叶片表型性状进行聚类分析,结果见图1。由图1可见:在欧氏距离15处,可将18个居群分为2类,第1类包含10个居群,第2类包含8个居群;在欧氏距离8处,可进一步将第1类分为2个亚类(第1和第2亚类),第2类分为3个亚类(第3、第4和第5亚类)。

P1: 浙江临安Lin’an of Zhejiang; P2: 浙江丽水Lishui of Zhejiang; P3: 浙江江山Jiangshan of Zhejiang;P4: 浙江龙泉Longquan of Zhejiang; P5: 湖南桑植Sangzhi of Hunan; P6: 湖南浏阳Liuyang of Hunan; P7: 湖南新晃Xinhuang of Hunan; P8: 湖南桂阳Guiyang of Hunan; P9: 江西婺源Wuyuan of Jiangxi; P10: 江西铜鼓Tonggu of Jiangxi; P11: 江西分宜Fenyi of Jiangxi; P12: 江西信丰Xinfeng of Jiangxi; P13: 福建南平Nanping of Fujian; P14: 福建明溪Mingxi of Fujian; P15: 福建福州Fuzhou of Fujian; P16: 福建武平Wuping of Fujian; P17: 广东乐昌Lechang of Guangdong; P18: 广东连州Lianzhou of Guangdong.
第1亚类包括湖南浏阳、福建明溪、浙江临安、广东乐昌、浙江丽水、浙江江山和福建南平7个居群;第2亚类包括湖南桂阳、福建福州和浙江龙泉3个居群;第3亚类包括江西铜鼓、广东连州、江西婺源、湖南桑植和江西分宜5个居群;第4亚类包括湖南新晃和江西信丰2个居群;第5亚类仅包含福建武平1个居群。地理位置相邻的居群不能完全聚在一起,但总体上看,第1类中的居群大多位于浙江省和福建省,第2类中的居群大多位于江西省。
3 讨论和结论
上述研究结果显示:南方红豆杉天然居群的叶片表型性状无论是在居群间还是居群内均存在极显著(P<0.01)差异,其叶片氮磷化学计量指标在居群间也存在极显著差异,表明南方红豆杉叶片的表型性状和氮磷化学计量存在丰富的地理变异。福建武平居群的南方红豆杉叶长、叶宽、叶面积和叶周长总体上显著(P<0.05)大于其他居群,这可能与该居群的生境有关。该居群位于福建省梁野山国家级自然保护区内海拔800~1 050 m的区域,土壤类型为山地黄红壤,林地土层、腐殖质层和地表枯枝落叶层较厚,林下土壤自然肥力高,且该区域雨量充沛、水力资源较为丰富,这些立地条件均有利于南方红豆杉叶片生长。南方红豆杉叶片的表型性状和氮磷化学计量指标的变异系数变幅较大(11.13%~29.56%),且以叶宽长比的变异系数最大,表明该性状可塑性最大,易受到个体发育和生境的双重影响。
南方红豆杉叶片表型性状的表型分化系数均值为24.94%,居群内方差分量百分比明显大于居群间,说明居群内变异是其表型性状变异的主要来源。陈洁[26]认为,异花授粉有利于植物居群间基因流动和遗传物质交换,南方红豆杉以风媒传粉、异交为主,种子有红色肉质假种皮,对鸟类、啮齿动物有一定吸引作用,这些繁殖特性推进了其基因流动,从而降低了遗传漂变对居群遗传结构的影响,而频繁的基因交流有助于维持居群内较高的遗传变异水平。南方红豆杉叶片的表型性状在居群间和居群内都存在极显著差异,这些差异是遗传和环境共同作用的结果。
基于居群水平的表型变异分析,在空间尺度上研究特定植物类群的表型性状与地理-气候因子间的关系,往往可以更准确地揭示植物变异格局[27]。通常情况下,经纬度和海拔的梯度变化能够综合反映气温、降水和植被等环境因子的作用[28]。本研究结果显示:南方红豆杉的叶长、叶面积和叶周长在地理空间上呈现以经度为主的单一变异模式,浙江省和福建省内的居群叶片较江西省内的居群叶片短小,这种在北半球随内陆向海洋过渡引起的地理经向变异规律在尾叶樱桃〔Cerasusdielsiana(Schneid.) Yü et Li〕[25]和红椿(ToonaciliateRoem.)[29]等植物种类中也有过报道,通常反映了水分和温度的梯度变化。刘从等[1]认为,植物体通常能够通过改变生物量分配和形态变化来适应环境的改变,即形态可塑性。植株叶片变短小,有利于削弱植物自身的边界阻力,增强叶片边缘对光热和CO2的传导能力,进而降低呼吸和蒸腾的成本,使较小的叶片对生境中光热的变化适应性更强[30]。在南方红豆杉5个叶片表型性状中,叶宽长比与最热月最高温极显著负相关,而与年均温、最冷月最低温、年均降水量、最湿月降水量和最干月降水量均无显著相关性,表明最热月最高温是推动南方红豆杉叶片表型性状变异的主要气候因子,也进一步验证了气温对植株叶片的生长具有较大影响。南方红豆杉各叶片表型性状与纬度均无显著相关性,可能与居群分布地的纬度跨度远小于经度跨度且涉及的叶片表型性状数量较少有关。此外,本研究仅进行了1个年份的采样分析,没有年份间重复采样,因此,需适当增加采样点和表型性状数量并进行年份间的重复研究,以获得更为可靠的研究结果。
除气候因子外,氮、磷元素对植株叶片生长也有较大影响[28]。研究结果表明:在供试的南方红豆杉18个天然居群中,浙江省和福建省内的居群叶片较江西省内的居群叶片短小,且浙江丽水、浙江江山、浙江龙泉、湖南桂阳、江西婺源、福建明溪和福建福州居群的叶片氮磷比均小于14,表明这些居群的植株叶片生长主要受氮元素限制[31]。而湖南浏阳、江西铜鼓、江西分宜、江西信丰和福建武平居群的叶片氮磷比均大于16,表明这些居群的植株叶片生长主要受磷元素限制。
由于地理位置和环境因子的影响,植物会形成连续变异、区域板块变异以及随机变异等多种地理变异模式[29]。聚类分析结果显示:在供试的南方红豆杉18个天然居群中,湖南新晃、江西信丰和福建武平3个居群的叶长、叶宽、叶面积和叶周长均大于其他居群,聚在第2类的第4和第5亚类;而湖南桂阳、福建福州和浙江龙泉3个居群的叶长、叶宽、叶面积和叶周长较小,且他们的地理位置不相邻,聚在第1类的第2亚类,这2类居群基本上都是地理距离较远的间隔居群。湖南浏阳、福建明溪、浙江临安、广东乐昌、浙江丽水、浙江江山和福建南平7个居群的叶片表型性状值均较小,聚在第1类的第1亚类;而江西铜鼓、广东连州、江西婺源、湖南桑植和江西分宜5个居群的叶片表型性状值均较大,聚在第2类的第3亚类,这2类居群中既有地理位置相邻的居群,也有地理不相邻的居群。因而,南方红豆杉叶片表型性状的地理变异既存在区域板块变异也存在随机变异,且大多为随机变异,形成这一现象的原因与居群分布的地理隔离有关,地理隔离导致种子和花粉难以在居群间传播,即使居群间地理距离较近,但基因交流较少;此外,居群的局部小气候复杂多变,环境的异质性也增大了居群间的分化程度。
综上所述,南方红豆杉天然居群叶片的表型性状和氮磷化学计量指标存在明显的地理变异,这些地理变异大多为随机变异。叶长、叶面积和叶周长这3个叶片表型性状在地理空间上呈现以经度为主的单一变异模式。随经度增大叶片趋于短小,叶片氮磷比越大叶片越宽大。叶片表型性状的变异主要来源于居群内,以叶宽长比的可塑性最大,易受到个体发育和生境的双重影响。
致谢:感谢中国林业科学研究院亚热带林业实验中心曾平生高级工程师、姚甲宝高级工程师、厉月桥高级工程师、刘素贞助理工程师和张利利工程师等对本研究提供的帮助!
