郑州沿黄湿地草本植物群落多样性研究
2021-03-15姚新治王鑫宇胡永歌陈延永何鹏飞田国行
姚新治 王鑫宇 胡永歌 陈延永 何鹏飞 田国行

摘要:以郑州沿黄湿地草本植物为研究对象,分析群落内部组成以及物种多样性,为郑州黄河湿地植物多样性保护提供参考依据。研究采用典型样方法和随机样方法对郑州沿黄湿地18块样地选取了90个样方开展植物现状调查。结果表明,样地内草本植物44科119属154种,以禾本科植物占比最多,其次是菊科、豆科植物;区系由5个分布区类型和1个变型区类型组成,其中世界分布类型和泛热带分布类型最多;草本植物群落多样性随着水分梯度的降低而下降;物种丰富度与Simpson优势度指数、Shannon-Wiener多样性指数、Pielou均匀度指数存在极显著相关性;不同样地之间植物群落生境存在明显差异是群落植物多样性存在差异的主要原因。
关键词:黄河湿地;植物群落;物种多样性;湿地保护
中图分类号:S181文献标志码:A
文章编号:1002-1302(2021)01-0186-06
作者简介:姚新治(1994—),男,河南信阳人,硕士研究生,主要从事景观规划与生物多样性研究。E-mail:862489032@qq.com。
通信作者:田国行,教授,博士生导师,主要从事城市绿地资源建设与调控研究。E-mail:tgh0810@163.com。
湿地拥有大陆、海洋2种生态系统的特性,不仅孕育着大量的动植物,还具备巨大的生态服务价值[1]。植被是湿地景观的主体部分,承载力之下的植物丰富度直接决定着湿地的生态环境[2-3],能够为广大动植物提供良好的生存空间,维持湿地生态系统的稳定性[4-5]。目前国内外学者针对湿地研究多集中在生态修复、景观改造和典型群落等方面[6-9],很少涉及湿地草本植物的研究。郑州段黄河凭借着优良的气候、土壤、水文等生态条件,已经成为了郑州市的生态屏障,是郑州市的后花园。但随着郑州大都市区建设,郑州黄河湿地遭受到前所未有的生态扰动,植被破坏严重。本研究以郑州黄河沿岸草本植物为研究对象,探索其草本植物群落多样性及特征,为郑州黄河湿地生物多样性保护及恢复提供参考。
1材料与方法
1.1研究区域概况
郑州沿黄湿地坐标位置在112°48′~114°14′E、34°48′~35°00′N之间。研究区地势西高东低,呈现明显差异,最低海拔75m,最高海拔277.7m,气候属北温带大陆性季風气候,平均气温15.7℃,最高气温42.5℃,最低气温-10.2℃。7月最热,月平均气温27.9℃;1月最冷,月平均气温11℃[10-11]。年均降水量641mm,多集中于夏季6—7月,12月至次年1月的降水量相对较少。
1.2样地调查
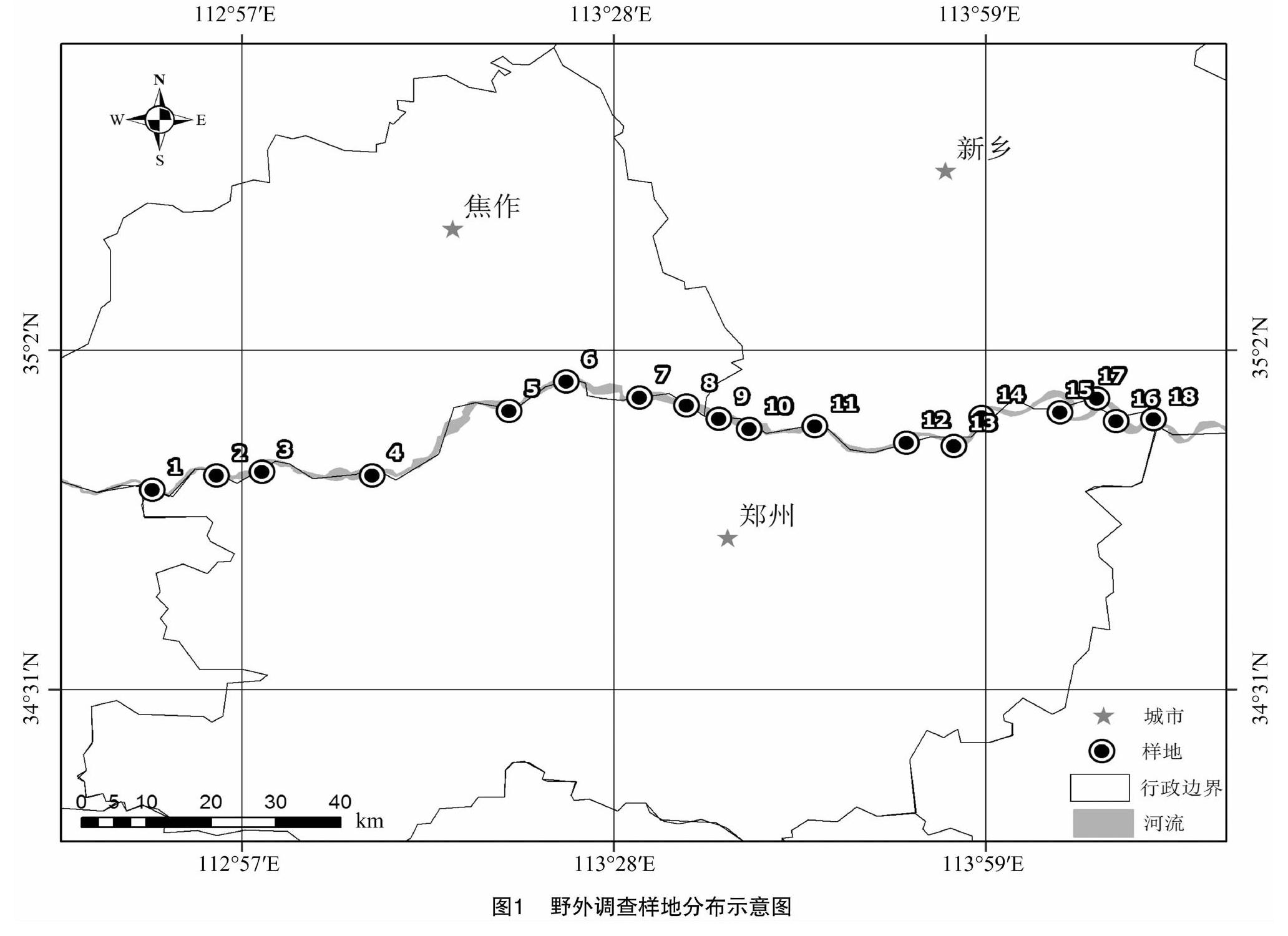
2019年6—7月,采用典型样方和随机样方法对郑州沿黄湿地进行湿地草本植被调查,沿水流方向自西向东,以水陆交界处为样地水平线,垂直河流方向为延伸线,约每10km设置1块样地,包含郑州黄河北岸大堤2块样地,共设置18块样地(图1),每块样地按河流方向和垂直河流方向设置5个面积为1m×1m的样方,记录90个样方内植物的物种、高度、株丛数、盖度、多度,并记录该样方地的经纬度、海拔高度等基本状况,以便于计算出各种植物的相对盖度、相对高度、相对多度和重要值等。
1.3数据处理
1.3.1重要值计算
重要值是指某一物种在群落中的重要程度或优势度的指标[12],它与种群的数量、多度、盖度等相挂钩。通过植物的重要值可以判定物种在植物群落内的重要性及其地位,并确定群落物种的群系和组成结构。本次调查中主要依据重要值确定物种的优势程度,即重要值高的物种为相对优势种。重要值的计算公式[13]如下:
重要值(IV)=(相对频度+相对盖度+相对高度)/3。
式中:相对盖度=某个物种的盖度/所有物种的总盖度×100%;相对高度=某个物种的高度/所有物种的总高度×100%;相对频度=某个物种的频率/所有物种的总频率×100%。
1.3.2物种多样性测度物种多样性的研究方法有很多种,既能用植被的盖度和植物数量为计算标准,也可以用植物重要值为计算指标[14]。该研究使用的是以重要值为计算指标的研究方法,此方法适用于草本植物群落。目前国内外普遍利用物种丰富度指数(S)、Simpson优势度指数(D)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(E)进行植物物种多样性的分析。多样性指数的公式如下:
式中:S为各样地的植物物种总数;Pi=Ni/N,表示物种i的相对重要值,N为各样地中所有植物物种的重要值之和,Ni为样地中第i个物种的重要值。
本研究以群落丰富度指数、优势度指数和均匀度指数为因变量,分析S、D、E和H之间的相关性,通过单因素方差分析(ANOVA)对相关指数的差异进行检验[15]。
2结果与分析
2.1物种组成分析
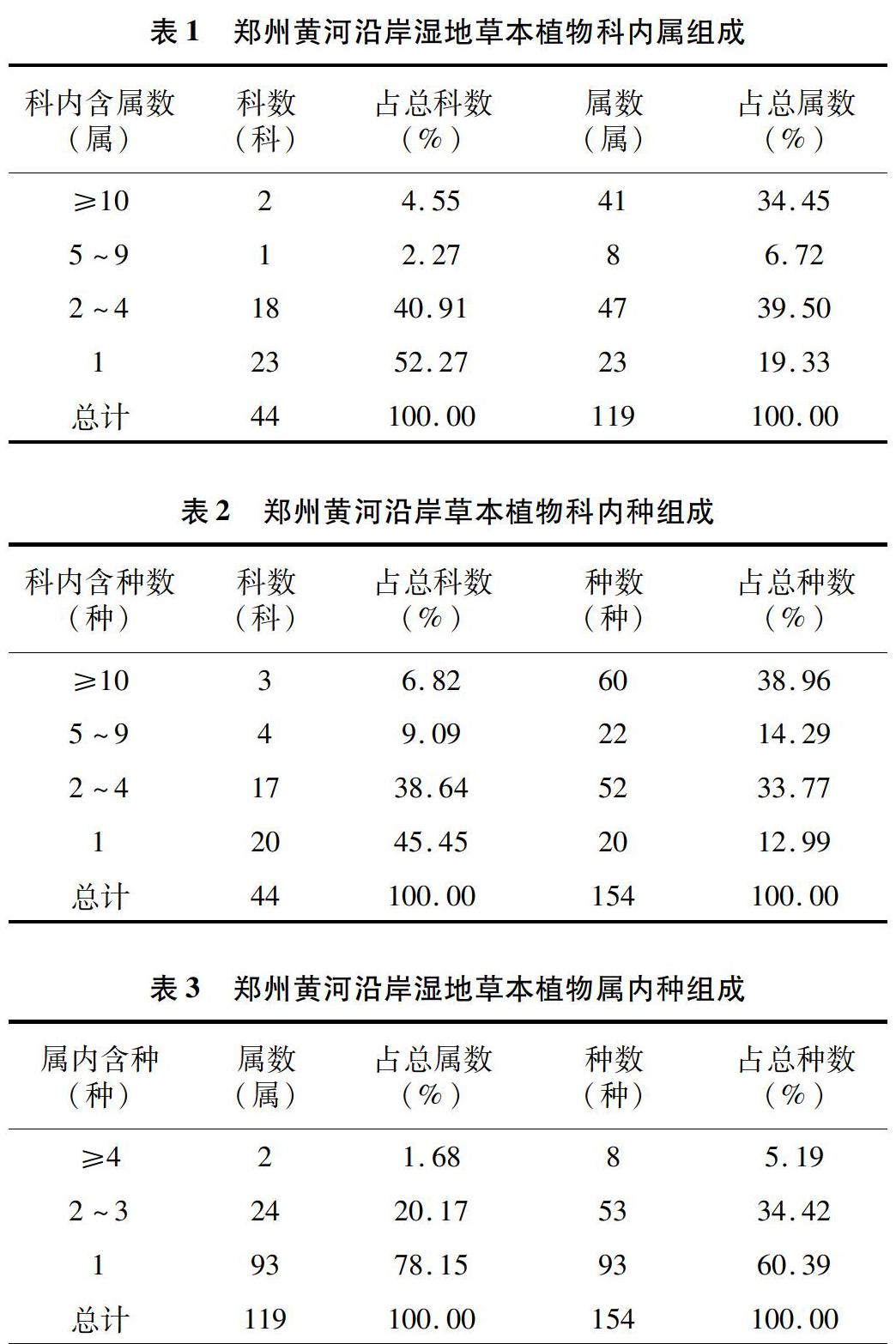
由表1可知,郑州黄河沿岸湿地草本植物共计44科119属154种。按照科的大小统计,其中科内含10属以上的有2科,占总科数的4.55%,共计41属,其中禾本科23属、菊科18属,占总属的34.45%;含5属以下的有41科,占总科数的93.18%,共计70属,占总属数的58.83%。根据科内种的分析(表2),含5种及以上的科有禾本科、豆科、藜科、莎草科、苋科等7科,占总科数的15.91%,共包含植物82种,占总种数的53.25%,含2~4种的科有车前科、唇形科、大戟科、蔷薇科、茄科、伞形科等17科,占总科数的38.64%,共计包含植物52种,占总种数的33.77%;还有葫芦科、堇菜科、无患子科、茜草科等20种单科植物。
按照属的大小统计(表3),含4种及以上的属仅有2属,分别是蒿属、蓼属,占总属数的1.68%;含2~3种的属有苍耳属、苋属、蓟属、稗属、蛇葡萄属等24属,占总属数的20.17%,共计包含了53种草本植物,占植物总数的34.42%;属内仅含1种草本植物的属有93属,占总属数的78.15%。可以看出植物属的组成具有极强的分散性,各属之间的物种数差异小,单物种属数最多,说明了郑州沿黄湿地草本植物组成比较复杂[16]。
2.2植物区系分析
根据吴征镒《世界种子植物科的分布区类型系统》[17],将郑州黄河沿岸湿地草本植物44科划分为5个分布区类型和1个变型区类型(表4),区系类型主要集中分布在世界分布、泛热带分布、北温带分布类型,其中世界分布的有禾本科、菊科、莎草科、车前科、蔷薇科等23科,占总科数的52.27%,共包含82属107种植物,占植物总数的69.48%;泛热带分布的有豆科、旋花科、大戟科等12科,包含了26属33种植物,占植物总数的21.42%;北温带分布的有桑科、蝶形花科等5科,包含8属10种植物,占植物总数的6.49%。
2.3重要值分析
依据郑州沿黄湿地18块样地的样方资料,统计分析了植物群落草本物种重要值,每个样地列出重要值最高的3种(表5)。由表5可以看出,郑州黄河沿岸湿地草本植物群落结构复杂,植物的重要值跨度较大。狗牙根、马唐等物种在多数群落中占有较大的优势,这与狗牙根等茎型禾草具有根状茎、耐性强的特点有关[18]。从群落变化来看,随着水分梯度的变化,群落内优势种变化差异明显,自西向东优势群落由白茅+醴肠群落向莎草+蒲苇群落再向狗牙根+马唐+狗尾草群落变化,这与植物的自身耐性以及周围环境相适应的结果有关。由各样地群落优势种分布来看,中生环境和旱生环境植物占据了很大的优势,表明了郑州沿黄湿地植被正由湿生植被向旱生植被演替过渡。
2.4植物多样性分析
由图2可以看出,物种丰富度指数S介于12~31之间,样地5的丰富度指数最大(31),样地14的丰富度指数最小(12),物种丰富度指数差距明显。这是由于样地5位于连接河道的洼地处,无人为干扰且水分充足,形成了小型的湿地生态系统;而样地13、14、15紧邻中牟渔业养殖基地,周边坑塘比例不断扩张,基塘比失衡,导致植物生长受限。17、18这2个样地位于郑州黄河北岸的大堤,堤上生活着大量未搬迁的居民,人为干扰是此地物种丰富度较低的主要原因。
Shannon-Wiener多样性指数H在2.4639~2.8621之间浮动,H指数越高,表明植物多样性越高。由图2可以看出,样地5最高,达3.2221,样地15最低,为2.1967,样地13、14、15、17、18都在2.4以下。由此可见,各样地植物群落的物种多样性指数总体偏低,且存在一定的差异性,也反映出该研究区植物群落结构简单且种间差异性大。
Simpson优势度指数(D)从生态学的角度来说,它可以反映出具有优势的植物在其群落内部所占据的位置和其发挥的作用重要性的高低[19]。由图2可以看出,各个优势度指数D值介于0.8416~0.9527之间,样地5最高,为0.9527;样地15最低,为0.8416;样地13、14、15、17、18的优势度较低,均在0.88以下,其次是样地1、8、11优势度低于0.9,其余样地均在0.9以上。样地3的S和H值都大于样地2,但其D值低,说明样地3群落内物种虽然丰富,但群落内部存在的不均匀性大于样地2,根据均匀度指数E的结果显示,样地3(E=0.8796)低于样地2(E=0.9321),与均匀度指数相符合。
由图2可以看出,各样地植物群落的Pielou均匀度指数(E)在0.9上下浮动,其中浮动最大的是样地4和15,样地4最高,为0.9532,样地15最低,为0.8323。总的来看,各群落的均匀度相对比较高,说明群落中各植物的分布较均匀。
总的来看,郑州沿黄湿地草本植物群落丰富度指数S与D指数、H指数、E指数自西向东均呈现递减的趋势,变化格局一致。各个群落4个指数的相关性分析(表6)显示,郑州沿黄湿地草本植物群落多样性与丰富度指数、优势度指数、均匀度指数相关线性表示为H=0.052S+1.664,r=0.937,n=18(P≤0.01);H=8.891D-5.452,r=0.934,n=18(P≤0.01);H=4.957E-1.935,r=0.591,n=18(P≤0.01)。这说明研究区内物种多样性指数H与S、D、E均存在极显著相关性关系。
3结论与讨论
郑州沿黄湿地草本植物44科119属154种,科属分化程度高,植物组成复杂且具有明显的气候性特征,有由热带向温带特征过渡的趋势,这与郑州的自然地理环境相符合[9]。从郑州黄河沿岸湿地植物组成可以得知,中生性植物具有绝对的优势,其次是具有高强度耐性的旱生植物,湿生和水生植物有退化的倾向。狗牙根、小飞蓬、白茅、狗尾草、野大豆、马唐、葎草等耐性较强的植物在保护区内是占比较大的优势种,湿地植物芦苇、莎草、稗草、香蒲等能够涵养水源的湿生植物相对劣势。这是由于黄河下游河床侵蚀下切严重,引起黄河水位降低[20-21],造成水土流失、湿地面积減小的局面,耐性强的中生环境植物具有较强的适应性,因此生长不受限制。各样地间不同程度的水淹对土壤含水量、微元素含量以及各种营养物质都能造成影响,从而对植物产生水分上的筛选,产生了优势物种与优势群落,这与郭雪莲等研究认为水分梯度是影响湿地植物分布的主要原因结论[22]一致。
郑州沿黄湿地草本植物群落具有一定的稳定性和复杂性。各群落优势种明显,占据主导地位,适应能力强,更能维持群落的稳定性,但各个群落内部的优势种重要值差也呈现出2种不同的趋势。样地1~8群落内优势种重要值相差较小,如样地6植物群落3种优势种中,狗牙根的重要值为13.60%,蒲苇的重要值为9.43%;样地9~18群落内部优势种重要值相差较大,比如样地15马唐的重要值为30.99%,而藜的重要值只有5.82%;同种物种在不同样地植物群落中的优势程度也不同,如狗牙根在样地8的重要值为21.54%,而在样地13的重要值为7.51%。造成以上现象的原因一方面是由于植物本身的世界广布和泛热带分布的特性,另一方面是由于各个样地距离城市的距离不同,导致人为干扰程度也各不相同。样方1~8分布在巩义、荥阳距离城市较远的地方,开发建设相对城市较低,植物生长相对自然;而样地9~18距离城市较近且海波低,多为平原,人类活动多,在人类高强度干扰下适应能力强的植物生长较好,适应能力差的植物生长受到影响,因此各种植物优势程度会呈现两极化现象。根据调查可知,样地所在区域环境对物种的分布和重要值有所影响,导致不同地点植物群落构成有所差别,主要原因是样地之间生境条件差异大造成的,这与张志铭等的结论[7,23-24]相同。
郑州沿黄湿地草本植物丰富度指数S与H指数、D指数、E指数呈现相同的变化趋势,即随着土壤水分梯度的减小、人为干扰程度的增大等阻生因子的加强,植物多样性逐渐降低。研究区内草本植物的H指数与S指数、D指数、E指数存在极显著相关性,而赵海莉等研究表明植物多样性指数与丰富度指数无显著相关性关系[25]。产生差异的原因是前者研究区域物种丰富度介于0~3之间,丰富度指数较小,差异也较小,降低了丰富度指数对物种多样性水平的影响;而郑州黄河沿岸不同样地之间丰富度指数差距较大,对物种多样性水平影响较大,能直观地反映出各个样地的物种多样性水平。
在调查的过程中发现,由于黄河水量逐年减小,土地沙化严重,沿岸植被正由湿生植物向旱生植物演替;滩区土地无序开垦比较严重,周围居民乱砍滥伐,植被遭到严重破坏,导致黄河沿岸植物多样性下降,呈现出湿地不湿、湿地不净、湿地不美的局面。因此,保护黄河生态安全,维护生物多样性功能是构建黄河流域生态保护和高质量发展的首要任务。首先要提升湿地水源涵养能力,增加地下水位养护湿地,构建分级的滩区生态安全格局[26],加强黄河滩区综合治理,合理开发滩区土地,实行集中统一管理;其次构建不同水分梯度的植物种群,加强植物群落的稳定性,严格控制外来植物种群对本土植被和湿地环境的破环;同时加大教育宣传,提升周边群众对黄河湿地的生态保护意识,共创美丽黄河。
参考文献:
[1]刘俊娟.丹江湿地植物多样性特征及其环境影响因素[J].西南农业学报,2017,30(12):2811-2819.
[2]李有志,崔丽娟,潘旭,等.辽河口湿地植物多样性及物种功能型空间分布格局[J].生物多样性,2015,23(4):471-478.
[3]谢冬明,黄庆华,易青,等.鄱阳湖湿地洲滩植物梯度变化[J].生态学报,2019,39(11):4070-4079.
[4]ZhangYR,WangRQ,KaplanD,etal.Whichcomponentsofplantdiversityaremostcorrelatedwithecosystemproperties?AcasestudyinarestoredwetlandinnorthernChina[J].EcologicalIndicators,2015,49:228-236.
[5]胡建成,古岐川,王瑜,等.金马河流域滩涂湿地植物多样性特征[J].四川农业大学学报,2018,36(4):450-456.
[6]陈明林,文慧慧,汪小平,等.安徽秋浦河源湿地植物物种多样性研究[J].湿地科学,2014,12(3):369-373.
[7]张志铭,张璐,叶永忠.郑州黄河湿地自然保护区植被群落物种多样性调查[J].河南科技大学学报(自然科学版),2010,31(2):81-85.
[8]张瑜,吴永华,赵峰.黄河兰州段湿地物种多样性及保护研究[J].草原与草坪,2014,34(6):56-61.
[9]赫晓慧,郑东东,郭恒亮,等.郑州黄河湿地自然保护区植物物种多样性对人类活动的响应[J].湿地科学,2014,12(4):459-463.
[10]程炳岩,朱业玉,王记芳.郑州城市气候环境的观测研究[J].气象,2004,30(2):50-54.
[11]李长看,马灿玲,王威,等.郑州黄河湿地生物资源及保护研究[J].农业科学与技术(英文版),2010,11(6):159-163.
[12]游水生,张志翔,李如泽,等.福建武平帽布米槠林火烧后植物种类变化的研究Ⅱ.火烧前后重要值和物种多样性变化[J].福建林学院学报,1998,18(1):65-68.
[13]王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报(自然科学版),2010,33(2):312-316.
[14]陈飞鹏,暨淑仪,汪殿蓓,等.海南南红稻田杂草群落物种多样性指数的测定及评价[J].华中农业大学学报,2001,20(5):438-441.
[15]张起鹏,张丽,张旭,等.兰州城市湿地植物多样性[J].安徽农业科学,2011,39(34):21195-21197,21269.
[16]张金铭,符龙飞,洪欣,等.石首麇鹿国家级自然保护区草本植物区系及物种多样性[J].生态学杂志,2019,38(2):513-520.
[17]吴征镒.种子植物分布区类型及其起源和分化[M].昆明:云南科技出版社,2006.
[18]張起鹏,王倩,张丽,等.黄河兰州段湿地植物多样性研究[J].水土保持通报,2012,32(3):240-244.
[19]张峰,上官铁梁.山西南方红豆杉(Taxusmairei)森林群落的生态优势度分析[J].山西大学学报(自然科学版),1988(3):82-87.
[20]梁少民,罗晓洁,孟祥宇,等.河南省沿黄湿地草本植物群落多样性特征分析[J].人民珠江,2018,39(8):9-13.
[21]王婷,马怀宝,陈书奎,等.2007年调水调沙小浪底水库异重流排沙分析[J].人民黄河,2008,30(12):54-56.
[22]郭雪莲,吕宪国,戴国华.三江平原不同水位梯度湿地地上生物量动态特征[J].生态环境,2008,17(5):1739-1742.
[23]杨文斌,刘坤,周守标.安徽新安江干流滩涂湿地草本植物区系及物种多样性[J].生态学报,2013,33(5):1433-1442.
[24]赵婧,祝遵凌.江苏盐城原生滨海湿地土壤细菌群落多样性分析[J].江苏农业科学,2019,47(2):258-261.
[25]赵海莉,赵锐锋,张丽华,等.黑河中游湿地典型植物群落特征与物种多样性[J].生态学杂志,2013,32(4):813-820.
[26]张明祥,张阳武,朱文星,等.河南省郑州黄河自然保护区湿地恢复模式研究[J].湿地科学,2010,8(1):67-73.
