籼型杂交稻恢复系动态株型与光能利用率评价
2021-03-17王丰廖亦龙柳武革刘迪林曾学勤傅友强朱满山李金华付崇允马晓智霍兴
王丰 廖亦龙 柳武革 刘迪林 曾学勤 傅友强 朱满山 李金华 付崇允 马晓智 霍兴
籼型杂交稻恢复系动态株型与光能利用率评价
王丰#, *廖亦龙#柳武革 刘迪林 曾学勤 傅友强 朱满山 李金华 付崇允 马晓智 霍兴
(广东省农业科学院 水稻研究所/广东省水稻新技术育种重点实验室,广州 510640;#共同第一作者;*通信联系人,E-mail: fwang1631@163.com)
【】理想株型是水稻高产稳产的前提。本研究拟对不同年代育成的籼型三系杂交稻恢复系动态株型与光能利用效率进行综合评价,探讨恢复系光能高效利用的理想动态株型特征,为提高光能高效利用的高产稳产杂交稻育种效率提供理论依据。以明恢63、明恢82、蜀恢527、广恢998、广恢122、广恢128、广恢308和桂99等8个大面积应用的籼型杂交稻恢复系为供试材料,在华南生态条件下考查了其分蘖、叶面积指数(LAI)、透光率、叶片大小与开张角、叶色(SPAD值)等性状的动态变化,以及抽穗期光合速率、生育期、产量相关性状和光能利用率(RUE)等指标。各恢复系之间在株型动态变化、产量和光能利用率(RUE)等方面存在较大差异。良好的动态株型与光能利用率密切相关,最为接近华南生态条件下理想动态株型的广恢998和广恢308,光能利用率最高。根据聚类分析,广恢998、广恢308和明恢82属于光能高效利用的类群Ⅰ。其他恢复系归为类群Ⅱ。最后,根据华南的光温生态特点,提出了水稻光能高效利用的理想动态株型模式及育种选择关键指标:1)移栽后分蘖和叶面积发展快,LAI达到1.0的时间≤27 d;茎蘖数达到峰值所需时间≤38 d,分蘖力中等,最高茎蘖数≤600苗/m2;2)前期窄叶长披,移栽后15 d顶部3片完全叶平均长度为37 cm,平均宽度≤0.9 cm,开张角度≥80°;后期叶片窄直,抽穗后20 d顶部倒3叶平均开张角≤65°,平均长度为43 cm,平均宽度≤1.3 cm,属于中叶型,有利于后期群体通风透光;3)后期叶片转色好,不早衰,SPAD值维持在35~40,抽穗前20 d至抽穗后20 d之间的叶片SPAD值降幅≤13%;普通栽培条件下平均每穗总粒数180粒左右,有效穗数300穗/m2以上,结实率≥85%。广恢998和广恢308具有较为理想的动态株型和较高RUE,利用其配制的杂交稻天优998、博优998和五优308等连续多年成为我国华南稻区主导品种和国家与省级区试对照品种。说明通过选育光能高效利用的理想动态株型恢复系,是组配选育高产稳产杂交稻行之有效的重要途径。
籼型杂交水稻; 恢复系; 动态株型; 光能高效利用; 双季稻区; 华南
水稻是世界50%以上人口的主食。不断提高水稻产量潜力,满足人们日益增长的刚性需求,成为育种家们孜孜以求的奋斗目标。众所周知,稻谷产量主要是由水稻植株捕获太阳光能后经叶片光合作用将水和CO2转化成氧气和葡萄糖,进而合成淀粉等化合物累积在籽粒中而形成。产量取决于稻株群体捕获吸收的光能以及将所捕获光能转化成化学能的转化效率[1-2]。提高水稻全生育期群体光能捕获能力和光能转化效率,即提高水稻的整体光能利用率,成为提高水稻产量潜力极为重要的技术途径。理想的动态株型是影响光能捕获效率和把所截获光能有效地分配到全部叶片中的最主要因素之一[3]。为此,通过塑造良好株型提高光能利用率作为水稻高产育种的重要目标[4-10]。
国内外科学家在通过株型塑造进而提高水稻群体光能利用率和产量潜力方面开展了大量研究。1959年广东省农业科学院首次通过人工杂交成功培育出半矮秆水稻品种“广场矮”[11],由此引领“第一次绿色革命”之后,水稻株型育种成为国内外科学家关注的重点课题,并从水稻育种实践中提出了不同的高产育种株型模式。1959年角田重三郎首次提出了株型概念[12],1968年澳大利亚Donald则首次提出了“作物理想株型”[13]。20世纪70年代,日本学者松岛省三提出了水稻“理想株型”的理论和高产水稻的一些具体株型指标[14]。20世纪80年代末国际水稻研究所提出适合直播条件下“少蘖、大穗、茎秆粗壮”的水稻新株型育种计划[5]。杨守仁、陈温福等提出北方粳稻的“理想株型”[6]及“直立穗型”[7];黄耀祥等[15]根据华南双季稻区气候生态特点,在矮化育种基础上,于20世纪70年代提出了“丛化育种”、80年代提出了“半矮秆早长超高产育种”等株型模式[8],莫永生等提出了“高大韧”稻[16];周开达等[9]根据西南寡日照气候生态条件提出了“亚种间重穗型”模式;袁隆平[10]则提出了长江中下游一季中稻区的超级稻理想株型模式,程式华等[17]提出了“后期功能型”理想株型模式等。这些水稻株型模式的提出,丰富了株型研究的理论,并在育种和生产实践中发挥了重要的作用。然而,这些研究大多集中在常规水稻或杂交水稻品种生育后期株型的表现上,对前、中期株型特征研究不多[18-19],尽管陈友订等[19-20]、康文启等[21]、苏祖芳等[22-23]、孙成明等[24-25]对水稻动态株型方面开展了研究与探讨,但并未从动态株型特征与光能利用效率两方面进行综合分析。因此,进一步探索全生育期前、中、后不同发育时期均具有光能高效利用特性的理想动态株型,对于提高水稻产量潜力,尤其是提高生育期相对较短的华南双季稻区水稻品种的产量潜力,显得尤为重要。
众所周知,作物杂种F1的特性是由其父本恢复系和母本不育系共同决定的。但不少研究表明,籼型三系杂交稻和两系杂交稻,其杂种F1的生育期[26]、株高[26-29]、剑叶长[29-31]、剑叶宽[31]、剑叶基角[31]、剑叶面积[31]、倒2叶基角[31]、倒2叶长与宽和顶部3叶面积[31]、穗长[29]、每穗总粒数[26-29]、着粒密度[26-27]、生物产量[26]、谷草比[26]、千粒重[29-30]、结实率[27, 30]等性状主要受基因的加性效应控制,父本恢复系的影响大于母本不育系[26-31]。由此可见,评价和分析现有恢复系的株型动态变化特征和光能利用率,对于进一步筛选和培育具有光能高效利用特性的理想动态株型杂交稻恢复系,进而培育具有光能高效利用特性的籼型杂交稻,具有重要的实践意义。
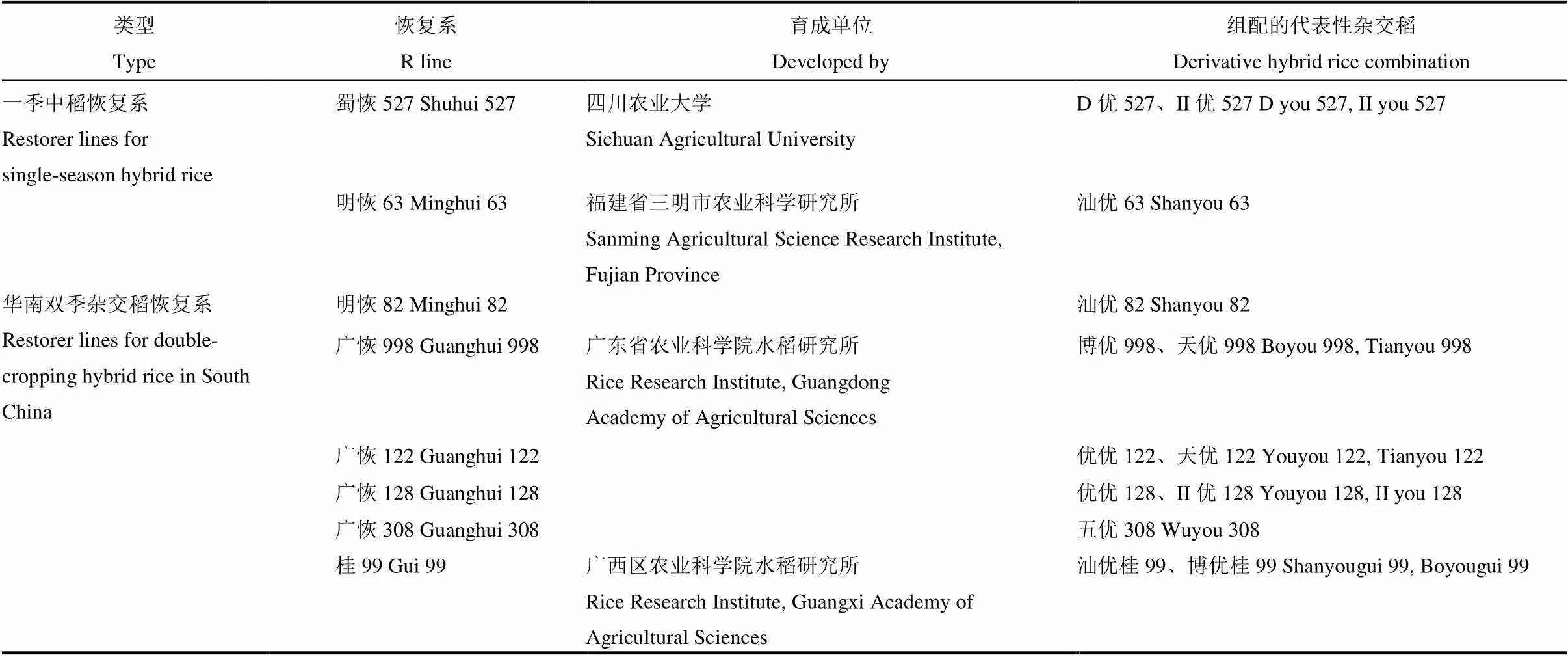
表1 供试恢复系及其代表性杂交稻组合
为此,本研究拟在华南双季稻区光温生态条件下对国内自1981年以来不同年代育成的大面积应用的籼型三系杂交稻恢复系动态株型特征与光能利用效率进行评价,以期为建立华南杂交稻恢复系光能高效利用的动态理想株型模式和培育光能高效利用的杂交稻新品种提供理论依据和技术支撑。
1 材料与方法
1.1 供试材料与种植
选用不同年代育成并在生产上大面积推广应用的8个籼型杂交稻恢复系(表1)作为供试材料,于2019年早季在广东省农业科学院广州大丰试验基地进行3次重复的随机区组试验。3月5日播种,4月9日插秧,每区栽插6行,每行12株,株行距16.7 cm×20 cm,单株植,普通肥水管理。
1.2 性状调查与数据分析方法
水稻不同生育时期的划分参照丁颖的划分标准[32]。
1.2.1 性状调查与测定方法
分蘖数动态调查:从移栽后的第14天(4月23日)开始,每隔4 d左右调查一次,并记录主要生育期,如分蘖盛期、分化始期、抽穗期和成熟期等。上部3叶开张角与叶片长、宽的测定:根据徐正进等[33]的方法,用量角器测量上部3叶开张角;分别在移栽后20 d和抽穗后用尺子直接测量10个单株主茎上的上部3片完全叶的长和宽;按照吉田昌一等[34]的方法计算剑叶的叶片面积。
叶片SPAD值(叶绿素含量)测定:用SPAD值大小表示叶色的浓淡,利用SPAD-502叶绿素仪测定。在5月10日(移栽后31 d)、6月2日(移栽后54 d)、6月19日(移栽后71 d)和7月2日(移栽后84 d)测定主茎顶部3片功能叶的SPAD值,每个小区测6株,每片叶测定4点,计算平均值。考虑到各恢复系间的生育进程不同,为便于在相同生育进程上比较各恢复系中后期的叶色变化(SPAD值),最后根据各恢复系的始穗期和前后4次的实测数据,利用插值法计算出各恢复系抽穗前第20天(中期)、抽穗期和抽穗后第20天(后期)的SPAD值。
叶面积指数(LAI)和透光率测定:利用美国生产的LI-COR LAI-2200C型号植物冠层分析仪在田间活体测定群体叶面积指数和透光率[35]。从移栽后17 d(4月26日)开始,随后在5月10日、6月2日、6月19日和7月2日,水稻生长的前、中、后3个时期共测定5次叶面积指数和透光率,并根据插值法计算出5月1日、5月6日和6月7日等3个时期的叶面积指数,用于估算各恢复系叶面积指数≥1.0的日期。此外,根据各恢复系始穗期和各生育期的实测数据,利用插值法估算出各恢复系在始穗前第15天(中期)的LAI和透光率,以及始穗后第20天(后期)的LAI和透光率。
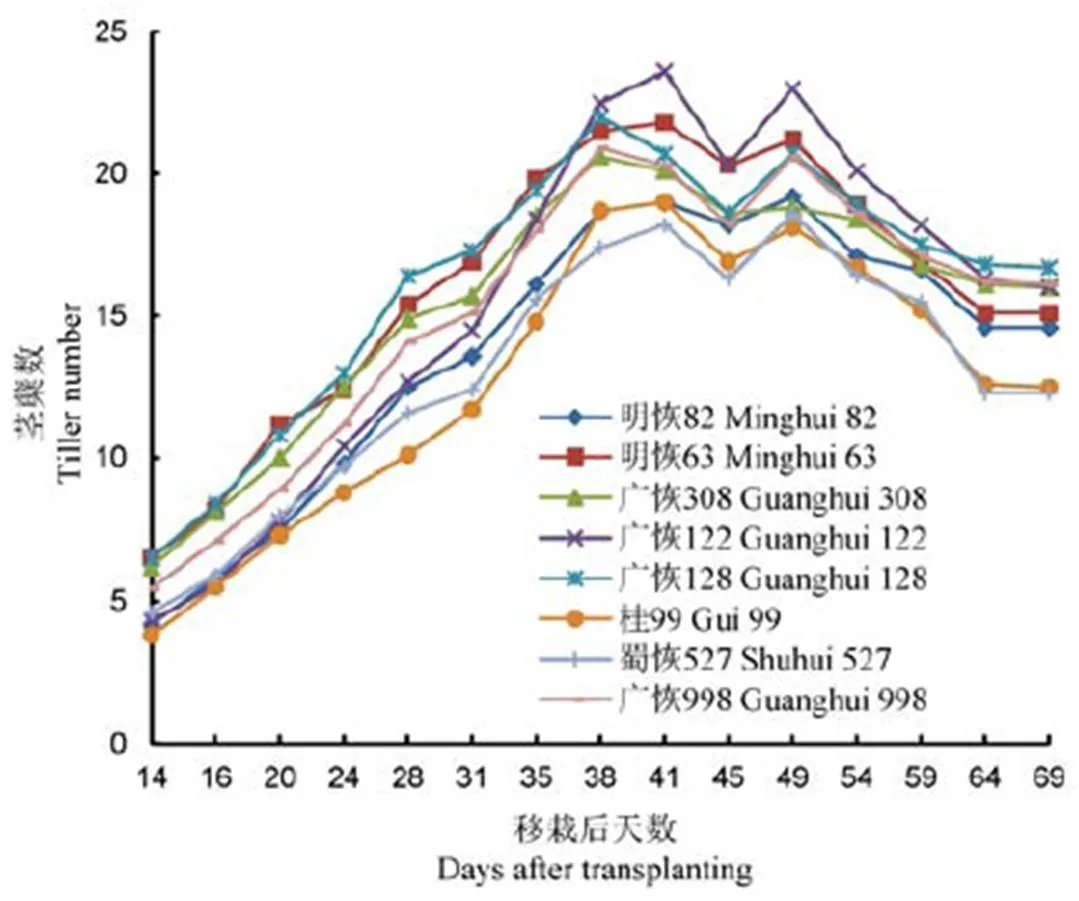
图1 移栽后分蘖数的动态变化
光合速率的测定:在各恢复系始穗期的上午,利用LI-6400XT便携式光合作用测量系统,在大田条件下直接测定主茎顶部3片功能叶,每个小区测3株,计算平均值。
单位面积产量与光能利用率计算:取小区非边行单株10株进行测产和考种分析,折合成每1 m2的稻谷产量。经济产量的光能利用率(RUE)估算方法参照黄农荣等[36]的方法进行。光能利用效率是指水稻在生长期内对太阳总辐射量的利用效率,计算公式:RUE(%)=(Ɛ×/)×100;其中,Ɛ为物质能量转换系数,指形成1 kg籽粒所需的能量(即燃烧热),稻谷的物质能量转换系数为15.5×106J/kg;为单位面积的稻谷产量(kg);为单位面积的秧田期(秧田面积按秧田∶本田=1∶10的比例计算)和本田生育期内的太阳总辐射量,即=∑(R),为每天的太阳辐射量, 由试验基地的EM50型科研级自动气象站(美国Dynamax公司生产)自动收集记录,表示播种到成熟的天数。
1.2.2 数据分析方法
利用Statistix 8.0软件对试验数据进行方差分析和LSD多重比较。不同英文字母表示不同材料间差异显著水平(<0.05);利用SPSS 19.0软件进行聚类分析。
2 结果与分析
2.1 茎蘖数动态变化
2019年4月9日插秧,移栽后14 d(4月23日)第一次调查茎蘖数。图1结果显示,每穴茎蘖数最高的是明恢63,随后依次是广恢128、广恢308、广恢998,最少的是桂99。明恢63、广恢128、广恢308和广恢998从移栽至移栽后31 d这段时间,分蘖速度明显高于其他恢复系;广恢122的茎蘖数在移栽35 d后开始超过广恢998和广恢308,移栽后第38天全面超过所有其他7个恢复系,成为分蘖启动慢,但后发性最强的恢复系。
各恢复系进入茎蘖峰值时间有明显差异。其中,广恢998、广恢308和广恢128移栽后第38天(5月17日)最早达到峰值;明恢63、蜀恢527、广恢122、明恢82和桂99至移栽后41 d(5月20日)才达分蘖峰值。说明广恢128、广恢308和广恢998不仅早生快发性好,而且进入茎蘖峰值时间早,这有利于减少无效分蘖和形成大穗。
由于移栽后第45天(5月24日)前后连续暴雨,分蘖加速衰亡,导致分蘖动态曲线在此处均出现一个小低谷。天气转好后,移栽后第49天又出现分蘖小幅增加。此后随着生育进程推进,小分蘖不断衰亡,茎蘖数逐渐减少,至移栽后64 d开始进入平稳期。
2.2 叶面积指数(LAI)动态变化
LAI的测定结果如表2所示。移栽后第17天供试恢复系LAI值的差异达极显著水平(=9.3,<0.001),明恢63的LAI最高,达到0.86,其次是广恢998,为0.51,最小的是桂99,仅为0.23。明恢63的LAI与其他7个恢复系的差异均达到显著水平;广恢998与明恢82、广恢128和广恢308等3个恢复系差异不显著,但与蜀恢527和桂99的差异均达显著水平。
LAI达到1.0以上,标志着水稻群体基本覆盖整个稻田,减少生育前期群体漏光。明恢63在移栽后第22天(5月1日)LAI就达到1.31,是最早达到LAI≥1.0的恢复系;其次是广恢998、广恢308、广恢128和蜀恢527,在移栽后第27天(5月6日)LAI达到1.0以上;明恢82和桂99于移栽后第31天才达到1.0以上;最迟的是广恢122,移栽后第31天其LAI还仅有0.99。供试恢复系LAI达到峰值的时间和峰值大小各不相同。其中,广恢998、广恢308、桂99、广恢128和广恢122在移栽后第54天达到峰值,而明恢63、蜀恢527和明恢82的LAI 峰值在移栽后第71天才达到。
为便于在相同生育进程上比较各供试材料的LAI大小,用插值法计算出抽穗前15 d(幼穗分化中期)和抽穗后第20天(后期)的LAI值(表2)。结果表明,抽穗前15 d(=86.7,<0.001)明恢63的LAI值最大,为4.32,与其他7个恢复系差异达到显著水平;其次是蜀恢527、广恢128、广恢998和广恢122,最小的是明恢82、广恢308和桂99。抽穗后第20天(后期)(=6.1,<0.005),LAI 值最大的还是明恢63,为4.72,其次是明恢82,为4.21,这二者差异不显著。LAI值最小的2个恢复系是广恢308和广恢998,其与明恢63、明恢82和蜀恢527的差异达到显著水平。
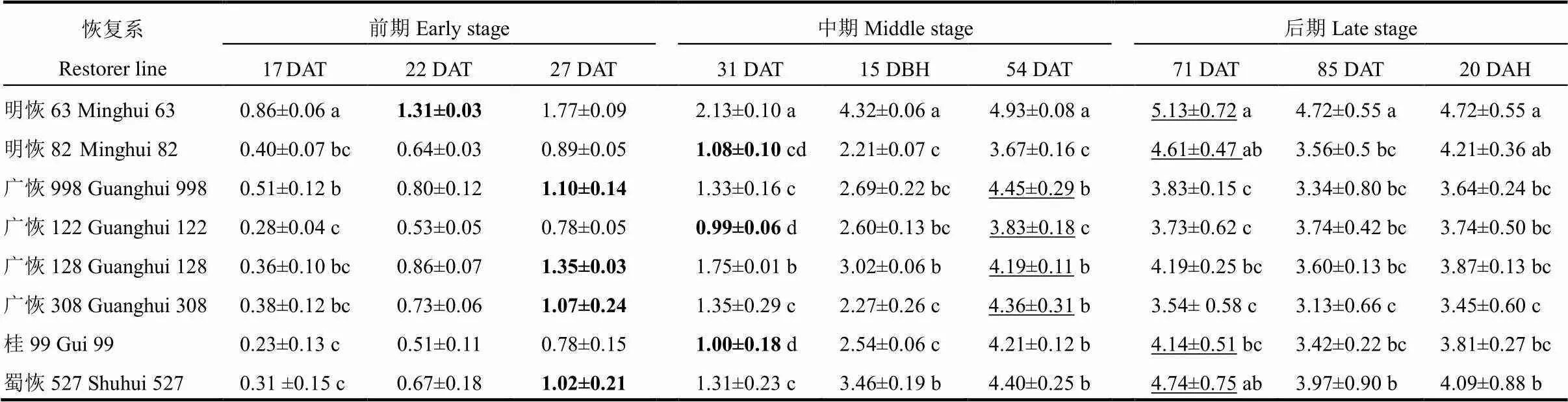
表2 不同时期LAI动态变化
DAT-移栽后天数;DBH-抽穗前;DAH-抽穗后。表中粗体数字表示该恢复系LAI首次达到1.0时的LAI值;有下划线的数值表示该恢复系茎蘖数达到峰值时的LAI值;表中数据为平均数±标准差(=3);采用单因素方差分析和LSD多重比较与显著性检验,同列中不同小写字母表示在0.05水平上差异显著(<0.05)。下同。
DAT means days after transplanting; DBH means days before heading; DAH means days after heading(the same in the following tables);The bold figure in the table represents the LAI value above 1.0 for the first time for the R line; The underlined figure refers to the LAI as the tiller number of the R line peaks. Data are mean±SD (=3). Values followed by different letters in the same column indicate significant difference among treatments at 5% probability level by the ANOVA and LSD all-pairwise comparisons test. The same as below.

表3 不同时期群体透光率变化动态
2.3 群体透光率动态变化
透光率测定结果如表3所示。前期即移栽后17天(4月26日),恢复系间透光率的差异达极显著水平(=7.73,<0.001),透光率最小的是明恢63和广恢128,分别为61.4%和67.9%;透光率最大的是广恢122和桂99,分别达到82.5%和80.9%。说明生育前期明恢63和广恢128群体漏光少,广恢122和桂99漏光率最大,其他4个恢复系居中。
中期即抽穗前第15天恢复系间透光率的差异达显著水平(=6.13,<0.005),明恢63和蜀恢527的透光率最小,仅为15.6%和19.1%,与其他6个恢复系差异达显著水平,说明其中期群体较大,通风透气性较差;明恢82的透光率最大,为31.7%,说明中期群体通风透光性最好;其他5个恢复系的透光率居中,均为23.7%~28.0 %,在中期群体较大情况下,其通风透光性较好。
后期即始穗后20 d恢复系间透光率的差异达显著水平(=9.85,<0.005),广恢308的透光率最大,达到11.5%,其他依次是广恢128、广恢998和广恢122,为10.16%~10.5%,这四者之间的差异未达到显著水平,而它们与其他4个恢复系的透光率差异均达显著水平,说明广恢308、广恢128、广恢998和广恢122后期群体通风透光性好;明恢63和明恢82后期透光率最低,分别为6.3%和6.9%,说明后期群体较为荫蔽,通风透光性差;蜀恢527和桂99的透光率分别为8.8%和8.7%,说明其后期群体通风透光性也较差。
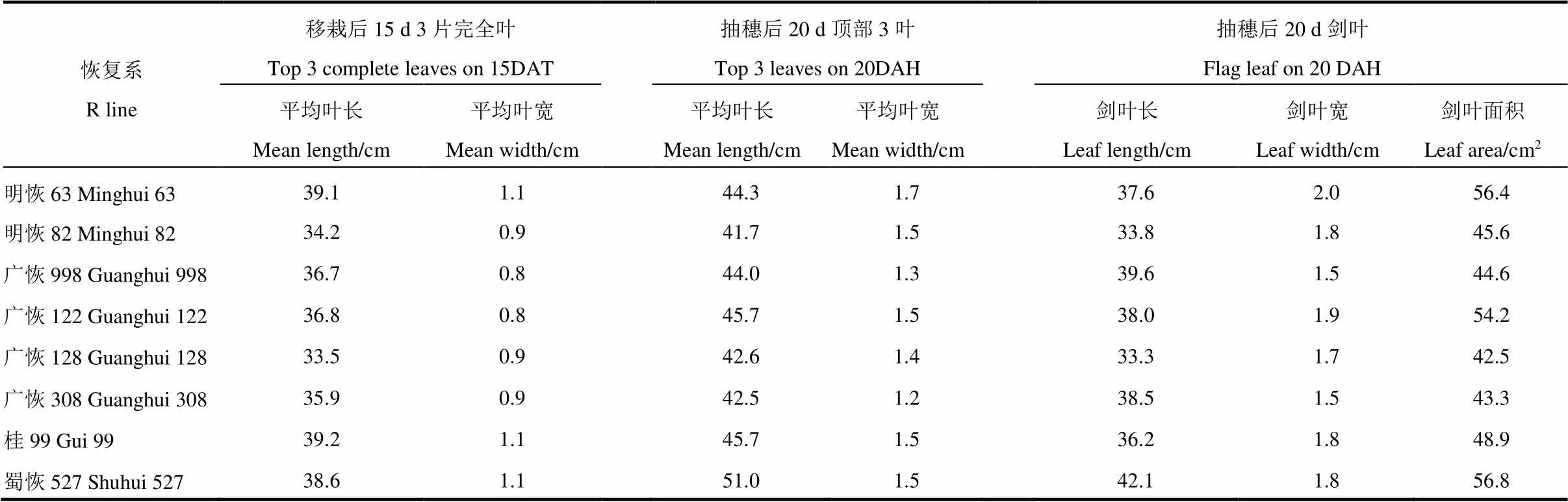
表4 不同生育时期顶部3片完全叶的长宽
2.4 上部3片完全叶长与宽
如表4所示,移栽后第15天,顶部3片完全叶平均长度最长的3个恢复系是桂99、明恢63和蜀恢527,分别为39.15 cm、39.01 cm和38.64 cm,叶片宽度也是最大的3个,为1.05~1.07 cm,均为宽长型大叶片。叶片最短的是广恢128,只有33.54 cm,叶片最窄的是广恢122和广恢998。其他恢复系叶长与叶宽均居中。
抽穗后20 d,顶部3叶平均长度最长的是蜀恢527,为51.0 cm,最短的是明恢82,为41.7 cm,其他为42.5~45.7,差异较小;3叶平均宽度最大的是明恢63,达到1.7 cm,最窄的是广恢308,为1.2 cm,广恢998次之,为1.3 cm。剑叶最长的也是蜀恢527,达到42.1 cm,其次是广恢998,为39.6 cm;剑叶最宽的明恢63,为2.0 cm,其次是广恢122;剑叶最窄的是广恢308和广恢998,均为1.5 cm。对比前期和后期叶片宽度,前期叶片宽大的恢复系后期叶片也是宽大的,这类恢复系后期(抽穗后第20天)的透光率(表3)也较低,不利于群体的通风透光;但前期叶片较窄的,后期叶片不一定就窄。如广恢122前期是8个恢复系中叶片最窄(0.8 cm)的,后期叶片的宽度反而较大,达到1.5 cm,仅次于叶片最宽的明恢63,明恢82也有相同的趋势;广恢308前期叶片宽度中等,后期叶片反而成为8个恢复系中最窄的(仅有1.2);只有广恢998是唯一前后期叶片较窄的恢复系。
从剑叶面积大小可看出,蜀恢527、明恢63和广恢122的叶面积最大,分别达到56.8 cm2、56.4 cm2和54.2 cm2。其他恢复系为42.5~48.9 cm2。根据周炳炎的划分[37],蜀恢527、明恢63和广恢122为大叶型;其他5个恢复系均为中叶型。后期叶片窄或较窄的中叶型恢复系广恢308、广恢998和广恢128,其群体后期的透光性是最高的(表3),说明这类恢复系后期有利于群体通风透光。
2.5 叶片开张角
叶片开张角是决定株型的重要因子之一,它与水稻植株的受光态势密切相关。表5结果显示,移栽后20 d(4月29日),明恢63和广恢122的倒3叶开张角最大,分别为90.3°和90.1°,其次是广恢998和广恢128,分别为85.8°和82.2°;开张角最小的是桂99,仅有42.5°。说明在生长前期明恢63、广恢122、广恢998和广恢128的株型较为披散,可减少群体漏光,这与前述群体透光率结果(表3)一致。
抽穗后20 d再次测定顶部3叶的开展角,结果表明(表5),广恢308的顶部3叶平均开张角最小,仅为33.9°,且剑叶、倒2和倒3叶的开张角度在8个供试恢复系中也是最小的,表现株型与叶型最为直挺;桂99的平均开张角最大,达到85.7°,但其剑叶、倒2和倒3叶的开张角在8个供试恢复系中均不是最大的。其他平均张角大小依次为蜀恢527>明恢63>广恢122>广恢998>广恢128>明恢82(63.5°~84.8°)。剑叶开张角度最大的是蜀恢527,倒2叶开张角最大的是明恢63,倒3叶开张角最大的是广恢122。根据开张角度可初步判断,广恢308、明恢82 、广恢128、广恢998和广恢122等在后期是群体受光姿态和通风透光较好的恢复系,桂99、蜀恢527和明恢63等后期群体受光姿态和通风透光性均稍差。但后期群体透光性除受顶部3片叶的开张角度大小影响外,还受LAI大小影响,尽管明恢82的开张角较小,由于后期它的LAI值较大(表2),后期群体透光率并不高(表3)。
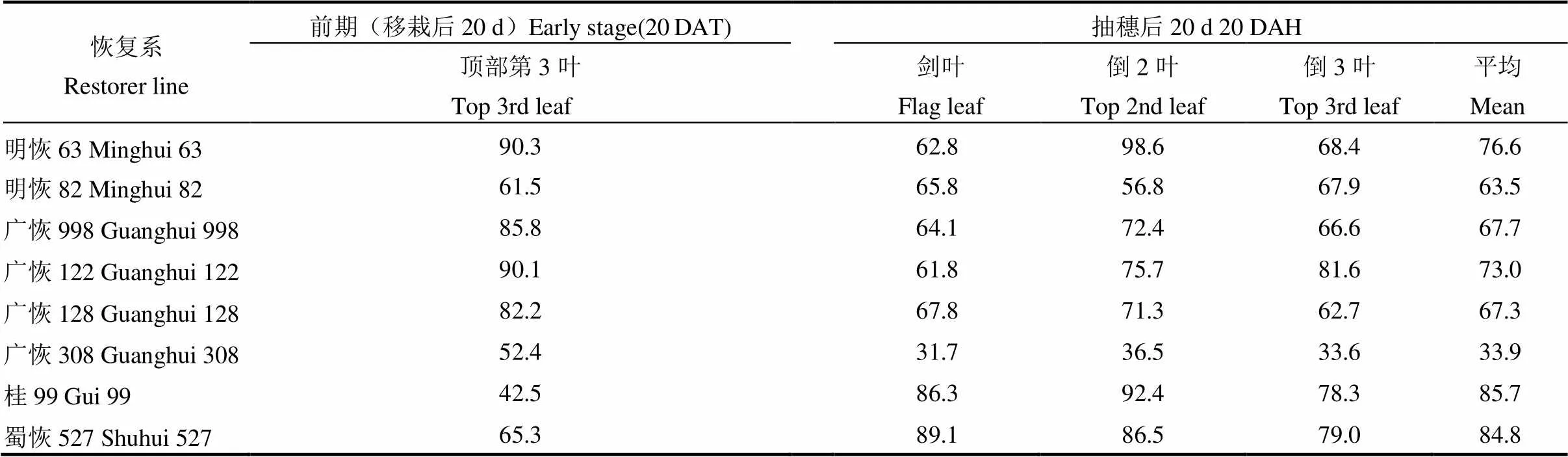
表5 前期顶部第3片叶的平均开张角度和抽穗后顶部3叶的开张角度

表6 叶片的SPAD值与光合速率
DBH and DAH mean days before and after heading, respectively.
2.6 叶绿素含量(SPAD值)与光合速率
叶片叶绿素含量(SPAD值)与光合速率关系密切[37]。在始穗前第20天(中期),8个供试恢复系的SPAD值差异极显著(=42.6,<0.001),其中,SPAD值最高的是桂99,为43.5,但它与广恢998、广恢308、广恢128和广恢122之间差异不显著。SPAD 值最低的是明恢63,为37.2,与其他7个恢复系的SPAD值差异达显著水平;在抽穗期(=10.83,<0.005),广恢308最高(43.9),明恢63最低,为38.7±0.6;抽穗后20 d(后期)SPAD值(=5.76,<0.005)最高的还是广恢308,为40.4;最低的则是蜀恢527和明恢63,分别为30.1和31.6,且二者差异不显著。从始穗前第20天至始穗后第20天这个时期SPAD值的变化态势来看,明恢63和广恢308是按照由较低-高-低的单峰曲线模式变化的,其他6个恢复系均是由高-较高-低的斜线模式变化。从SPAD值的变幅看,变幅最小的是广恢308(6.3%),变幅最大的是广恢128(23.0%),其他依次为蜀恢527>广恢122>明恢63>明恢82>广恢998>桂99。说明广恢128和蜀恢527后期易早衰;广恢308的叶色变化最小,有利于后期保持较强光合能力;其他6个恢复系的变化居中,较有利于后期转色顺调。
抽穗期各恢复系的叶片光合速率(表6)差异达显著(=6.18,<0.005),最高的广恢998为23.8 μmol/(m2·s),其次是广恢128、明恢82和广恢308。这4个恢复系的光合速率相当,差异不显著;最低的明恢63为20.4 μmol/(m2·s),但与桂99、蜀恢527、广恢122和广恢308等4个恢复系的差异也未达到显著水平。各恢复系的抽穗期位于6月1日至12日之间,除6月1日、6月5日和6月11日3天下雨外,其他日期上午的天气均基本正常。广恢308于6月1日始穗,其光合速率测定因下雨推迟到6月2日,其他恢复系均为始穗期当天测定完成。其中,明恢82、广恢122、广恢998和广恢128的测定当天上午天气晴好,蜀恢527、广恢308、桂99和明恢63的测定当天阳光不太稳定,对其光合速率的测定结果会有一定影响。
2.7 产量相关性状与光能利用率
表8结果显示,生育期广恢308最短,仅为122 d,其次是广恢998和明恢82,均为125 d,明恢63的最长,达135 d;每穗总粒数方面,明恢82最多,平均每穗187.7粒,明恢63最少,仅为123粒;结实率方面,广恢998最高,为84.8%,广恢128最低,仅为72.4%;有效穗数(=5.6,<0.005)方面,广恢998有效穗最多,达到228.0穗/m2,其次是广恢128,为222.0穗/m2,二者差异不显著。蜀恢527的有效穗最少,仅有147.0穗/m2;单位面积产量(=5.52,<0.005)方面,广恢998最高,为0.61 kg/m2,其次是明恢63,桂99最低,仅为0.51 kg/m2。广恢998、明恢63和明恢82三者之间的单位面积产量差异不显著,广恢998与其他5个恢复系之间差异达显著水平。

表8 稻谷产量性状与光能利用率
HD, Heading date; GD, Growth duration; TGW, Thousand-grain weight; SSR, Seed setting rate; EPN,Effective panicle number per m2; GY,Grain yield; RUE,Radiation use efficiency.

图2 供试恢复系RUE聚类结果
供试恢复系间的光能利用率(RUE)差异达极显著水平(=7.81,<0.001),广恢998最高,为0.479%,其次是广恢308、明恢82和明恢63,分别为0.452%、0.448%和0.441%,蜀恢527最低,仅为0.384%。广恢998与广恢308的RUE差异不显著,但与明恢82、明恢63和其他4个恢复系之间的差异均显著;广恢308与明恢82和明恢63差异不显著,与其他4个恢复系之间的RUE差异均达显著水平。利用系统聚类Ward法对8个恢复系进行聚类分析(图2),在欧氏距离25处划分为2类,其中RUE较高的三个恢复系广恢998、广恢308和明恢82聚在类群I,而RUE值相对较低的其他5个恢复系则聚在类群II。说明在华南生态条件下,广恢998、广恢308和明恢82同属于光能高效利用型恢复系。
对比各恢复系单位面积产量和光能利用率可见,明恢63的单位面积产量居第2高位(0.603 kg/m2),但其光能利用率(0.411%)反而低于产量位居第4的广恢308(0.452%)。这主要是因为明恢63的生育期比广恢308长11 d,其接受的光辐射总量大,因此,基于产量和有效辐射总量计算RUE值的高低,肯定与其产量高低不完全相同。
3 讨论
3.1 籼稻恢复系动态株型与光能利用率表现与评价
研究株型的目的是为了探索并塑造水稻理想株型以获得高产更高产[4-25, 38-39]。作物产量形成是在一定的空间和时间范围内综合光合能力的体现。水稻产量=[(光合时间×光合面积×光合能力)−光合产物的消耗]×经济系数,即由光合时间、光合面积、光合能力、光合产物消耗和光合产物的分配共同决定[1],空间上主要是群体捕获光能的能力,时间上主要与群体有效光合时间相关[3, 13, 17, 39],除了光能高截获能力和高光合能力之外,光合产物在籽粒中的分配比例也直接影响到最终的经济产量[39]。高亮之等[40]在水稻的前期、中期和抽穗期通过对4种典型的水稻株型(上挺下挺、上挺下披、上披下挺和上披下披)进行群体光合量的数值模拟,认为在水稻生长的前期,以上披下披株型的群体光合量为最高;在抽穗期则以上挺下披型的群体光合量最高;叶片的开张角反映了叶片在空间的伸展态势和植株的披散程度[22-25, 39]。本研究结果表明,前期(移栽后20 d)顶部3片完全叶的开张角较大的前4个恢复系依次是明恢63>广恢122>广恢998>广恢128,其平均开张角为82.2°~90.3°,均表现为上披下披型。其他4个恢复系桂99<广恢308<广恢82<蜀恢527,平均开张角相对较小,为42.5°~65.3°;中期随着节位升高,各恢复系的叶片也逐渐上举;始穗后20 d(后期)顶部3叶平均开张角较小的恢复系是广恢308<明恢82<广恢128<广恢998,为33.9°~67.7°;其他4个恢复系的开张角均超过70°,最大的桂99达到85.7°。由此可见,广恢998和广恢128前期的开张角大,后期开张角较小,基本属于高亮之等[39]认为的中后期群体光合量最大的上(后期)挺下(前期)披型。广恢308和明恢82前期、后期开张角度均小,属于上挺下挺型;明恢63和广恢122为上披下披型;蜀恢527为上披下挺型;桂99则为下挺上披型。对比光能利用效率结果可见,符合下(前期)披上(后期)挺株型的广恢998光能利用效率最高,达到0.479%,但同为下(前期)披上(后期)挺株型的广恢128,其光能利用率比上(后期)挺下(前期)挺株型的广恢308和明恢82还低。这可能主要与广恢128产量主要构成因子结实率较低(72.4%)、后期叶片SPAD值降幅大、叶片早衰有关。
水稻品种要获得更高的光能利用效率,除了应具备有利于光能高效捕获能力的动态株型外,还应具备光能高效利用与转化的生理机能[3]。本研究中广恢308尽管前期株型直挺,但分蘖早和快、群体LAI达到1.0以上和茎蘖达到峰值所需的时间均短,中后期株型直挺,通风透光性好,最终的光能利用率达到0.452%,仅次于最高的广恢998;明恢63后期叶片宽大、中后期群体透光率差;明恢82前期分蘖和叶面积发展较慢,群体漏光较多,但在抽穗后20天群体的LAI和叶片SPAD值维持在较高水平,且后期叶片转色好,较大的有效光合叶面积和较长的有效光合时间,最终其综合光能利用效率比较高,与居第二位的广恢308差异不显著;广恢128的前、中、后期其动态株型表现较为理想,但中后期叶色衰变较快,叶片有效光合时间短,结实率偏低(72.4%),导致其光能利用率显著低于最高的广恢998;广恢122、桂99和蜀恢527前期分蘖与叶面积发展均较慢,但广恢122在移栽后35 d左右分蘖迅速加快,茎蘖峰值是所有供试恢复系中最大的,但有效穗并不多,说明无效分蘖多;桂99前期开张角度小,群体漏光多,后期顶部3叶较长宽,叶片开张角较大,群体透光率较低;蜀恢527中后期叶片长宽、开张角大,群体披散荫蔽,透光性差。广恢998前、中、后期株型动态变化是所有供试材料中综合表现较为理想的恢复系,加之后期叶片较窄,抽穗后20 d的SPAD值较高,光合速率最高、结实率和单位面积有效穗数均是最高的,其产量和光能利用率也均为最高。由此可见,恢复系的动态株型表现理想与否,最终与其光能利用效率密切相关。
3.2 华南生态条件下的恢复系理想动态株型模式
不同稻作生态区,因其气候生态条件和品种生育期的长短各不相同,实现水稻高产稳产的理想动态株型也会存在差异,这就是为什么我国育种家分别提出了不同高产株型模式[5-10, 15-17]的重要原因。周炳炎[38]根据剑叶面积,将水稻品种分为大叶型(≥50 cm2)、中叶型(35~49 cm2)和小叶型(≤35 cm2)。在雨水较多、高温高湿的华南稻区,中叶型的窄长叶品种比大叶型的宽大叶品种更具优势。因为宽大叶品种,尤其是叶片不够厚的宽大叶品种,后期的叶片易承接更多雨水的重压,导致顶部3片主要功能叶披散,使群体隐蔽,通风透光差,极易滋生病虫,最后影响到产量和光能利用效率。窄长叶品种,尤其是后期顶部3叶厚直的品种,因其叶片窄小挺直,叶片承接雨水的叶面小,雨水对其的重压自然也小,后期叶片可保持挺直,群体通风透光性良好,有利于后期群体光能利用效率的提高。本研究中光能利用率相对较高的3个恢复系广恢998、广恢308和明恢82即属于这类叶片较窄的中叶型恢复系。
为此,笔者根据华南高温多湿的气候生态条件,认为华南双季稻区光能高效利用杂交稻恢复系的理想动态株型模式应当具备如下特点:
1)生育前期(移栽至幼穗分化):前期叶片窄长,株型较为松散;即移栽后15 d顶部3片完全叶的平均叶宽≤0.90 cm,平均叶长约37 cm,开张角≥80°,分蘖早、叶面积发展快,移栽后群体LAI达到1.0的时间≤27 d,以解决生育前期植株小、叶面积不足导致群体漏光严重和光能利用率不高问题。
2)生育中期(幼穗分化至始穗):进入幼穗分化阶段即停止分蘖,分蘖峰值出现早(早稻移栽后≤38 d,晚稻≤33 d),茎蘖数峰值≤600个/m2,无效分蘖少,减少营养消耗,使光合产物(非结构性碳水化合物)能够尽量累积储存在茎秆和叶鞘,为孕育大穗和后期灌浆充实提供充足营养[42];功能叶片SPAD值≥40,光合速率≥24 μmol/(m2·s),植株叶片随着节位的升高逐渐直立上举,株型集散适中,植株群体透光率≥25%。
3)生育后期(始穗至成熟期):顶部3叶平均叶宽≤1.25 cm(剑叶宽1.5 cm左右),平均叶长≤44 cm(剑叶长≤40 cm),开张角≤65°,较厚直;群体透光率≥10%,通风透光性好;早稻耐高温,晚稻耐低温,叶片转色顺调,不早衰,抽穗期与抽穗后20 d叶片的SPAD值降幅较小(≤13%),有效光合时间长,SPAD值为35~40;普通栽培条件下平均每穗总粒数为180左右,有效穗数300穗/m2以上,结实率达85%以上。
3.3 不同动态株型恢复系的光能利用率及其影响因素
水稻种植区域、季别、栽培管理水平和品种的株叶形态与生理特征,包括分蘖快慢、叶片大小与着生姿态、叶面积、光合速率等等,都直接影响水稻群体有效光合叶面积和光能利用效率[2-3, 22-25, 43-44]。例如,美国品种Lemont在光照强度大、日照时数长的生态条件下,表现出单叶的光合速率高、光饱和点高和较高的产量,但在阴雨天气较多的广东种植,其叶面积指数发展慢,光能截获能力差,产量很低[2]。由此可见,同一品种在不同生态区种植,其植株形态、光合效率与光能利用率的表现存在较大差异。本研究利用分别来自广东、广西、福建和四川等不同稻作生态区大面积应用的杂交稻恢复系为供试材料,在广州生态条件下种植,它们的株型动态表现各异,光能利用效率差异也明显。聚类分析结果表明,归为光能高效利用群(类群I)的广恢998、广恢308和明恢82,均为来自相近生态区的广东和福建,生育期较短,属早晚兼用型杂交稻恢复系,说明它们更适合华南的光温生态条件。但在长江流域一季中稻区大面积推广应用的杂交稻恢复系明恢63和蜀恢527,则均聚类在光能利用率较低的类群II,其光能利用效率分别为0.411%和0.384%。说明这两个在长江流域一季中稻区广泛应用的优良恢复系,在华南高温多湿、雨水较多的光温生态条件下表现不太适应,最突出的问题是生育期缩短、中后期叶片宽大、披散,易造成群体荫蔽。尤其是来自低温、阴雨、寡日照天气较多的四川的恢复系蜀恢527,似乎更不适应强光、高温高湿的华南生态条件。由此可见,在某一稻作区表现高产和光能高效利用的品种(系),引种至其他生态区,则不一定能表现出同样的株型、产量和光能利用效率。利用明恢63组配育成的大面积推广的杂交稻汕优63就是一个很好的例证,它在广东不论当早稻还是晚稻种植,一般都表现后期叶片宽披(国家水稻数据中心),但在四川、福建、湖北等一季中稻区种植,则株叶挺直,表现出很好的株型和较高的光能利用率与产量水平[45]。
3.4 利用具有理想动态株型的光能高效恢复系组配高产杂交稻的可行性
杂交稻的株型是由其母本不育系和父本恢复系共同决定的。要选育具有理想动态株型的杂交稻,必须首先从选育和塑造具有理想动态株型的恢复系和不育系着手。不少研究籼型杂交稻亲本配合力和遗传力的结果表明,生育期[26]、株高[26-29]、剑叶长[29-31]、剑叶宽[31]、剑叶基角[31]、剑叶面积[31]、倒2叶基角[31]、倒2叶长与宽和顶部3叶面积[31]、穗长[29]、每穗总粒数[26-29]、着粒密度[26-27]、生物产量[26]、谷草比[26]、千粒重[29-30]、结实率[27, 30]等性状的一般配合力方差大于特殊配合力方差,说明杂种F1代这些性状表现是由其亲本的基因加性效应所决定,且父本恢复系的影响大于不育系。前披后直(即前期叶片披垂,后期叶片直立)的动态株型性状,被认为是由单基因控制的显性性状[46]。因此,培育和塑造具有理想动态株型的光能高效利用恢复系,有利于高产稳产杂交稻的培育。本研究结果表明,广恢998也具有前披后直的动态株型特征,且该恢复系具有光能利用效率高、有效穗数多、结实率高等特点,利用其组配的杂交稻一般也具有叶片较窄、株型前披后直、有效穗多、结实率和产量高等特点[47]。这与王贵学等[46]的结论一致,进一步证明了恢复系的理想动态株型,对其杂交稻株型特性具有重要的决定作用。
育种实践表明,利用本研究中产量和光能利用效率均最高的恢复系广恢998、产量与光能利用率均较低的恢复系桂99和广恢122,分别与不育系博A组配,育成的杂交稻博优998、博优桂99(原名博优903)和博优122,产量水平也各不相同。博优998和博优903(对照)于2001-2002年同时参加华南晚籼组区域试验,前者平均亩产分别为493.6 kg和439.6 kg,分别比博优903增产12.72%和6.10%,差异均达极显著水平;2000年晚造参加广东省区试,博优998平均亩产为466.5 kg,比博优122增产6.65%,增产极显著(国家水稻数据中心);广恢998和明恢63分别与不育系珍汕97A 组配,育成汕优998和汕优63,1999年晚造参加广东省区试,汕优998亩产为439.7 kg比汕优63增产4.89%(国家水稻数据中心)。此外,利用本研究中光能利用率最高的广恢998(0.479%)和第二高的广恢308 (0.452%)分别与天丰A 和五丰A组配,育成的杂交稻天优998和五优308均达到超级稻产量指标,并被农业部认定为超级稻。黄农荣等研究华南双季稻主栽品种的光能利用效率结果表明,天优998和五优308的光能利用率分别达到0.58%和0.61%,均被划分为光能高效利用型品种[36]。
生产上,广恢998组配育成的博优998、天优998、优优998等系列组合,在国家和省级区试中,均表现为有效穗数多,结实率高、高产稳产。其中,博优998连续15年成为国家区试对照种,超级稻天优998连续6年成为广东、江西和国家区试对照种[47]。广恢308组配的超级稻五优308也显示出广泛的适应性和高产稳产特性,连续多年成为国家和江西、湖南等省的区试对照品种。这几个品种连续多年成为我国南方稻区的主推品种,在生产上发挥了巨大作用,已累计推广面积超过800多万hm2(国家水稻数据中心)。据统计,广恢998是国内组配杂交稻最多的恢复系之一[48]。由此可见,利用产量和光能利用率高的恢复系组配选育的杂交稻的产量和光能利用率一般也高;而由产量和光能利用率较低的恢复系组配的杂交稻的产量和光能利用率则较低。
尽管籼型杂交稻中很多重要株型、产量等相关性状受父本恢复系的影响较母本不育系大,但不等于说母本不育系不重要。例如,本研究中恢复系广恢128虽然前、中、后不同时期均具有较好的动态株型,但其后期叶片转色不顺调,结实率偏低,导致其本身的产量和光能利用效率并不高。然而,利用其与后期转色好、结实率很高的优IA、II-32A和龙特甫A组配,育成的优优128、II优128和特优128表现高产稳产,其中,优优128于1997-1998年国家华南早籼迟熟组区试,比对照品种平均增产13%以上(国家水稻数据中心);II优128于1995-1996年广东省区试,比对照汕优63增产5.69%-6.35%。说明动态株型理想,但存在其他不利性状,通过选用优势互补的不育系进行组配,也能组配出优良的杂交稻,但与本身具有理想动态株型、高产与光能高效兼具的恢复系相比,选育出高产杂交稻组合的育种效率可能会低些。
本研究结果和育种实践充分说明,通过培育具有理想动态株型的光能高效恢复系技术途径,进而选育出高产稳产、光能高效利用的杂交稻新组合,不仅行之有效,而且育种效率可以大大提高。
谢 辞:本研究得到钟旭华研究员、黄农荣研究员、张彬副研究员在透光率等性状测定、实验数据处理、文稿修改等方面的大力支持与帮助,在此表示感谢!
[1] 潘瑞炽. 水稻的光能利用[J]. 广东师范学报: 自然科学版, 1976(1): 44-54.
Pan R C. Light energy utilization of rice [J].:, 1976(1): 44-54. (in Chinese)
[2] 屠曾平. 水稻光合特性研究与高光效育种[J]. 中国农业科学, 1997, 30(3): 28-35.
Tu Z P. Studies on the photosynthetic characteristics of rice and breeding for high use efficiency of solar energy [J]., 1997, 30(3): 28-35. (in Chinese with English abstract)
[3] 程建峰, 沈允刚. 作物高光效之管见[J]. 作物学报, 2010, 36(8): 1235-1247.
Cheng J F, Shen Y G. My humble opinions on high photosynthetic efficiency of crop[J]., 2010, 36(8): 1235-1247. (in Chinese with English abstract)
[4] Khush G S. What it will take to feed 5.0 billion rice consumers in 2030[J].,2005, 59: 1-6.
[5] Peng S, Khush G S, Cassman K G. Evolution of the new plant ideotype for increased yield potential[C]//Breaking the yield barrier: Proceedings of A Workshop on Rice Yield Potential in Favorable Environments. IRRI, 1994: 5-20.
[6] 杨守仁. 水稻超高产育种的新动向—理想株型与优势利用相结合[J]. 沈阳农业大学学报, 1987, 18(1): 1-5.
Yang S R. The new trend of rice super high-yield breeding: Combination of ideal plant type and heterotic utilization [J]., 1987, 18(1): 1-5. (in Chinese with English abstract)
[7] Chen W F, Xu Z J, Zhang W Z, Zhang B L, Yang S R. Creation of new plant type and breeding rice for super high yield[J]., 2001, 27(5): 665-672.
[8] 黄耀祥. 半矮秆、早长根深、超高产、特优质中国超级稻生态育种工程[J]. 广东农业科学, 2001(3): 2-6.
Huang Y X. The ecological breeding engineering of semi-dwarf, early growing and deep root, super high yield and very high grain quality[J]., 2001(3): 2-6. (in Chinese)
[9] 周开达, 马玉清, 刘太清, 沈茂松. 杂交水稻亚种间重穗型组合的选育: 杂交水稻超高产育种的理论与实践[J]. 四川农业大学学报, 1995, 13(4): 403-407.
Zhou K D, Ma Y Q, Liu T Q, Sheng M S. The Breeding of sub-specific heavy ear hybrid rice: Exploration about super high yield breeding of hybrid rice [J]., 1995, 13(4): 403-407. (in Chinese with English abstract)
[10] 袁隆平. 超级杂交稻育种[J]. 杂交水稻. 1997, 12(6): 1-6.
Yuan L P. Hybrid rice breeding for super high yield[J]., 1997, 12(6): 1-6. (in Chinese)
[11] 广东省农业科学院. 广东水稻矮化育种工作的基本总结[J]. 作物学报, 1966(1): 1-7.
Guangdong Academy of Agricultural Sciences. Preliminary summary of the semi-dwarf rice breeding in Guangdong[J]., 1966(1): 1-7. (in Chinese)
[12] 薛德榕. 水稻生理生态译丛(1)[M]. 北京: 农业出版社, 1981.
Xue D R. Translation of Rice Physiology and Ecology (1) [M]. Beijing: Agricultural Press, 1981. (in Chinese)
[13] Donald C M. The breeding of ideotypes[J]., 1968, 17(3): 385-403.
[14] 松岛省三. 稻作的理论与技术[M]. 庞诚, 译. 北京: 中国农业出版社, 1981.
Matsushima S. Theory and Technology of Rice Cultivation [M]. Translated by Pang Cheng. Beijing: China Agricultural Press, 1981. (in Chinese)
[15] 黄耀祥. 水稻丛化育种[J]. 广东农业科学, 1983(1): 1-5.
Huang Y X. Rice cluster breeding[J]., 1983(1): 1-5. (in Chinese)
[16] 莫永生, 何龙飞, 黄天进, 韦政, 农友业. 高大韧稻育种论[J]. 中国农学通报, 2004, 20: 82-86.
Mo Y S, He L F, Mo T J, Wei Z, Nong Y Y. A new rice breeding pathway: Higher, thicker and tougher rice breeding [J]., 2004, 20: 82-86.
[17] 程式华, 曹立勇, 陈深广, 朱德峰, 王熹, 闵绍楷, 翟虎渠. 后期功能型超级杂交稻的概念及生物学意义[J]. 中国水稻科学, 2005, 19(3): 280-284.
Cheng S H, Cao L Y, Chen S G, Zhu D F, Wang X, Min S K, Zhai H Q. Conception of late-stage vigor super hybrid rice and its biological significance[J]., 2005, 19(3): 280-284.(in Chinese with English abstract)
[18] 石利娟, 邓启云, 刘国华, 庄文, 陈立云. 水稻理想株型育种研究进展[J]. 杂交水稻, 2006, 21(4): 1-6.
Shi L J, Deng Q Y, Liu G H, Zhuan W, Chen L Y. Progress in ideal plant type breeding in rice[J]., 2006, 21(4): 1-6. (in Chinese with English abstract)
[19] 陈友订, 张旭. 华南水稻动态株型研究[M]. 上海: 上海科学技术出版社, 2011.
Chen Y D, Zhang X. Research on Dynamic Plant Type in Indica Rice[M]. Shanghai: Shanghai Scientific and Technical Publishers, 2011. (in Chinese with English abstract)
[20] 陈友订, 刘传光, 周新桥, 陈达刚, 郭洁, 陈平丽, 万邦惠, 张旭. 华南双季超级籼稻动态株型育种理论研究与应用[J]. 广东农业科学, 2019, 46(9): 8-17.
Chen Y D, Liu C G , Zhou X Q, Chen D G, Guo J, Chen P L, Wan B H, Zhang X. Studies and applications on dynamic plant type breeding theories in double cropping superL. subsp.Tingin South China [J]., 2019, 46(9): 8-17. (in Chinese)
[21] 康文启, 欧阳由男, 董成琼, 朱练峰, 禹盛苗, 许德海, 金千瑜. 水稻动态株型模式及其指标探讨[J]. 中国稻米, 2007, (1): 1-6.
Kang W Q, Ouyang Y N, Dong C Q, Zhu L F, Yu S M, Xu D H, Jin Q Y. Investigation on dynamic plant type model and its index in rice [J]., 2007, (1): 1-6.(in Chinese)
[22] 苏祖芳, 孙成明, 张亚洁. 高产水稻生育前期株型指标的研究[J]. 作物学报, 2002, 28(5): 660-664.
Su Z F, Sun C M, Zhang Y J. Study on index of plant type in early stage of high yield rice [J]., 2002,28(5): 660-664. (in Chinese with English abstract)
[23] 苏祖芳, 许乃霞, 孙成明, 张亚洁. 水稻抽穗后株型指标与产量形成关系的研究[J]. 中国农业科学, 2003, 36(1): 115-120.
Su Z F, Xu N X, Sun C M, Zhang Y J. Study on the relationship between rice plant type indices after heading stage and yield formation[J].. 2003, 36(1): 115-120. (in Chinese with English abstract)
[24] 孙成明, 苏祖芳, 许乃霞, 周培南. 水稻有效分蘖叶龄期株型特征与产量关系初探[J]. 江苏农业研究, 2000, 21(3): 10-15.
Sun C M, Su Z F, Xu N X, Zhou P N. Relationship between characters of plant type in critical stage of effective tillering emergence and yield in rice [J]., 2000, 21(3): 10-15.
[25] 孙成明, 苏祖芳, 张亚洁, 沙爱红, 桑大志. 水稻拔节期株型特征及其与产量关系研究[J]. 扬州大学学报, 2002, 23(2): 46-49.
Sun C M, Su Z F, Zhang Y J, Sha A H, Sang D Z. Study on relationship between characters of plant type in elongation stage and yield in rice [J]., 2002, 23(2): 46-49.
[26] 宋宇, 邹小云, 贺浩华, 傅军如, 李海波, 徐亮, 辛晓云. 籼型三系杂交水稻产量及其相关性状的配合力分析[J]. 江西农业大学学报, 2004, 26(5): 719-725.
Song Y, Zou X Y, He H H, Fu J R, Li H B, Xu L, Xin X Y. Analysis on combining ability of yield characters and related characters in three-line indica hybrid rice [J]., 2004, 26(5): 719-725. (in Chinese with English abstract)
[27] 张利华, 王建军, 李旭晨, 王林友. 籼型三系杂交水稻主要农艺性状的配合力分析[J]. 浙江农业科学, 2003, 15(1): 1-7.
Zhang L H Wang J J, Li X C, Wang L Y. Analysis on combining ability of main agronomic characters in three-line indica hybrid rice [J]., 2003, 15(1): 1-7. (in Chinese with English abstract)
[28] 涂诗航, 张水金, 董瑞霞, 杨东, 谢鸿光, 郑家团. 籼型三系杂交水稻亲本主要农艺性状配合力及遗传力分析[J]. 福建农林大学学报, 2008, 37(3): 230-234.
Tu S H, Zhang S J, Dong R X, Yang D G, Xie H G, Zheng J T. Analysis of combining ability and heritability of the major agronomic characters in some parents of three-line Indica hybrid rice[J]., 2008, 37(3): 230-234. (in Chinese with English abstract)
[29] 齐绍武, 盛孝邦. 籼型两系杂交水稻主要农艺性状配合力及遗传力分析[J]. 杂交水稻, 2000, 15(3): 38-41.
Qi S W, Shen X B. Analysis on combining ability and heritability of major agronomic characters in two line indica hybrid rice [J]., 2000, 15(3): 38-41. (in Chinese with English abstract)
[30] 廖伏明, 周坤炉, 盛孝邦, 阳和华, 徐秋生. 籼型三系杂交水稻主要农艺性状配合力研究[J]. 作物学报, 1999, 25(5): 622-631.
Liao F M, Zhou K R, Sheng X B, Yang H H, Xu Q S. Studies on combining ability of major agronomic characters in three-line indica hybrid rice [J]., 1999, 25(5): 622-631. (in Chinese with English abstract)
[31] 邓启云, 石利娟, 庄文, 袁隆平. 杂交水稻叶片形态性状配合力分析[J]. 沈阳农业大学学报, 2007, 38(5): 695-700.
Deng Q Y, Shi L J, Zhuang W, Yuan L P. Analysis on combining ability of leaf morphological characters in hybrid rice[J]., 2007, 38(5): 695-700. (in Chinese with English abstract)
[32] 丁颖. 中国水稻栽培学[M]. 北京: 农业出版社, 1961.
Ding Y. Rice Cultivation in China [M]. Beijing: Agriculture Press, 1961. (in Chinese)
[33] 徐正进, 董克. 水稻叶片基角、开张角和披垂度的同时测定方法[J]. 沈阳农业大学学报, 1991, 22(2): 185-187.
Xu Z J, Dong K. Simultaneous determination of base angle, opening angle and drape of rice leaves [J]., 1991, 22(2): 185-187. (in Chinese with English abstract)
[34] 吉田昌一, 福尔若D A, 科尔J H. 水稻生理学实验手册[M]. 北京: 科学出版社, 1975: 65-68.
Yoshida S, Forno D A, Gock J H. Laboratory manual for physiological studies of rice[M]. Beijing: Science Press, 1975: 65-68. (in Chinese)
[35] Garrigues S, Shabanov N V, Swanson K, Morisette J T, Baret F, Myneni R B. Intercomparison and sensitivity analysis of leaf area index retrievals from LAI-2000, AccuPAR, and digital hemispherical photography over croplands [J]., 2008, 148(8/9): 1193-1209.
[36] 黄农荣, 傅友强, 钟旭华, 梁开明, 潘俊峰, 刘彦卓, 胡香玉, 彭碧琳, 陈荣彬, 胡锐. 华南双季稻主栽品种的光能利用效率及聚类分析[J]. 中国生态农业学报, 2019, 27(11): 1714-1724.
Huang N R, Fu Y Q, Zhong X H, Ling K M,Pan J F, Liu Y Z, Hu X Y, Peng B L, Chen R B, Hu R. Radiation use efficiency and its classification of main varieties cultivars in double-cropping rice region of South China [J]., 2019, 27(11): 1714-1724. (in Chinese with English abstract)
[37] 方志伟, 张荣铣, 朱培仁. 水稻叶片叶绿素含量的变化与光合作用的关系[J]. 南京农业大学学报, 1987, 10(4):18-22.
Fang Z W, Zhang R X, Zhu P R. Change in chlorophyll content and photosynthetic rate and their relationship in individual leaf from fully expanded stage to senescence[J]., 1987, 10(4): 18-22.
[38] 周炳炎.水稻理想株型与超高产育种途径的探讨[J]. 湖北农学院学报, 1995, 15(1): 52-60.
Zhou B Y. Discussion on ideal plant type and super high yield breeding of rice [J]., 1995, 15(1): 52-60. (in Chinese)
[39] Peng S B, Khush G S, Virka P, Tang Q Y, Zou Y B. Progress in ideotype breeding to increase rice yield potential [J]., 2008, 108: 32-38.
[40] 高亮之, 金之庆, 张更生, 石春林, 葛道阔. 水稻最佳株型群体受光量与光合量的数值模拟[J]. 江苏农业学报, 2000, 16(1): 1-9.
Gao L Z, Jin Z Q, Zhang G S, Shi C L, Ge D K. A numerical model to simulate the incident radiation and photosynthate for rice canopies with optimum plant type [J]., 2000, 16(1): 1-9. (in Chinese with English abstract)
[41] 孙宏伟.水稻超高产育种的可能途径[J]. 垦殖与稻作, 2006(1): 3-5.
Sun H W. The possible approaches of super high yield breeding in rice[J].. 2006(1): 3-5. (in Chinese)
[42] Horie T. Increasing yield potential in irrigated rice: Breaking the yield barrier//Peng S, Hardy B. Rice Research for Food Security and Poverty Alleviation [M]. International Rice Research Institute, Los Banos, Philippines, 2001: 3-25
[43] 胡凝, 吕川根, 姚克敏, 邹江石. 卷叶水稻的光分布模拟及适宜叶面卷曲度分析[J]. 中国水稻科学, 2008, 22(6): 617-624.
Hu N, Lu C G, Yao K M, Zou J S. Simulation on photosynthetically active radiation distributing in rice canopy with rolled leaves and its optimum leaf rolling index[J]., 2008, 22(6): 617-624. (in Chinese with English abstract)
[44] 李绪孟, 王小卉, 郑华斌, 黄璜. 水稻群体冠层光分布及光合作用模型[J]. 生物数学学报, 2014, 29(1): 87-98.
Li X M, Wang X H, Zheng H B, Huang H. A rice canopy light distribution and photosynthesis model [J]., 2014, 29(1): 87-98. (in Chinese with English abstract)
[45] 谢华安. 汕优63选育理论与实践[M]. 北京: 中国农业出版社, 2005: 32-61.
Xie H A. Breeding Theory and Practice of Shanyou 63 [M]. Beijing: China Agriculture Press, 2005: 32-61. (in Chinese)
[46] 王贵学, 黄俊丽, 金良, 郑家奎, 张子龙. 水稻动态株型材料的遗传特性[J]. 重庆大学学报, 2006, 29(7): 108-110.
Wang G X, Huang J L, Jin L, Zheng J K, Zhang Z L. Heredity of rice transforming leaf morphology[J]., 2006, 29(7): 108-110. (in Chinese with English abstract)
[47] 廖亦龙, 王丰, 符福鸿, 柳武革, 李金华, 朱满山, 黄慧君, 刘迪林, 付崇允. 优质抗病恢复系广恢998的选育及其组合的推广应用[J]. 中国稻米, 2018, 24(2): 87-90.
Liao Y L, Wang F, Fu F H, Liu W G, Li J H, Zhu M S, Huang H J, Liu D L, Fu C Y. Breeding of restorer line Guanghui 998 with good grain quality and blast resistance and the extension and utilization of its hybrid combinations [J]., 2018, 24(2): 87-90. (in Chinese with English abstract)
[48] 鄂志国, 程本义, 孙红伟, 汪玉军, 朱练峰, 林海, 王磊, 童汉华, 陈红旗. 近40年我国水稻育成品种分析[J]. 中国水稻科学, 2019, 33(6): 523-531.
E Z G, Cheng B Y, Sun H W, Wang Y J, Zhu L F, Lin H, Wang L, Tong H H, Chen H Q. Analysis on Chinese improved rice varieties in recent four decades[J]., 2019, 33(6): 523-531. (in Chinese with English abstract)
Evaluation of Dynamic Plant Type and Radiation Use Efficiency ofHybrid Rice Restorer Lines
WANG Feng#,*, LIAO Yilong#, LIU Wuge, LIU Diling, ZENG Xueqing, FU Youqiang, ZHU Manshan,LI Jinhua, FU Chongyun, MA Xiaozhi, HUO Xing
(,,,;;,:)
【】A restorer (R) line with ideal dynamic plant type (DPT) is the premise of hybrid rice breeding with high and stable yield. The purpose of this study is to evaluate the DPT and radiation use efficiency (RUE) of the R lines bred in different periods, and investigate the characteristics of ideal DPT for efficient utilization of light energy, and lay a theoretical basis for breeding of high-yielding hybrid rice with high RUE. 【】A field experiment was conducted under the ecological conditions of South China with eight widely and commercially used R lines ofhybrid rice Minghui 63, Minghui 82, Shuhui 527, Guanghui 998, Guanghui 308, Guanghui 128, Guanghui 122 and Gui 99 as material. The dynamic changes of tiller number, leaf area index (LAI), light transmittance of the population, leaf size, leaf angle and leaf color (SPAD value), as well as the photosynthetic rate in the heading stage, heading date, yield related traits and RUE were measured. 【】There existed significant difference in dynamic changes of plant type, yield and RUE among the restorer lines. Ideal DPT was closely related to RUE.With the most ideal DPT under the ecological conditions of South China, Guanghui 998, Guanghui 308 and Minghui 82 had the highest RUE.According to the cluster analysis, Guanghui 998, Guanghui 308 and Minghui 82 were classified into ClusterⅠwith high RUE, and the other five restorers fallen into ClusterⅡ.Finally, based on the ecological conditions in South China,an ideal DPT model of rice for efficient utilization of light energy was proposed. And the key indicators of breeding and selection were as follows: a) The tiller number and leaf area increased rapidly after transplanting; it cost less than 27 d to make LAI≥1.0, and less than 38 d to peak tiller number with moderate tillering ability, and the highest tiller number less than 600/m2; b) Narrow, droopy and long leaves in the early stage. On15 days after transplanting, the average length, width and opening angle of the top three complete leaves were about 37 cm, ≤0.9 cm and ≥80°, respectively; narrow and straight leaf in late stage; on the 20th day after heading, the average opening angle, length and width of the top three leaves were ≤65°, 43 cm and ≤1.3 cm, respectively, belonging to medium sized leaf type. c)In the late stage, the leaf color turned from green to orange or yellowish gradually without premature senescence.The SPAD values ranged from 35 to 40, the decrease of SPAD value between 20 days before heading and 20 days after heading was less than 13%; the average number of grains per panicle was about 180 with the number of effective panicles more than 300 / m2and seed setting rate≥85%. 【】Guanghui 998 and Guanghui 308 have better DPT and higher RUE. The hybrid rice Tianyou 998, Boyou 998 and Wuyou 308 derived from these restor lines, have become the leading rice varieties for many years in southern China. Breeding hybrid rice with high and stable yield through the developing of R lines with ideal DPT and high RUE is an effective and important way.
hybrid rice; restorer line; radiation use efficiency; dynamic plant type; double rice-cropping region; South China
10.16819/j.1001-7216.2021.0207
2020-02-24;
2020-09-27。
国家重点研发计划资助项目(2017YFD0100102);国家现代农业产业技术体系建设专项(CARS-01-18);广东省重点领域研发计划资助项目(2018B020202004);广东省农业科学院科技创新战略专项(高水平农科院建设项目);广东省农业科学院农业优势产业学科团队建设项目。
