丙酸对山羊小肠上皮细胞糖异生途径关键基因表达的影响
2021-03-19宁丽丽霍俊宏占今舜杨天宇赵国琦
宁丽丽 詹 康 霍俊宏 占今舜 彭 程 杨天宇 赵国琦,2,3*
(1.扬州大学 动物科学与技术学院,江苏 扬州 225009;2.扬州大学 农业科技发展研究院,江苏 扬州 225009;3.扬州大学 教育部农业与农产品安全国际合作联合实验室,江苏 扬州 225009;4.江西省农业科学院 畜牧兽医研究所,南昌 330200)
反刍动物体内的碳水化合物主要经过瘤胃微生物发酵后生成短链脂肪酸(Short chain fatty acid, SCFA,包含乙酸、丙酸和丁酸等),再经过糖异生途径转变为葡萄糖被机体吸收利用[1]。糖异生的主要场所是肝脏,肝脏糖异生可产生血液中80%以上的葡萄糖,因此肝脏糖异生对反刍动物十分重要[2]。反刍动物肝脏主要以丙酸、氨基酸(AA)、乳酸和甘油作为糖异生底物合成葡萄糖。丙酸对反刍动物来说更为重要,因为丙酸是反刍动物肝脏糖异生的主要底物,可为糖异生提供60%以上的碳源[3]。有研究发现,向每千克体重的山羊瘤胃灌注1.2 g丙酸后,其血液葡萄糖含量显著升高[4]。奶牛被灌注160.4 g丙酸,可促进犊牛和泌乳中期奶牛肝脏糖异生[5];体外培养的犊牛肝细胞添加2.50 μmol/L丙酸可促进肝细胞糖异生[6]。在反刍动物围产期,肝脏糖异生必须持续快速地运转,一旦不能满足其增加的能量需要,就会出现低血糖和能量负平衡现象,导致代谢紊乱[2,7]。在人和大鼠中小肠被认为是糖异生的另一重要器官,在其饥饿和胰岛素依赖型糖尿病的情况下会出现小肠糖异生现象[8]。小肠糖异生可能在影响血糖调节的门脉系统中起信号转导作用[9]。之前的研究已经发现15%~50%的SCFA可以进入反刍动物消化吸收系统末端[10],这表明部分SCFA可以到达小肠。所以丙酸可能被小肠细胞吸收合成葡萄糖满足对能量的需求。
丙酸可以直接促进牛肝细胞糖异生途径相关基因的表达,包括丙酮酸羧化酶(PC)、细胞质磷酸烯醇丙酮酸羧激酶(PCK1)和线粒体磷酸烯醇丙酮酸羧激酶(PCK2)[6]。丙酮酸羧化酶负责将线粒体中的丙酮酸转化为草酰乙酸,其启动子是通过过氧化物酶体增殖物激活受体γ辅激活因子1α(PGC-1α)的活化而被转录激活[11];PGC-1α可以调节关键的线粒体基因,这些基因有助于促进糖异生途径[12]。细胞质磷酸烯醇丙酮酸羧激酶负责将草酰乙酸转化为磷酸烯醇丙酮酸,这是细胞质糖异生的关键部分。但是草酰乙酸不能直接穿过线粒体进入细胞质,因此PCK2在将草酰乙酸转化为线粒体中的磷酸烯醇丙酮酸中发挥关键作用。丙酸对山羊小肠上皮细胞PCK2mRNA的调控作用尚未被研究。丙酸通过肠脑轴诱导糖异生关键基因的表达,对糖代谢和小肠能量稳态有积极的促进作用[13-14]。由瘤胃微生物产生的SCFA 50%~85%直接通过网胃壁吸收,15%~50%的SCFA不通过网胃吸收,而直接进入牛的小肠吸收系统[10]。由此,推测丙酸可激活山羊小肠糖异生基因的表达,因为已经有研究发现山羊小肠上皮细胞可以运输和吸收丙酸[15],并且丙酸对维持小肠细胞葡萄糖稳态有促进作用[14]。而Zhan等[16]研究表明小肠上皮细胞可以吸收和运输丙酸,并且丙酸可以诱导奶牛小肠上皮细胞糖异生途径关键基因的表达。
之前的研究均集中在反刍动物肝细胞的糖异生途径,而丙酸对山羊小肠上皮细胞(GIEC)糖异生相关限速酶基因表达的影响目前尚不清楚。因此本研究旨在探究丙酸对GIEC中关键糖异生基因mRNA表达的影响,包括线粒体的PCK2、PC、果糖1,6二磷酸酶1(FBP1)和PGC1A,为进一步探究丙酸对小肠上皮细胞糖异生途径的影响提供理论依据。
1 材料方法
1.1 试验材料
DMEM/F12培养基、澳洲胎牛血清、非必需氨基酸(NEAA)、磷酸盐缓冲液(PBS)和胰蛋白酶(Gibco,美国);丙酸、青霉素、链霉素、两性霉素、L-谷氨酰胺溶液和乙二胺四乙酸(EDTA)(Sigma,美国);PrimeScriptTMRT Master Mix和SYBR®Premix Ex TaqTM Ⅱ(TaKaRa,中国);荧光定量96孔板和8连管(Bio-rad,美国);总RNA提取试剂盒(Tiangen,中国)。试验所用的山羊小肠上皮细胞GIEC[15]由扬州大学动物培养物保藏与应用研究所(IACCA)提供。
1.2 试验方法
1.2.1不同浓度丙酸对GIEC糖异生途径相关基因表达的影响
将GIEC(每孔2×105个细胞)接种到6孔板中,培养24 h后进行不同浓度丙酸对GIEC糖异生途径相关基因表达影响的试验。试验分为4个处理组,每个处理3个重复。每个处理组分别添加0、0.75、1.50、和3.00 mmol/L丙酸。放置在37 ℃、5% CO2的培养箱中。孵育6 h后,消化并收集细胞进行RNA提取。
1.2.2丙酸在不同时间点对GIEC糖异生途径相关基因表达的影响
将GIEC(每孔2×105个细胞)接种到6孔板中,培养24 h后进行丙酸在不同时间点对GIEC糖异生途径相关基因表达影响的试验。试验分为2个处理组,每个处理12个重复。每个处理组分别添加0和3.00 mmol/L丙酸。放置在37 ℃、5% CO2的培养箱中。在培养3、6、12和24 h时,每个处理消化并收集3个重复的细胞进行RNA提取。
1.2.3总RNA提取
按照总RNA提取试剂盒(Tiangen,中国)提取总RNA。1%凝胶电泳检测RNA完整性。最后,取 1 μL 提取的样品进行总RNA浓度和纯度的测定。
1.2.4反转录成cDNA
按照Takara反转录试剂盒进行,整个过程在冰上操作。反转录体系为10 μL,反应条件:37 ℃ 15 min 和85 ℃ 5 s。
1.2.5Real-time PCR
荧光定量PCR反应配置总体系为20 μL,其中SYBR®Premix Ex TaqTMⅡKit 10 μL;10 μmol/L的PCR Forward primer和PCR Reverse Primer各0.8 μL;Water PCR grade 6.4 μL;cDNA 2 μL。引物详情见表1。反应条件:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 30 s,40个循环,每个样品都有3个重复。计算方法按照2-ΔΔCt。

表1 荧光定量PCR引物
1.3 数据统计分析
结果采用“平均数±标准差”表示。运用SPSS 16. 0统计软件中的One-Way ANOVA模块进行单因素方差分析,显著性检验应用LSD法。P<0.05 表示差异显著,P<0.01表示极显著差异。
2 结果分析
2.1 不同浓度丙酸对GIEC中糖异生途径关键基因mRNA表达的影响
采用qRT-PCR分别检测GIEC在不同处理中PCK2、PC、FBP1和PGC1AmRNA的表达变化(表2)。与未添加丙酸相比,添加3.00 mmol/L丙酸可显著提高PCK2和PGC1A的mRNA表达量(P<0.05);不同浓度的丙酸对PC和FBP1的mRNA表达量没有显著影响(P>0.05),但是添加3.00 mmol/L丙酸对其表达量有上调的趋势,并且表达量最高。
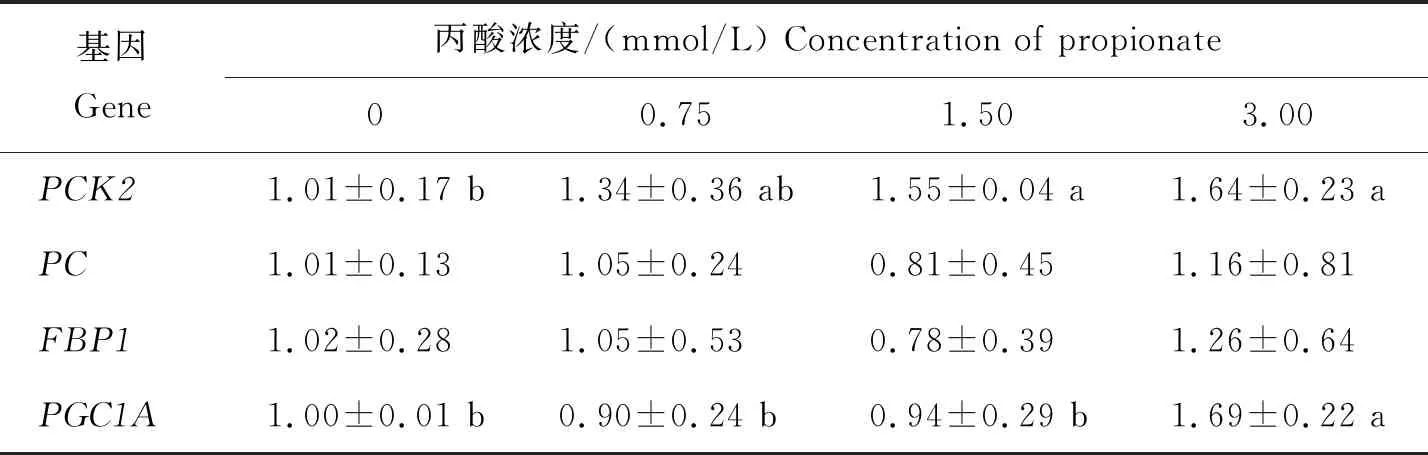
表2 不同浓度丙酸对GIEC中糖异生途径关键基因mRNA表达的影响
2.2 丙酸在不同时间点对GIEC中糖异生途径关键基因mRNA表达的影响
GIEC在不同处理时间的PCK2、PC、FBP1和PGC1AmRNA的表达变化见图1。在孵育3、12和24 h时,与未处理组相比,3.00 mmol/L丙酸对PCK2和PGC1A的mRNA表达量没有显著影响(P>0.05);在孵育6 h时,丙酸极显著增加了PCK2和PGC1A的mRNA表达量(P<0.01),并且表达量最高,且3.00 mmol/L丙酸对PC和FBP1的mRNA表达量无显著影响(P>0.05)。
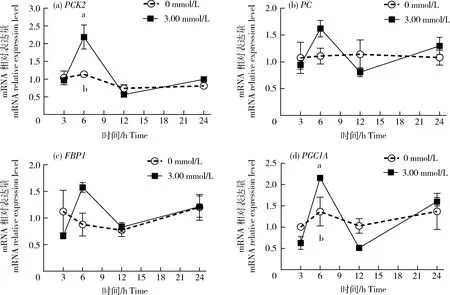
数据标注不同小写字母表示差异显著(P<0.05)。
3 讨 论
糖异生主要有2种途径:一种是使用AA、乳酸、丙酮酸和甘油等经典途径,另一种是使用丙酸作为糖异生底物的途径[2]。在反刍动物中,后者更为重要,因为后者负责将60%~ 74%的丙酸在肝脏中转化为葡萄糖,而丙酸是迄今为止含量最丰富的生成葡萄糖的酸(占瘤胃释放有机酸总量的15%~40%)和糖异生的主要底物[17]。有研究已经证明,SCFA可以被人体回肠吸收,SCFA是肠腔中的主要阴离子,是一种易于吸收和代谢的胃肠道特殊能量来源[18]。此外,15%~50%的SCFA可以进入反刍动物消化吸收系统末端[10],这表明部分SCFA可以到达小肠。反刍动物小肠上皮细胞可以运输和吸收丙酸[15],并且丙酸可以促进牛小肠上皮细胞中糖异生基因的表达[16],这表明丙酸可能诱导山羊小肠细胞中关键糖异生基因的表达。以往的研究主要集中在经典的糖异生途径,但丙酸对山羊小肠上皮细胞糖异生基因表达的影响尚不清楚。因此本试验主要研究丙酸对山羊小肠上皮细胞中糖异生途径关键基因表达的影响。

FBP1的功能是催化果糖1,6-二磷酸水解为果糖6-磷酸。ATP的缺乏会降低FBP1的活性和果糖6-磷酸的产量,最终降低葡萄糖产量[24]。本研究结果显示,与未处理组相比,3.00 mmol/L丙酸处理后FBP1的mRNA表达量提高了23%,并且在 6 h 时表达量最高,这表明丙酸可以诱导山羊小肠上皮细胞产生更多的果糖6-磷酸来合成葡萄糖。本研究还发现,GIEC中添加丙酸后并没有检测到G6PC的mRNA表达,与Zhan等[16]在牛小肠上皮细胞中的研究结果一致。在牛肝细胞中添加丙酸、cAMP或胰岛素也不会改变G6PC的mRNA表达量[6]。PGC1A是一种调节与能量代谢相关基因的转录辅激活因子[25]。此外,PGC1A还是一种重要的蛋白质,可以上调糖异生相关基因的表达,还可以改变PCK启动子的转录活性以调控肝脏中的PCK水平[12]。在本研究中,3.00 mmol/L丙酸处理GIEC显著增加了PGC1A的mRNA表达量,并且依然在6 h时作用效果最好。
4 结 论
丙酸可以在山羊小肠细胞中诱导糖异生途径关键基因PCK2、PC、FBP1和PGC1A的mRNA表达,并且PCK2在GIEC糖异生途径中发挥关键作用,为研究丙酸在诱导糖异生关键基因表达方面提供了新思路。
