不同耐低氮性玉米品种的花后碳氮积累与转运特征
2021-03-18吴雅薇孔凡磊袁继超
吴雅薇 蒲 玮 赵 波 魏 桂 孔凡磊 袁继超
四川农业大学农学院 / 农业部西南作物生理生态与耕作实验室 / 作物生理生态及栽培四川省重点实验室, 四川成都 611130
玉米是中国第一大粮食作物, 其产量直接关系到我国粮食供应的安全[1]。我国耕地面积有限, 其种植面积难以再大幅增加, 因此提高单产成为主要目标, 而施用化肥特别是氮肥是提高玉米单产的主要途径[2]。在我国实际生产中, 农户通常通过投入大量的氮肥来获取作物高产, 导致氮肥利用率(nitrogen use efficiency, NUE)较低, 仅为30%左右, 远低于欧美等发达国家[3-4]。而且, 随着氮肥使用量的增加,各种环境问题(土壤酸化、水土污染等)也随之而来,这要求我们协调好玉米增产和NUE之间的关系。选育氮高效型玉米品种是提高氮肥利用效率、确保玉米增产的同时减少环境污染的有效途径之一[5-6]。
作物的氮高效主要分 3种: 一是作物在低氮素水平下仍具有维持正常生长的能力, 并获得与正常氮时相近的产量, 即低氮高效(耐低氮)型品种; 二是随氮浓度增加, 植物吸收氮素养分并获得较高产量的潜力, 即高氮高效型品种; 三是双高效型[7-8]。通常氮效率表示的是氮素投入与获得产出的比例,研究目的是投入氮素不变或增施氮素能获得更高的产出; 作物耐低氮性的研究目的是减少氮素的投入能获得与正常施氮产出相近或降低较少[9]。基于人们已基本满足温饱, 急需解决环境污染和降低生产成本, 对作物的耐低氮性研究已成为重点。
玉米生育后期(吐丝后)的物质积累与转运是籽粒产量形成的物质基础, 籽粒中的氮素主要通过根系吸收和营养器官的转运[10], 碳水化合物主要来源于花后的光合生产和花前储存在营养器官中的碳水化合物[11]。非结构性碳水化合物(non-structural carbohydrate, NSC, 可溶性糖和淀粉的总和)是作物光合作用的产物, 其在营养器官的贮藏量反映了植株碳吸收(光合作用)与消耗间的平衡[12-13]。氮肥是影响作物碳、氮物质积累和转运的重要因素, 适宜的氮肥用量使玉米在生育后期营养器官中氮素向籽粒的转移和光合产物的积累增加, 进而提高了籽粒产量[14]。前人研究表明, 玉米品种生育后期的物质积累和转运存在基因型差异[7,15]。温立玉等[16]研究表明, 氮高效玉米品种‘鲁单818’具有较氮低效品种更高的花前氮转移效率和花后氮吸收能力。针对不同氮转运效率玉米品种, 陈延玲[17]发现‘先玉335’较‘郑单958’在同等氮水平下有相同的产量, 但是却有较高的籽粒氮浓度, 主要因为它具有较高的营养器官氮素转运效率以及成熟期较低的茎秆氮素残留。苌建峰等[14]研究表明, 碳氮代谢协调性较好的品种在碳、氮的转运过程中, 能很好地协调光合产物和氮素在籽粒灌浆和维持营养器官功能需求之间的矛盾, 尤其是低氮水平下。作为我国玉米三大主产区, 与北方地区不同, 西南丘陵玉米产区坡地比重大、土地贫瘠, 实际生产中, 农民常规施纯氮量高达300 kg hm-2, 且川中丘陵常年阴雨寡照, 昼夜温差小, 玉米干物质转运、积累能力较北方更小[18-19]。有关氮肥水平对玉米干物质和氮素的积累与转运的影响前人已有大量报道[20-21], 不同玉米品种生育后期碳、氮代谢物质积累和转运对氮肥水平响应差异及其对产量贡献程度也已见研究, 但多集中于北方地区[22-23], 鲜见针对四川丘陵地区玉米品种耐低氮能力与碳氮积累转运关系的研究。
本研究以不同耐低氮性玉米品种为材料, 通过分析氮肥用量对不同耐低氮性玉米品种生育后期碳、氮物质积累和转运效率对氮肥水平响应的差异及其对籽粒产量的影响, 探明川中丘区与不同品种相匹配的适宜施氮量和增产提效潜力, 为充分发挥本区低氮高效型玉米品种的减氮提效作用, 实现产量和NUE的协同提高提供理论依据。
1 材料与方法
1.1 试验地概况
于2017—2019年, 在四川省德阳市中江县合兴乡新建村进行(30°38′59″N, 104°53′36″E)。该地区属于亚热带季风性湿润气候, 气象数据如图1, 3年生育后期均无补充灌溉。2018年7月大量降雨, 极大影响了玉米吐丝至灌浆阶段的生长活动, 进而导致本试验年际间的差异。供试土壤为紫色壤土。玉米播前0~20 cm土壤基础地力数据如表1。由于2017—2019年玉米前茬作物不同(2017: 蔬菜; 2018: 羊肚菌; 2019: 小麦), 导致3年地力存在差异。供试材料为前期筛选出的耐低氮品种‘正红 311’(Zhenghong 311, ZH311)和低氮敏感品种‘先玉 508’(Xianyu 508, XY508)[6], 2个品种生育期均为120 d左右。
1.2 试验设计
本试验为 3年定位试验, 采用二因素随机区组试验设计, A因素为不同耐低氮性玉米品种‘正红311’和‘先玉 508’, B 因素为氮肥(纯氮)用量, 所用氮肥为尿素, 设B0 (0 kg hm-2)、B1 (150 kg hm-2)、B2(300 kg hm-2)和B3 (450 kg hm-2) 4个氮肥处理。共8个处理, 每处理 3次重复, 共 24个小区, 小区面积6.0 m×7.5 m。
3月下旬覆膜直播, 单株栽培, 宽窄行种植, 行距为1.5 m + 0.5 m, 密度52,500株 hm-2; 50%的氮肥做底肥, 50%在大口期追施; 磷钾肥采用大田生产常规施用量, 过磷酸钙600 kg hm-2, 氯化钾150 kg hm-2, 一次性基施, 其他管理措施同当地生产实际, 7月下旬收获。
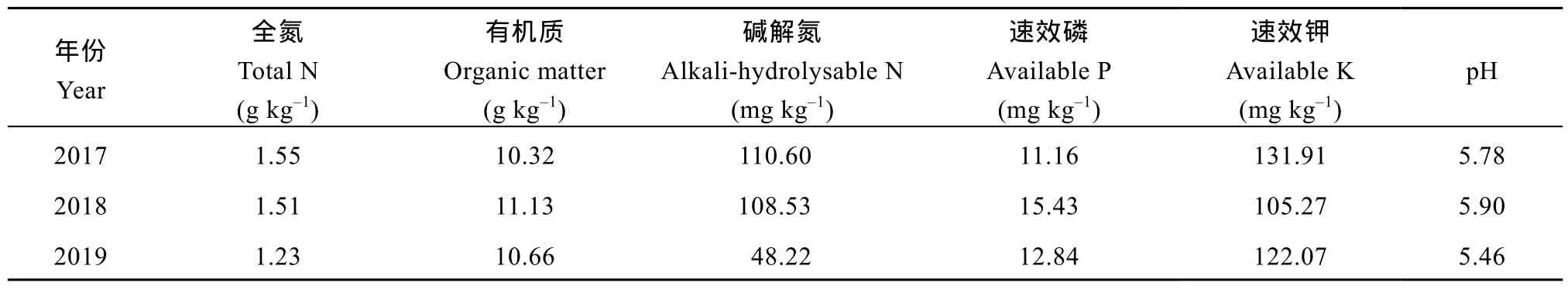
表1 2017-2019年玉米播前0~20 cm土层基础地力Table 1 Foundation soil capacity of 0-20 cm soil layer before maize sowing date from 2017 to 2019
1.3 测定指标与方法
1.3.1 干物质积累与转运量测定 2017—2019年, 在吐丝盛期, 选择长势和吐丝期基本一致的植株挂牌标记。由于玉米的茎叶干重仍会在吐丝期后有一定增长, 直到吐丝7 d后才发生物质转运, 因此于吐丝后 7 d和成熟期, 每个小区分别选取挂牌植株5株, 将植株地上部分为茎鞘、叶片和穗(成熟期分为籽粒、苞叶和穗轴), 在105℃下杀青30 min后75℃烘至恒重后称重。用高速粉碎机将样品全株粉碎备用, 用作全氮、可溶性糖和淀粉的测定。并计算以下指标[24]:
1.3.2 氮素积累与转运测定 2018—2019年, 取样方式同1.3.1, 采用凯氏定氮法[25]测定各器官全氮含量。并计算以下指标[26-27]:
1.3.3 非结构性碳水化合物积累与转运测定2018—2019年, 取样方式同 1.3.1, 采用蒽酮-硫酸比色法[28]测定可溶性糖、淀粉含量, 可溶性糖和淀粉含量即为非结构性碳水化合物(NSC)含量, 各器官 NSC积累量 = 各器官干物质重量×各器官NSC百分含量。并计算花前 NSC转运量(NSC translocation amount of pre-silking, NSCRA)和转运率(NSC translocation rate of pre-silking, NSCRR)、花前 NSC转运量对籽粒贡献率(contribution rate of NSC translocation amount of pre-silking, CNSCRA)以及收获指数(NSC harvest index, NSCHI), 公式同1.3.2氮素相关指标计算。
1.3.4 产量及产量构成因素 于成熟期每小区随机连续取20穗, 调查百粒重。各小区实收计产, 折算成 14%含水量的单位公顷产量, 以实测产量除以穗数和百粒重计算穗粒数。
1.4 数据处理与统计
采用Microsoft Excel 2019和DPS 7.05数据分析软件整理与分析数据, 表2以三因素随机区组程序进行方差分析, 其余图和表以二因素随机区组程序(氮肥×品种) LSD法进行差异显著性检验。采用Graphpad Prism 7和Microsoft Excel 2019软件作图。
2 结果与分析
2.1 关键指标方差分析结果
方差分析表明(表2), 种植年份、玉米品种和氮肥水平及三者的互作效应显著影响玉米的籽粒产量,花前干物质、碳氮转运量及花后干物质、碳氮积累量。其中, 在品种和氮肥的互作效应上, 籽粒产量以及花前干物质、氮素和NSC转运量达到极显著水平;花后干物质、氮素和NSC积累量达显著水平, 二者的互作效应对物质转运的影响较物质积累更大。
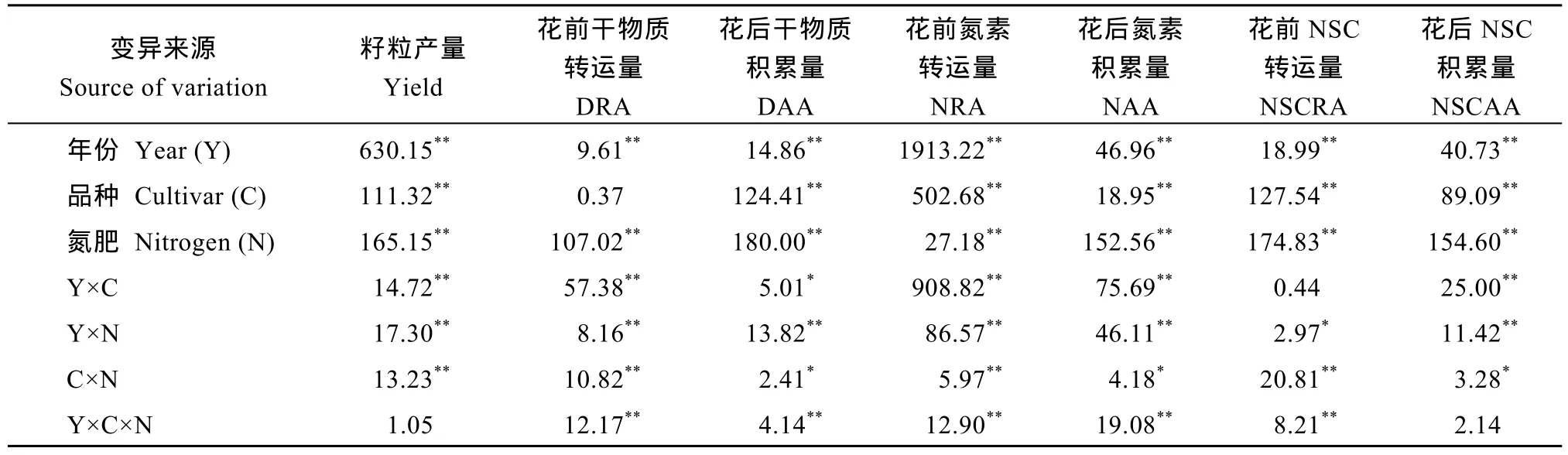
表2 年份、品种与氮肥水平对玉米产量和碳氮物质转运、积累量影响的方差分析Table 2 Variance analysis of year, cultivar, and nitrogen application on grain yield, nitrogen and carbon translocation, and accumulation amount in maize
2.2 不同施肥水平下玉米产量的品种间差异
图2结果表明, 不同玉米品种间产量及其构成因素对氮肥水平的响应存在差异。总的来说, 正红311籽粒产量, 尤其在低氮条件下(≤150 kg hm-2)较先玉508更具优势。2017—2019年, 0 kg hm-2氮处理下, 正红311籽粒产量较先玉508分别高31.98%、26.40%和14.02%; 150 kg hm-2氮处理下, 正红311籽粒产量较先玉 508分别高 27.06%、16.94%和24.10%。除2017年外, 正红311和先玉508籽粒产量在中高氮(300~450 kg hm-2)差异不显著。低氮条件下, 品种间籽粒产量上的差异受各产量构成因素的影响程度不同。除2017年外, 穗粒数在正红311和先玉 508上的差异不显著; 而正红 311的有效穗数(除2019年)和百粒重(除2018年)在低氮条件下显著高于先玉508。正红311各产量构成因素在低氮处理下要优于先玉 508, 但造成产量差异的主要因素受年际间影响。
2.3 不同施肥水平下玉米干物质积累和转运的品种间差异
氮肥用量对玉米植株干物质积累量(生物量)影响显著(图3), 但影响的程度在品种间存在一定差异。随施氮量增加, 先玉 508的干物质积累总量不断增加, 正红 311则先增后略降。各氮处理下正红311的干物质量均显著大于先玉508。从2个生育时期(吐丝后 7 d和成熟期)干物质量平均结果来看,2017年正红311植株在0、150、300和450 kg hm-2处理下的干物质积累量分别较先玉508高29.59%、19.54%、35.44%和18.16%, 2018年分别高57.19%、30.03%、39.03%和45.91%; 2019年分别高26.86%、27.81%、30.46%和17.20%。
表3结果表明, 增施氮肥显著降低花前干物质转运量(DRA)、转运率(DRR)及其对籽粒重的贡献率(CDRA)和收获指数(HI), 提高植株花后物质积累量(DAA) 及其对籽粒重的贡献率, DAA对籽粒贡献率大于 DRA。低氮处理(0~150 kg hm-2), 正红 311的DRA和CDRA较先玉508更高或接近, 中高氮(300~450 kg hm-2)下则相反, 其DRA和CDRA较先玉508更低, 尤其 2019年, 正红 311在 300 kg hm-2和450 kg hm-2干物质没有转运。
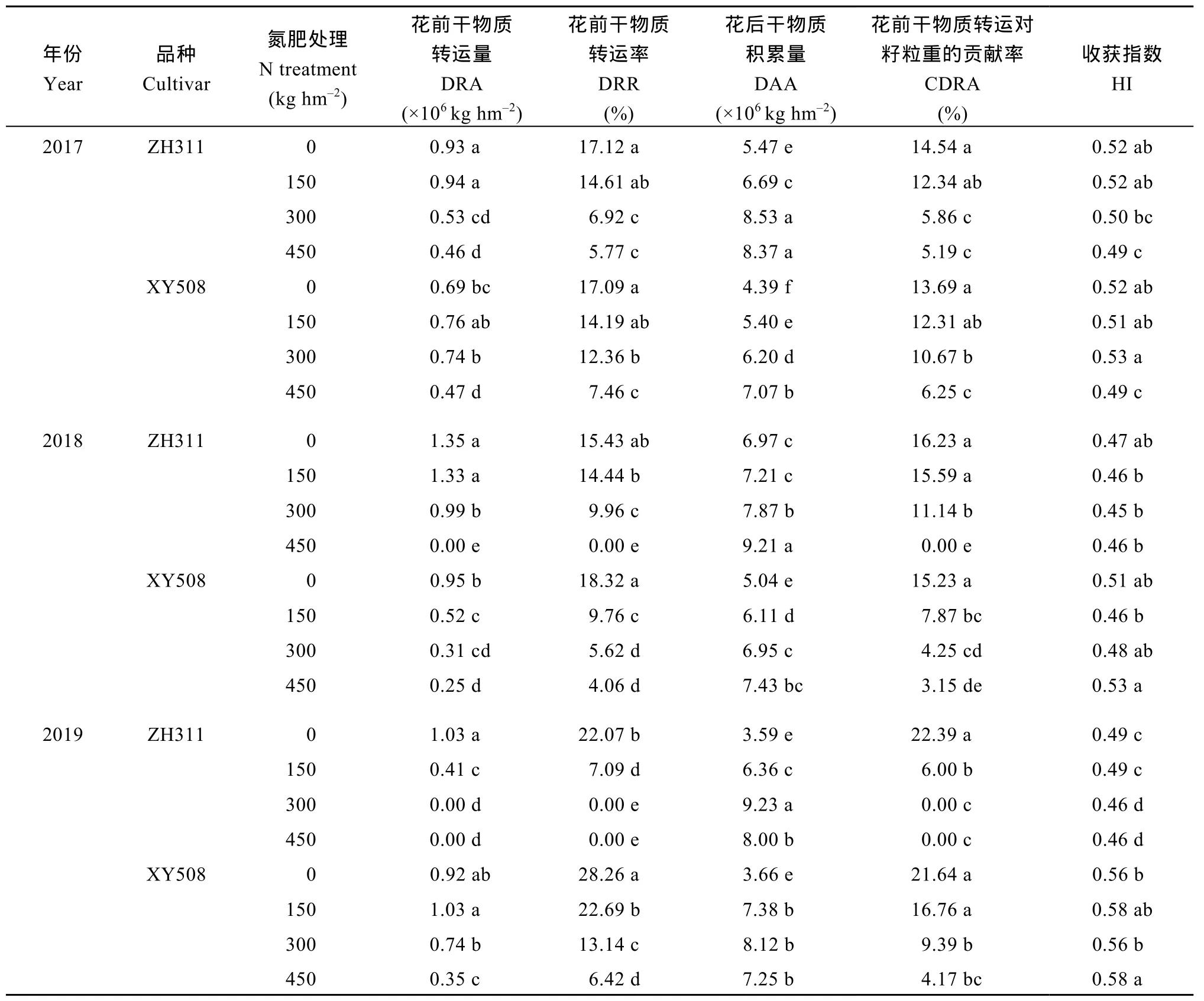
表3 氮肥水平对不同耐低氮性玉米品种吐丝后干物质转运的影响Table 3 Effects of N level on dry matter translocation of maize with different low N tolerance after silking stage
2.4 不同施肥水平下玉米非结构性碳水化合物积累和转运的品种间差异
由图4可知, 各氮处理下, 正红311的NSC含量均显著大于先玉508。从2个生育时期NSC积累量平均结果来看, 2018年正红 311在 0、150、300和450 kg hm-2处理下, 植株NSC较先玉508分别高56.59%、25.46%、31.61%和24.31%; 2019年较先玉508分别高22.38%、17.20%、25.74%和1.54%。品种间2年的差异均在0 kg hm-2处理下最大, 2018年的差异大于2019年。
由表4可知, 与干物质转运相似, 增施氮肥显著降低吐丝后植株 NSC转运量(NSCRA)、转运率(NSCRR)及其对籽粒 NSC (CNSCRA)的贡献率和NSC收获指数(NSCHI), 提高植株花后NSC积累量(NSCAA)及其对籽粒 NSC的贡献率。正红 311的NSCRA、NSCRR和CNSCRA较先玉508更低。2018年, 正红311 NSCRR在0~450 kg hm-2氮处理下分别较先玉508低19.63%、46.58%、88.85%和100.00%,2019年分别低33.54%、66.02%、100.00%和100.00%;正红311的CNSCRA在0~450 kg hm-2氮处理下较先玉508分别低(2018年) 29.44%、45.24%、92.18%和100.00%, 2019年分别低32.20%、60.99%、100.00%和100.00%, 二者差异随施氮量的减少而减小。与花前NSC转运相反, 正红311的NSCAA及其对籽粒NSC的贡献较先玉 508高, 二者差距随施氮量的减少增大。
2.5 不同施肥水平下玉米氮素积累和转运的品种间差异
氮水平对玉米氮积累量同样存在品种差异(图5)。各氮处理下, 正红311的氮积累量均显著大于先玉 508。从 2个生育时期氮素积累量平均结果来看, 正红311植株在0、150、300和450 kg hm-2处理下, 氮积累量在2018年较先玉508高44.16%、32.18%、49.22%和 43.67%, 2019年高 10.30%、27.51%、58.84%和 23.09%, 2018年的差异较 2019年大。花前和花后氮素积累量在成熟期植株氮素积累量的占比存在年际差异, 2018年, 正红 311在0~450 kg hm-2氮肥水平下花前氮素积累量分别占成熟期氮素积累量的 79.64%、78.02%、82.51%和67.12%, 2019年花前氮素积累量分别占 69.28%、46.06%、40.97%和50.93%; 2018年, 先玉508在各氮水平下花前氮素积累量分别占成熟期氮素积累量的71.74%、59.78%、58.70%和58.47%, 而2019年分别占 64.32%、51.03%、51.97%和 49.85%。总的来说, 2018年花前氮素积累率较 2019年大, 正红311花前氮素积累率较先玉508高。
如表5所示, 随着施氮量增加, 植株的氮素转运率(NRR)及其对籽粒氮素(CNRA)呈下降趋势, 花后氮素积累量(NAA)及其对籽粒氮素的贡献率呈增加趋势, 高氮会降低氮收获指数(NHI)。各指标年际差异较大, 品种间的氮素转运、积累对氮水平响应程度也不同。2018年, 正红 311的 NRA、NRR和CNRA均显著大于先玉508, NAA及其对籽粒氮素的贡献则更小。正红311的NRA在0~450 kg hm-2处理下分别较先玉508高90.55%、119.20%、150.88%和130.66%; NRR分别高25.16%、40.69%、43.61%和52.69%; CNRA则分别高24.46%、74.34%、87.41%和54.68%。正红311的NAA在0~450 kg hm-2处理下较先玉 508分别低-0.77%、31.98%、33.12%和-11.50%。与2018年不同, 2019年, 正红311的NRA、NRR和CNRA较先玉508更小。0~450 kg hm-2处理下, 正红 311的 NRA较先玉 508分别低 14.50%、16.09%、52.09%和 22.62%; NRR分别低 22.29%、30.23%、66.84%和41.89%; CNRA分别低18.94%、31.30%、66.91%和31.11%。2个品种间NAA及其对籽粒氮素贡献的差异2年表现相同规律。0~450 kg hm-2氮处理下, 正红311的NAA较先玉508分别高32.53%、41.20%、73.99%和23.20%。总的来说, 正红311花后氮素的积累, 尤其是低氮条件下(≤150 kg hm-2), 比先玉508的大。
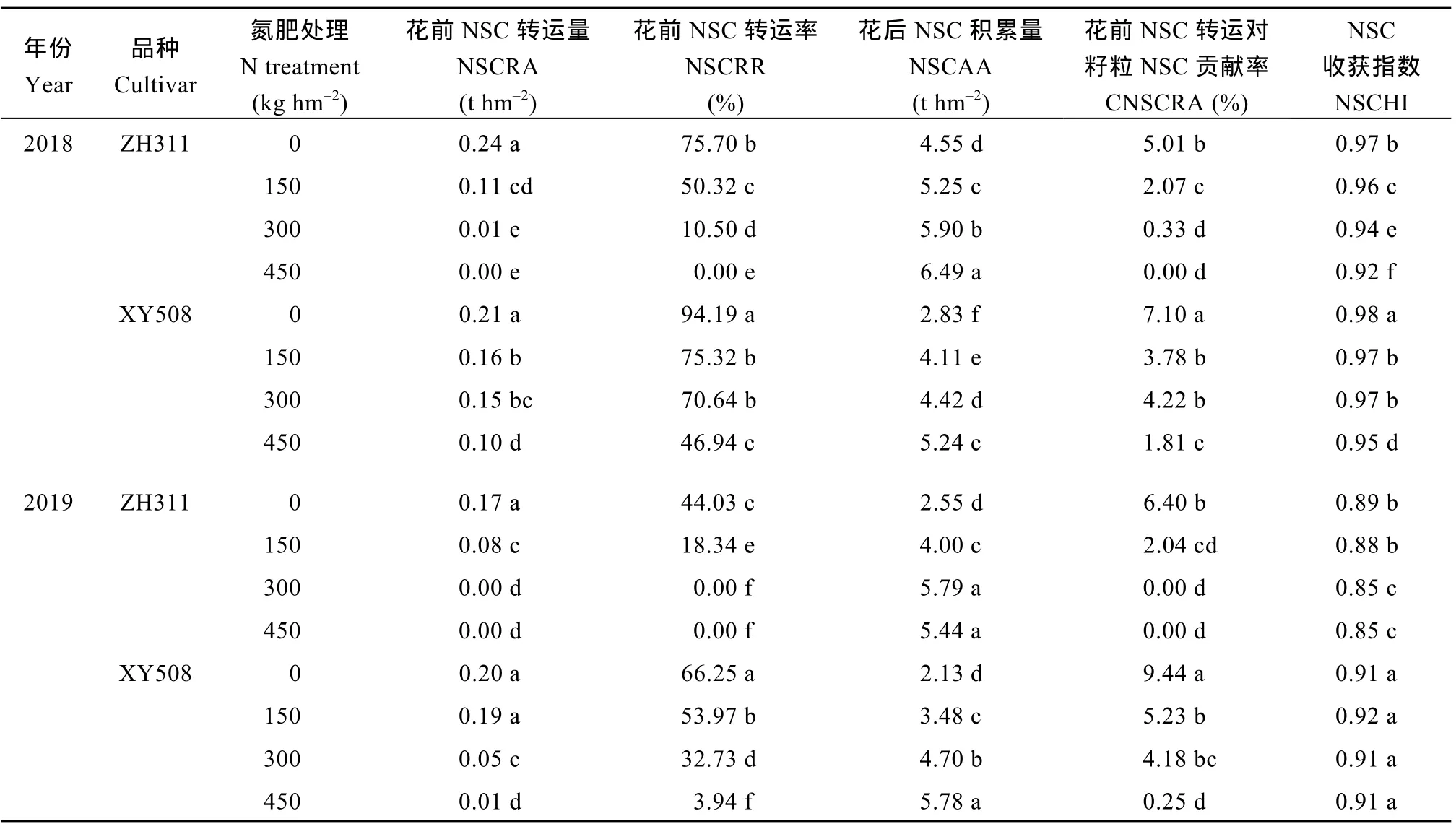
表4 氮肥水平对不同耐低氮性玉米吐丝后非结构性碳水化合物转运的影响Table 4 Effects of N level on non-structural carbohydrate (NSC) translocation of maize with different low N tolerance after silking stage
2.6 物质积累、转运与籽粒产量的关系
不同品种物质积累、转运与籽粒产量的关系见表6。对正红311而言, 其籽粒产量、有效穗数与花后干物质、NSC积累量关系密切, 其关系均达显著或极显著水平; 穗粒数和氮素转运量与NSC积累量密切相关; 百粒重的形成则与干物质和NSC的转运以及氮的积累量有显著关系; 干物质的花前转运量与氮积累量和 NSC转运量相关系数达显著、极显著水平,而花后干物质积累量与NSC的积累极显著正相关。
与正红 311不同的是, 先玉 508的产量除与花后干物质、NSC积累量密切相关外, 还与花前干物质和 NSC的转运量密切相关; 有效穗数受氮素转运、积累量的影响也达到显著水平; 穗粒数与干物质转运呈显著负相关, 和其他指标关系不大; 百粒重除了和干物质转运量关系不大, 和各其他物质、转运积累量均显著、极显著正相关; 干物质转运量与氮素的积累、转运及 NSC的转运的关系不密切,与其余物质积累量呈显著或极显著负相关, 干物质积累量与NSC的转运呈负显著水平; 氮素转运和积累极显著正相关; NSC的转运量与其积累量呈显著负相关。
3 讨论
3.1 增施氮肥促进花后碳氮物质积累, 降低花前物质转运效率
作物养分吸收、积累是干物质形成和累积的基础, 也是产量形成的前提[29]。在生产成本渐高及环境污染日益严重的形势下, 如何在保证产量的同时, 进行氮肥优化管理, 实现较高的氮肥利用率,是玉米生产中一个亟待解决的问题[4,30]。众多研究表明[27,31], 在受到缺氮胁迫时, 营养器官中氮、碳向籽粒转移的比例明显增加。缺氮胁迫时, 土壤中的氮无法满足籽粒灌浆需要, 茎叶中的氮素转移量便增加, 如果氮转移过多, 则会削弱光合活性, 减少同化物合成[10,32]。吐丝前的光合产物在营养器官中以非结构性碳水化合物的形式贮存[5,33], 在花后作为重要的“源”, 为籽粒灌浆提供物质基础[34]。
本研究表明, 增施氮肥使玉米植株的干物质、氮素积累量增多; 花前干物质和NSC的转运量、转运率及其对籽粒的贡献率随施氮量显著降低, 花后积累量及其对籽粒的贡献率则随氮肥施用量增加;在土壤供氮不足时, 氮素的转移量和比例较氮素充足(过量)供应时也更大, 这与前人结果相似[22,24]。无论是籽粒干重还是 NSC, 均以花后物质积累对籽粒贡献更大, 且花后贡献率随氮肥增施逐渐增高。尽管低氮环境下, 玉米植株有较高的花前碳水化合物积累量及氮转运量以弥补产量损失, 但合理的氮肥用量能够维持植株碳氮代谢的平衡, 进而影响玉米生育后期营养器官碳、氮素的转运及光合产物的积累, 促进碳、氮同化物向籽粒的供应, 从而提高籽粒产量[22,35]。
3.2 不同耐低氮性玉米品种生育后期干物质积累与转运特性存在一定差异
大量研究表明, 同一作物不同品种间的耐低氮能力和氮效率存在一定差异[5-7]。作物的耐低氮能力,一般用低氮与正常氮评价指标的相对值表示, 是品种之间指标相对值的比较, 据此可以分为耐低氮型、中间型、低氮敏感型3类[8,36]。我们在前期研究中发现, 耐低氮品种正红 311较低氮敏感品种先玉508在中低氮条件下有较强的氮素吸收、积累速率等氮代谢能力[37-38], 以及较高的光合生产与干物质积累能力[36]。在本研究中, 这一差异也引起了品种间生育后期营养器官干物质和碳水化合物积累、转运量及其对籽粒的贡献率对氮肥水平的响应的不同。

表5 氮肥水平对不同耐低氮性玉米品种氮素转运的影响Table 5 Effects of N level on N translocation of maize with different low N tolerance after silking
花前的物质积累是提高花后光合生产的基础,而花后的光合生产能力决定了籽粒产量[39]。耐低氮性品种正红 311生育后期植株干物质和NSC积累量均显著高于氮低效品种先玉 508, 且正红311在中低氮水平下(0~300 kg hm-2)的优势较大。本研究表明,0~150 kg hm-2施氮量下, 正红311干物质和NSC转运量及积累量较先玉508更高, 但二者转运率差别不大。同时, 正红 311在低氮条件下花后干物质和 NSC的积累对籽粒干重以及碳水化合物的贡献程度较高,但花前转运量对籽粒贡献率不具优势。产量构成因素在产量形成中的作用与品种特性和环境条件有关[40]。本研究中, 正红311植株干物质在花后的积累能够促进玉米显著促进有效穗数增加, 进而提升产量;而先玉 508的百粒重受干物质积累的影响更明显, 过多的物质转运反而不利于产量构成因素的建成。与先玉508相比, 正红311, 尤其在低氮条件下, 保持茎叶花前积累干物质向籽粒更合理(低)的转移比例,以维持花后茎叶更高的光合物质生产(积累)量, 保证植株穗部正常生长发育, 进而保证籽粒产量, 这可能是正红311耐低氮的生理机制之一。
3.3 不同耐低氮性玉米品种生育后期氮素积累与转运特性存在一定差异
米国华等[7]认为玉米氮高效品种应该在生育后期, 充分利用前期高效吸收的氮, 供给籽粒生长,从而减少叶片中氮素的输出, 保证物质生产, 为籽粒灌浆提供碳水化合物。郭松等[41]认为最优的氮高效品种要在花后吸氮能力强的基础上, 尽可能增加营养器官中氮素向籽粒的转运, 从而保证产量提高的同时, 籽粒含氮量得到稳定。本研究表明, 2个品种氮素转运量、转运率及其对籽粒的贡献程度存在较大的年际差异, 这可能与降雨量和土壤肥力有关。2018年降雨量(1374.1 mm)较2019年(776.2 mm)高近2倍, 尤其是吐丝至灌浆期。大量的降水可能导致氮素等养分流失, 根系对土壤氮素吸收量降低[13,27]。2018年土壤含氮量较2019年高, 表明在土壤肥力较高时(2018年), 正红 311利用其强大的氮素吸收能力[36-37], 在花前能积累较多氮素, 贮藏于茎叶中的氮素向籽粒的转运量及其贡献率也就较高; 而在土壤肥力较低时(2019年), 由于花前吸收积累的氮素相对较少, 花后积累的氮素及其对籽粒的贡献率就相对较高; 先玉508两年花前氮素转运和花后氮素积累对籽粒的贡献率变化较小, 氮素花后积累对籽粒的贡献率均较高, 因此增施氮肥延缓根系衰老, 增加花后氮素积累对增加其籽粒氮积累显得尤其重要。与正红 311不同, 先玉 508籽粒产量及其构成因素受氮素转运量、转运率的影响较大。适宜的氮素转移、积累的能够为营造玉米吐丝前后良好的营养条件通过提高有效穗数和穗粒数, 提高结实率,减少籽粒败育, 利于高产[42]。
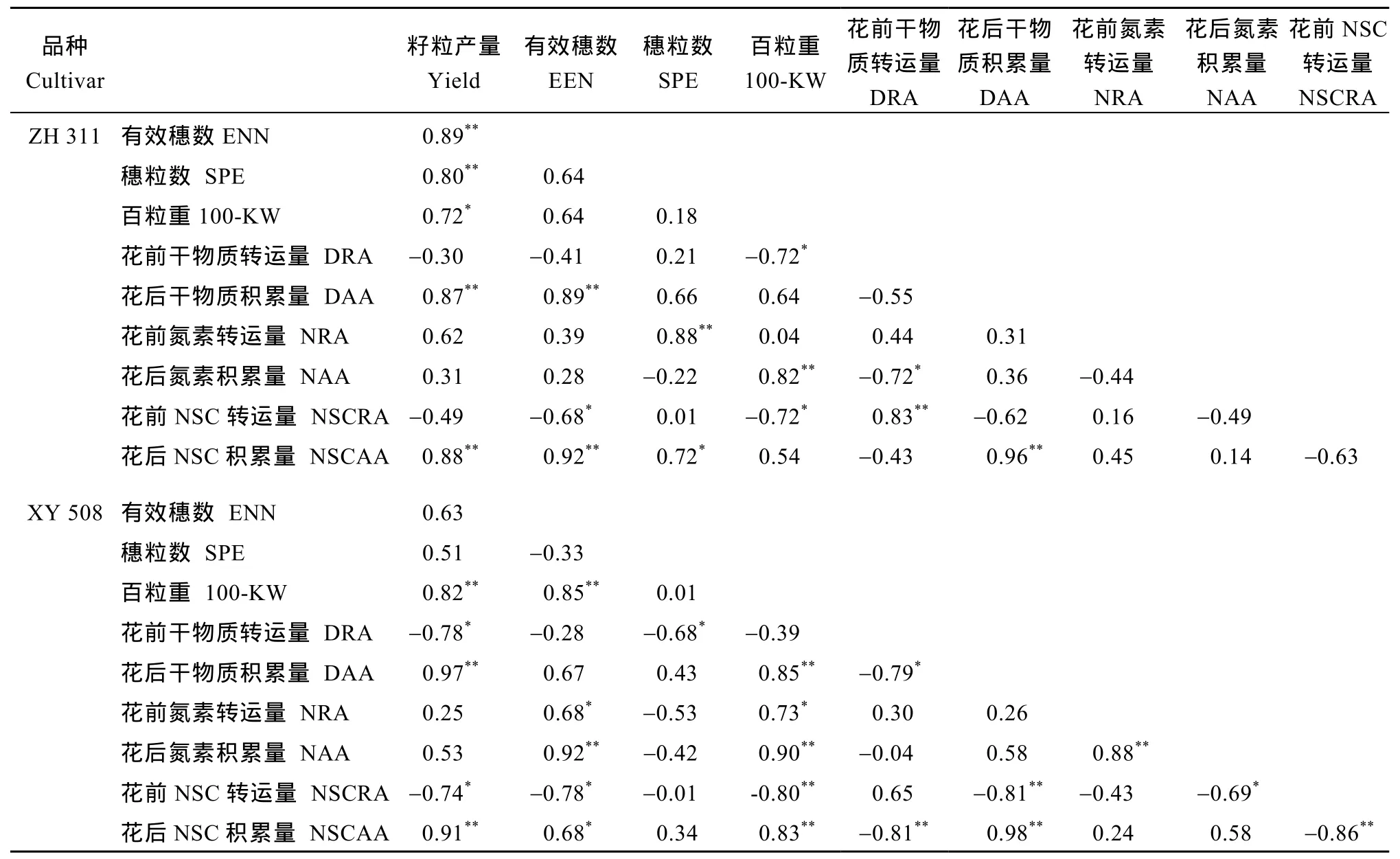
表6 籽粒产量与花前干物质、氮素和非结构性碳水化合物转运量及花后积累量的关系Table 6 Relationship between grain yield and pre-silking translocation and post-silking accumulation of dry matter, nitrogen, and non-structural carbohydrates
综上所述, 正红311在中低氮条件下, 花前干物质和氮素的转运率较先玉508差异不大, 但其花后积累能力较强, 表明正红311能够在花后保持较强的代谢和光合生产能力以保证充足的碳水化合物供应。供氮不足时, 正红311营养器官较高的物质积累量为籽粒灌浆提供更充足的花前物质基础, 同时具有更强的氮素积累(吸收)能力, 在一定程度上保证了花后光合产物的合成[20,43]。面对低氮胁迫, 正红 311在保证花前贮存物质充分向籽粒运输的同时, 维持着较高的同化物积累能力, 以弥补缺氮造成的产量损失。值得注意的是, 尽管 2个品种的籽粒产量随施氮量增加而提高, 但收获指数随施氮量减小, 且增施氮肥对先玉508产量的促进更大。正红311的产量优势随施氮量的增加而减小, 在450 kg hm-2施氮量下其产量较300 kg hm-2降低了1.24%。在实际生产中, 对耐低氮品种进行种适度减氮以促进花前物质向籽粒的转运, 提高收获指数, 避免植株徒长旺长和资源的浪费; 对低氮敏感品种, 可适当增施氮肥以激发其产量潜力。
4 结论
供氮不足时, 玉米通过增加花前贮存碳、氮物质对籽粒的转运以保证产量。增施氮肥能够提高玉米花后干物质和碳水化合物生产及其对籽粒质量的贡献率。与低氮敏感品种先玉 508相比, 耐低氮品种正红311在低氮条件下, 正红311在花前、花后均保持较高的植株干物质、NSC和氮素积累量, 花后物质积累量及其对籽粒质量的贡献率更高, 从而拥有更高的籽粒产量; 品种间花前物质转运率及其对籽粒质量的贡献率差异不大。面对低氮胁迫, 正红311既保证了花前贮存物质充分向籽粒运输, 同时维持着较高的同化物积累能力来影响其产量形成。
