151 份大豆种质的CDDP 标记遗传多样性分析
2021-03-16王雪娇苏二虎赵晓宇贾利敏王新华
王雪娇,苏二虎,李 强 *,赵晓宇 ,陈 强,贾利敏 ,王新华
(1.内蒙古自治区农牧业科学院,内蒙古呼和浩特 010031;2.杭锦后旗农牧业技术推广中心,内蒙古杭锦后旗 015400)
DNA 分子标记是以个体间核苷酸序列变异为基础的遗传标记,它是生物个体在DNA 水平上遗传变异的直接反映。目前,许多分子标记技术被应用于作物种质资源的遗传多样性分析、QTL 定位及分子标记辅助育种等方面,但它们大多属于随机DNA 分子标记,所得位点通常与目的基因距离较远、目标性较差。CDDP 标记就是一种新型目的基因标记技术,其引物来自于 WRKY、MYB、ERF、KNOX、MADS 及ABP 等转录因子基因家族,涉及到植物生长发育、抗病、代谢等多个性状。Poczai 等[1]利用CDDP 与ITM(intron-targeting markers)相结合的方法分析了欧白英的遗传多样性,标记多态性条带百分率为90%;李莹莹等[2]利用CDDP 标记研究了菏泽牡丹的遗传多样性,标记多态性比率高达95.58%;Li 等[3]对菊花进行了CDDP 遗传多样性研究,多态性比率也高达92.53%,可有效地揭示菊花品种间的亲缘关系;17条引物共扩增出120 个DNA 条带,多态性条带百分率为68.33%,平均多态性信息含量为0.644,在引物多态性方面较以上研究略低,可能是由于物种间基因组差异较大造成的。
1 材料与方法
1.1 试验材料
选取151 份国内外不同地理来源的大豆代表性品种。国内品种主要来源于黑龙江(42 个)、吉林(40个)、内蒙古(15 个)、辽宁(9 个),其他省区 13 个,国外品种32 个。
1.2 DNA 的提取
采用天根公司植物基因组DNA 提取试剂盒提取151 份大豆材料总DNA。用1.2%琼脂糖凝胶电泳和紫外分光光度计检测其质量及完整性,根据检测结果,将总DNA 稀释为20 ng/μL 的工作液,置于-20℃冰箱中备用。
1.3 CDDP 引物及PCR 反应体系
21 条 CDDP 引物来自 Collard 等[4]文献报道,均由上海生工生物工程有限公司合成。CDDP-PCR 反应体系为20 μL,包括Taq 聚合酶用量1.5 U、Mg2+浓度 2.0 mmol/L、引物浓度 0.375 μmol/L、dNTPs 浓度 0.3 mmol/L、DNA 模板用量 40 ng。PCR 反应程序为:94℃预变性 3 min;94℃变性 50 s,48.5℃~52.5℃退火 1 min,72℃延伸 2 min,35 个循环;最后 72℃延伸 8 min,4℃保温[5]。
1.4 电泳及检测
取 10 μL 扩增产物,加入 2 μL 6×loading buffer,在1.5%琼脂糖凝胶(含核酸染料)中电泳,以D2000 Marker 作标准分子质量进行对照,在SN-NJ0601凝胶成像系统观察记录并采集图像。
1.5 数据统计与分析
采用Gel-Pro analyzer 凝胶图像分析软件并结合人工识别的方法,对清晰稳定的主带进行统计,在相同迁移位置上有带赋值为“1”,无带赋值为“0”,建立原始数据矩阵。
用Excel 2003 计算总条带数、多态性条带数、多态性条带百分率及多态性信息含量。应用Popgene1.32 软件计算有效等位基因数(Ne)、Nei’s 基因多样性指数(H)、香农信息指数(I)、不同群体间的 Nei’s遗传一致度和遗传距离等,并根据Nei’s 遗传一致度采用非加权成对算术平均法(UPGMA)在Ntsys-pcVer.2.10软件中绘制群体间聚类关系图。应用POWERMARKER Version 3.25 软件根据 Nei’s(1983)遗传距离,绘制个体间聚类图。
2 结果与分析
2.1 CDDP 引物的筛选及多态性分析
采用优化好的大豆CDDP-PCR 反应体系及24个差异明显的大豆品种,从21 条引物中筛选出17条谱带清晰、稳定且多态性好的引物对151 份大豆种质资源进行多态性扩增(图1),共产生120 个条带,其中,多态性条带82 个,多态性条带百分率为68.33%(表1)。每条引物的多态性变幅为33.33%~92.31%,每条引物检测的位点数为3~13 个,平均位点数为7.06 个,平均多态性位点4.82 个;不同引物扩增出的总条带数、多态性条带数差异较大,多态性条带数最多的是引物MADS-2,为12 条,多态性条带百分率为92.31%;多态性条带数和多态性条带百分率最低的是引物WRKY-R1,分别为1 条和33.33%,其他引物多态性条带百分率均高于50%。17 条引物多态性信息含量(PIC) 的变异范围为0.289~0.948,平均为 0.644,表明 CDDP 标记能较好地反映151 份大豆品种的遗传多样性。

图1 引物Myb2 对24 个大豆品种的扩增结果

表1 17 条CDDP 引物对151 个大豆种质的扩增结果及多态性
2.2 不同地理来源大豆品种的遗传多样性分析
由表2 可知,大豆品种群水平上的遗传多样性均高于6 个地理来源群体的平均水平。品种群水平上的有效等位基因数(Ne)、Nei’s 基因多样性指数(H)和香农信息指数(I)分别为 1.740 7、0.414 2 和0.601 2;不同地理来源群体水平上的指标Ne、H、I 分别为1.684 0、0.386 2 和0.565 1。6 个不同地理来源大豆品种的 Ne 变异在 1.637 3~1.738 8;H 是衡量供试品种间遗传多样性最常用的指标,能够反映物种等位基因的丰富度和均匀程度,其变异范围为0.360 2~0.406 3;I 变异范围为 0.527 9~0.586 1;Ne、H、I 指标均表现为其他省区>国外>吉林>辽宁>黑龙江>内蒙古,均以其他省区最高,内蒙古最低。综合比较分析各项遗传多样性参数可以看出,内蒙古和黑龙江大豆品种的遗传多样性较低,国外品种和其他省区大豆具有较高的遗传多样性水平。
2.3 不同地理来源大豆品种群的遗传一致度和聚类分析
从表3 可以看出,6 个大豆地理来源群间的遗传一致度变化范围为0.858 0(国外和其他省区)~0.965 7(内蒙古和黑龙江),平均为0.920 4;群体间遗传距离变化范围为0.034 9(内蒙古和黑龙江)~0.150 1(国外和其他省区),这表明内蒙古和黑龙江的大豆品种遗传关系很近,国外品种与国内品种遗传关系较远,特别是与其他省区类群品种遗传关系最远。
为了进一步揭示不同地理来源大豆群体的分化趋势与亲缘关系,根据大豆品种群间的 Nei’s 遗传一致度,进行UPGMA 聚类分析。从图2 可以看出,遗传相似系数在 0.882 处,6 个品种群可以分为两大类,即国内品种和国外品种,表明参试国内外大豆品种间的亲缘关系较远。进一步细分,国内群在遗传相似系数为0.922 处又可分为3 个亚类,即:黑龙江、内蒙古、吉林品种聚为一类,辽宁、其他省区品种各为一类。聚类结果与地理分布有一定的相关性。

表2 不同地理来源群体遗传多样性分析

表3 基于CDDP 标记的不同地理来源大豆种质群的遗传一致度和遗传距离
2.4 参试大豆种质聚类分析
由图3 可知,供试材料被分为三大类。第Ⅰ类群共3 份材料,占供试材料的2.0%,包括黑龙江的地方品种压破车、吉林的吉农12 和其他省区的中黄13。第Ⅱ类群包括65 份材料,占供试材料的43.0%,该类群又可进一步细分为4 个亚类,其中,第1 亚类有10 份材料,包括其他省区品种6 份,占60.0%,辽宁品种3 份,国外品种1 份,多数为晚熟品种;第2亚类有20 份材料,包括吉林品种16 个,占80.0%,黑龙江、辽宁、内蒙古和新疆品种各1 份,其中,新疆的石大豆1 号为引进的吉林品种;第3 亚类有26 份材料,包括吉林和黑龙江品种各10 个,2 个地区品种占76.9%,内蒙古品种3 个,辽宁品种2 个,国外品种1 个;第4 亚类有9 份材料,包括黑龙江品种6个,占66.7%,吉林品种2 个及新疆品种1 个。第Ⅲ类群共83 份材料,占供试材料的55.0%,该类群又可细分为3 个亚类,其中,第1 亚类有5 份材料,包括吉林品种3 个,内蒙古和辽宁品种各1 个;第2 亚类有28 份材料,包括黑龙江品种17 个,占60.7%,吉林品种4 个,国外、内蒙古、其他省区品种各2 个,辽宁品种1 个;第3 亚类有50 份材料,包括国外品种28 个,占该亚群的56.0%,占国外全部供试材料的87.5%,另外,还有内蒙古品种8 个,占内蒙古全部供试材料的57.1%,黑龙江品种7 个,吉林品种4个,其他省区品种2 个,辽宁品种1 个。

图2 基于CDDP 标记的6 个不同地理来源群体UPGMA聚类关系
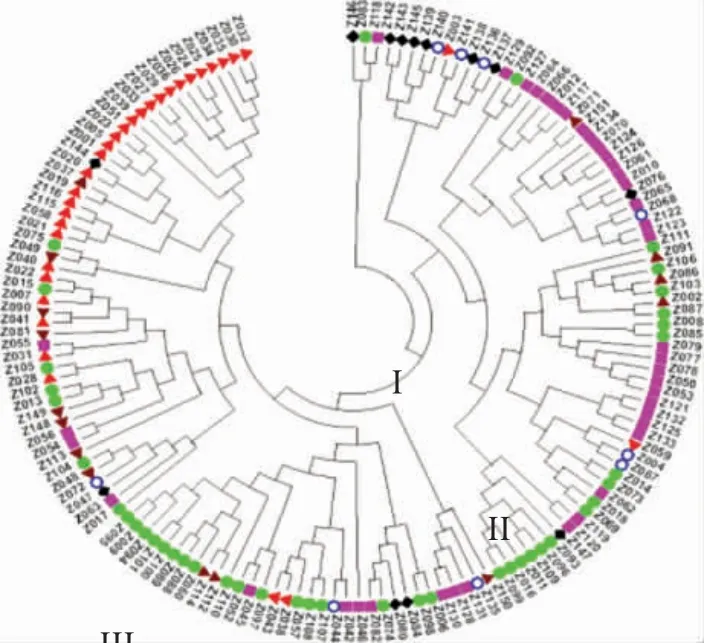
图3 151 份大豆种质UPGMA 聚类结果
由图3 可以进一步看出,国内外品种基本可以分开,国外品种87%以上聚在一起,第Ⅲ类群第3 亚类中28 个国外品种聚在一起;来源相同地区的品种有聚成一类的现象,特别是来自相同育种单位的品种大多都聚在了一起,如合丰50 和合丰47、黑农56和黑农53、吉育57 和吉育67、通农11 和通农14 等聚在了一起;另外发现,内蒙古与黑龙江品种亲缘关系较近,第Ⅱ类群中内蒙古的蒙豆9 号、蒙豆30 号、登科1 号与黑龙江的紫花2 号、黑河38、垦丰20、合丰55 等聚在了一起,原因是它们含有一些共同的亲本,血缘比较近;黑龙江与吉林的种质资源交流较多,因此在不同类群中都有聚在一起的现象,如第Ⅰ类群中的黑龙江的压破车与吉林的吉农12,第Ⅱ类群的牡丰1 号与黄宝珠、黑农44 与吉育72,第Ⅲ类群中的青豆与小金黄等都聚在一起,而白城秣食豆、薄地高、茶色豆、小金黄、青豆等地方品种大多聚为一类。
3 讨论与结论
该研究聚类分析发现,国内外大豆品种基本可以分开,遗传差异较大,关荣霞等[6]通过对中国32 个品种与40 份国外种质的聚类分析发现美国种质可以和中国大豆育成品种区分开来,且国外种质具有更多的特有等位变异,说明中外大豆品种具有较大的遗传差异;魏崃等[7]也认为中国和美国大豆遗传距离较大,存在较大的遗传差异,美国大豆资源可以用来丰富中国大豆的遗传基础,这些结果与该研究结果一致。另外,该研究还发现地理来源相同的品种有聚在一起的趋势,特别是相同育种单位的大豆品种聚在一起,这与张军等[8]的研究结果一致,可能因为相同地区大豆育种者都使用了少数几个优良骨干亲本,而且世代材料选择经验相似,种植的地理生态环境相同等,导致了育成品种间亲缘关系比较近。
CDDP 分子标记技术多态性水平较高,信息含量丰富,可有效揭示大豆种质资源的遗传多样性。聚类分析结果表明,国外与国内大豆品种遗传距离较远,地方品种与育成品种遗传差异较大,可利用国外品种和地方品种拓宽我国大豆的遗传基础。该研究结果可为大豆种质资源科学评价和利用提供理论依据和参考。
