短时高低温胁迫对南亚实蝇生长发育及繁殖的影响
2021-03-12黄禹禹顾祥鹏彭孝琴陈国华齐国君张晓明陶玫
黄禹禹 顾祥鹏 彭孝琴 陈国华 齐国君 张晓明 陶玫

摘要 :南亞实蝇Bactrocera tau (Walker)是一种重要的入侵性农业害虫。主要分布在我国云南、广东、四川和河南等地,南亚实蝇的为害严重降低了蔬菜和水果的品质。短时极端温度是影响南亚实蝇种群的重要因子。为了明确短时高低温对南亚实蝇的影响,通过选择12 h的短时低温(-4、0、4、8℃)及12 h的短时高温(34、38、42、46℃),在实验室条件下系统研究短时高低温胁迫对南亚实蝇种群生长发育和繁殖的影响。结果表明,在短时高/低温处理后,南亚实蝇各虫态的存活率随着处理温度的升高/降低逐渐下降;随着处理短时高/低温的升高/降低,南亚实蝇发育历期、产卵前期和后代性比逐渐增加;随着短时高温处理温度的升高,雌雄成虫的寿命先延长后缩短,单雌产卵量逐渐减少;随着短时低温处理温度的降低,雌雄成虫的寿命逐渐缩短,单雌产卵量先增加后减少。短时高温和短时低温均对南亚实蝇的生长发育和繁殖有着不利的影响,但一定的低温(8℃)处理后对雌成虫繁殖力有一定的促进作用。
关键词 :南亚实蝇; 高温; 低温; 生长发育; 繁殖
中图分类号:
S 433文献标识码: A
DOI: 10.16688/j.zwbh.2020113
Effects of shortterm high and low temperature on the development and reproduction of Bactrocera tau (Walker)
HUANG Yuyu1, GU Xiangpeng1, PENG Xiaoqin1, CHEN Guohua1,QI Guojun2, ZHANG Xiaoming1*, TAO Mei1*
(1. College of Plant Protection, Yunnan Agricultural University, National Key Laboratory for Conservation and
Utilization of Biological Resources in Yunnan, Kunming 650201, China; 2. Guangdong Provincial Key
Laboratory of High Technology for Plant Protection, Research Institute of Plant Protection,
Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China)
Abstract :Bactrocera tau (Walker) is an important agricultural invasive insect pest. It is mainly distributed in Yunnan, Guangdong, Sichuan and Henan provinces of China. The serious damage of B.tau can reduce the quality of vegetables and fruits. The shortterm extreme temperature is an important factor affecting the population of B.tau. In order to determine the effect of shortterm high and low temperature on the growth, development and reproduction of B.tau, the effects of shortterm low temperature (-4, 0, 4, 8℃) for 12 hours and shortterm high temperature (34, 38, 42, 46℃) for 12 hours were studied under laboratory conditions. The results showed that the survival rates of all stages gradually decreased with the increase and decrease of the treatment temperature after shortterm high and low temperature treatments. The developmental period, preoviposition period, single female oviposition, longevity and the sex ratio of offspring were significantly affected by the treatments. The developmental period, sex ratio and preoviposition period gradually increased with increasing/decreasing temperature; the longevity of male and female adults initially increased and then decreased, while the single female oviposition gradually decreased with increasing treatment temperature. Meanwhile, their longevity gradually decreased and the single female oviposition increased and then decreased with decreasing treatment temperature. Overall, the shortterm high and low temperature had negative effects on the growth, development and reproduction of B.tau. This could slightly promote the reproduction of females in a certain range of shortterm low temperature (8℃).
Key words :Bactrocera tau; high temperature; low temperature; growth; development; reproduction
南亚实蝇Bactrocera tau (Walker),隶属双翅目Diptera实蝇科Tephritidae寡鬃实蝇属Dacus Fabricus,1992年寡鬃实蝇属被修订为果实蝇属Bactrocera Macquart,南亚实蝇也称为南亚果实蝇和南亚寡鬃实蝇,俗名瓜蛆、蹦蹦虫、黄蜂子。南亚实蝇于1849年在福建省首次报道,但在很长一段时间内没有造成较大为害[1]。至今,南亚实蝇已在亚洲的热带和亚热带以及南太平洋地区有分布,近年在我国随着水果蔬菜等农产品的频繁贸易和调运,迅速扩散,目前为止已经在云南、广东、四川和河南等多个省分布和为害[25]。南亚实蝇以雌成虫产卵于果实内,卵孵化为幼虫后取食果肉组织,被害后的组织在一定条件下会变畸形或直接腐烂,轻者降低果实的品质和产量,重者造成绝收,给农业生产带来了巨大的损失[68]。南亚实蝇危害大,扩散快的主要原因之一就是它的寄主范围广,目前报道的寄主植物种类达16科91种,包括葫芦科、杜英科和茄科的多种重要农作物[910]。南亚实蝇的为害给多种农作物造成较大的经济损失,如在重庆地区5月-9月南亚实蝇对南瓜的为害率最高可达73.10%[4];对海南省瓜菜种植园中的葫芦科、茄科、番木瓜科和西番莲科等作物均能造成严重危害和减产[11]。此外,南亚实蝇的大量野生寄主种类能促进南亚实蝇的扩散和为害,导致防治困难[1213]。在南亚实蝇发生的地区,其成虫常年可见,且一年内有两次高峰期,分别为1月-3月和6月-7月,且夏季的种群数量大于春季[14]。
昆虫属于典型的变温动物,其生长发育易受到温度的影响[1516]。当环境温度超出昆虫正常生长发育的适温范围,即使是短暂的也会对昆虫的生长或存活造成巨大影响甚至导致种群灭亡[16]。依据温度变化高低及作用时间的长短,可将温度变化对昆虫的影响分为长时高低温胁迫和短时高低温胁迫,由于胁迫类型的不同,对昆虫产生的影响也不一致[17]。在气候变暖的趋势下,高温的范围、频率和持续时间持续增加[1820]。过去几十年以来,全球平均气温呈逐步上升趋势[21],
与持续性有规律的温度变化相比,早春天气不稳定,日气温往往与正常气温相差甚远,温度的突然短暂变化对昆虫的生长发育和繁殖可能产生更大影响。在短时高温处理后,亚洲玉米螟 Ostrinia furnacalis卵的孵化率、幼虫的化蛹率、蛹的羽化率和成虫存活率都随处理温度的升高而逐渐降低[22];徐天杨的研究表明,玉米螟赤眼蜂Trichogramma ostriniae经过42℃的短时高温处理3 h和4 h后,雌蜂的寿命显著缩短[23];罗敏等研究表明在经过2 h的2~14℃的低温胁迫后,广聚萤叶甲Ophraella communa除蛹以外的其他虫态的发育及存活率受到显著影响,且成虫寿命及产卵量也显著降低[24];当东亚飞蝗Locusta migratoria manilensis (Meyen) 受到≤5℃的低温胁迫时,其各虫态的取食行为基本停止,当处理温度为0℃以下时,1龄幼虫只能存活18 h[25]。南亚实蝇种群增长的最适温区为25~31℃,在此温度区间内其种群增加最快,若外界温度偏离了南亚实蝇的最适温区,则会对南亚实蝇的繁殖和发育带来积极或消极的影响,从而影响到南亚实蝇种群的发展[26]。
在我国多数地区,南亚实蝇全年均有发生,必然会经受到由于初春气候的不稳定及夏季温度的骤升而出现偏离正常温度较大的短时高低温,这样不正常的短时低温和高温可能对南亚实蝇的种群造成影响。本研究在实验室模拟以上情景,研究了12 h的短时低温(-4、0、4、8℃)胁迫及12 h的短时高温(34、38、42、46℃)胁迫对南亚实蝇生长发育和生殖的影响。为探讨在早春和夏季的田间控制南亚实蝇种群提供基础,最终为南亚实蝇的田间综合防治提供参考。
1 材料与方法
1.1 试验材料与温度设定
供试南亚实蝇于2017年采自云南省红河州蒙自市南郊菜园,后置于养虫室中,在温度(24±1)℃、RH(70±5)%条件下继代饲养5代。试验中的短时高低温处理均在人工气候箱及冰箱中进行,RH(70±5)%,光周期L∥D=14 h∥10 h。处理的温度设定为-4、0、4、8、24、34、38、42、46℃,其中对照组温度为(24±1)℃,0℃以下的短时低温试验在冰箱进行。每个处理设4个重复,每个温度处理12 h。处理后立即转入养虫室,在温度(24±1)℃、RH(70±5)%条件下饲养。
试验仪器:BIC300人工气候箱,上海博迅医疗设备厂。SZX7体视显微镜,日本奥林巴斯株式会社。BCD201STPA冰箱,海尔集团。
西葫芦Cucurbita pepo和南瓜C.moschata均购买自沃尔玛超市的有机食品区,洗净晾干后保存于4℃冰箱冷藏室中备用。
1.2 试验方法
1.2.1 短时高低温对卵、幼虫和蛹生长发育的影响
选取1日龄卵为试验材料,每组10粒卵,为1次重复,放置于事先准备好的1 cm厚西葫芦切片上,并置于7.5 cm培养皿中。将盛有卵的培养皿放置于人工气候箱中进行短时高温或短时低温处理处理12 h,0℃以下的短时低温处理在冰箱进行。处理后的卵立即放入养虫室进行饲养,每隔24 h利用体视显微镜观察记录卵的发育进度及存活情况,直至供试卵孵化或死亡(卵发黑变色或失水干瘪视为死亡)。每个温度为1个处理,每个处理设置4次重复(即4组卵)。
选取1日龄幼虫作为试验材料,每组10头幼虫,为1次重复,放置于切成4 cm厚成块的西葫芦上,后放入特制的纸杯中,纸杯中含有人工饲料和带有幼虫的西葫芦,纸杯放入透明玻璃罐中,用纱网袖套封口。将盛有幼虫纸杯的玻璃罐放入提前开启设置好的人工气候箱进行短时高温或短时低温处理处理12 h,0℃以下的短时低温处理在冰箱进行。处理后的幼虫立即放入养虫室进行饲养,定时更换补充西葫芦确保幼虫有充足的食物,同时观察幼虫发育进度、存活情况及其发育历期。幼虫即将化蛹时,将纸杯置于底部铺有3 cm厚细沙土的沙盆里,西葫芦上盖一浸湿3层的纱布补充水分,每隔24 h观察记录1次幼蟲的化蛹情况,直至供试幼虫化蛹或死亡[24]。每个温度为1个处理,每个处理设置4次重复。
选取1日龄的蛹为试验材料,每组为10头蛹,为1次重复,将蛹放置于铺有浸润滤纸的直径7.5 cm培养皿中,将培养皿放入透明玻璃罐中,玻璃罐口用纱网袖套封口。将盛有蛹的玻璃罐放入提前开启设置好的人工气候箱进行短时高温或短时低温处理处理12 h,0℃以下的短时低温处理在冰箱进行。处理完成后立即将蛹放入养虫室,定时喷水补充水分,同时每隔24 h观察记录蛹的发育进度、存活情况及其发育历期,直至供试蛹羽化或死亡(蛹发黑变色或干瘪破损,则视为死亡)。每个温度为1个处理,每个处理设置4次重复。
1.2.2 短时高低温对成虫存活、繁殖、寿命及后代性比的影响
选取24 h内羽化的同一批成虫作为试验材料。把1头雌虫与1头雄虫放置于1个透明玻璃罐中,并用2个7.5 cm培养皿分别盛放水和雀巢果珍粉供其补充水和食物,盛水的培养皿内放置脱脂棉,在其上铺盖湿润滤纸,防止实蝇溺亡。玻璃罐用纱网袖套封口,便于添加水和食物,将盛有成虫的玻璃罐放入提前开启设置好的人工气候箱进行短时高温或短时低温处理处理12 h,其中的0℃以下短时低温处理在冰箱进行。处理完成后将成虫立即放入养虫室,每罐每日放入1块1 cm3的南瓜块用来收集卵,每隔24 h更换1次并观察记录产卵前期、日产卵数、单雌产卵量和成虫寿命,直至供试成虫全部死亡。将每日收集到的卵分组放入养虫笼中,并以西葫芦饲养,待成虫羽化后分别统计不同处理下成虫的后代雌性比。每10对南亚实蝇为1个重复,每个温度下的处理设置4次重复。
1.2.3 计算公式
相关参数计算公式如下:
校正死亡率=[(处理死亡率-对照死亡率)/(1-对照死亡率)]×100%[27];
校正存活率=1-校正死亡率[27];
某虫态的发育历期=(Σ存活的某虫态的发育历期)/某虫态存活数量[2829];
LT50(半致死温度):某一特定的处理时间内,受试动物死亡一半时所需要的温度。
S(x)=[exp(a-bx)]/[1+exp(a-bx)]
式中:S(x)为昆虫种群在一定时间或低温下的死亡率(%),x为温度T,a和b为模型参数。当死亡率S(x)为50%时,即a-bx=0,x=a/b,x值即致死中温度(LT50)[30]。
1.2.4 数据处理
数据分析采用SPSS 22.0软件,使用Duncan氏法在α=0.05水平上比较不同虫态对不同温度耐热差异显著性,不同性别成虫在同一温度寿命差异显著性通过t检验(P<0.05)进行分析[28]。
2 结果与分析
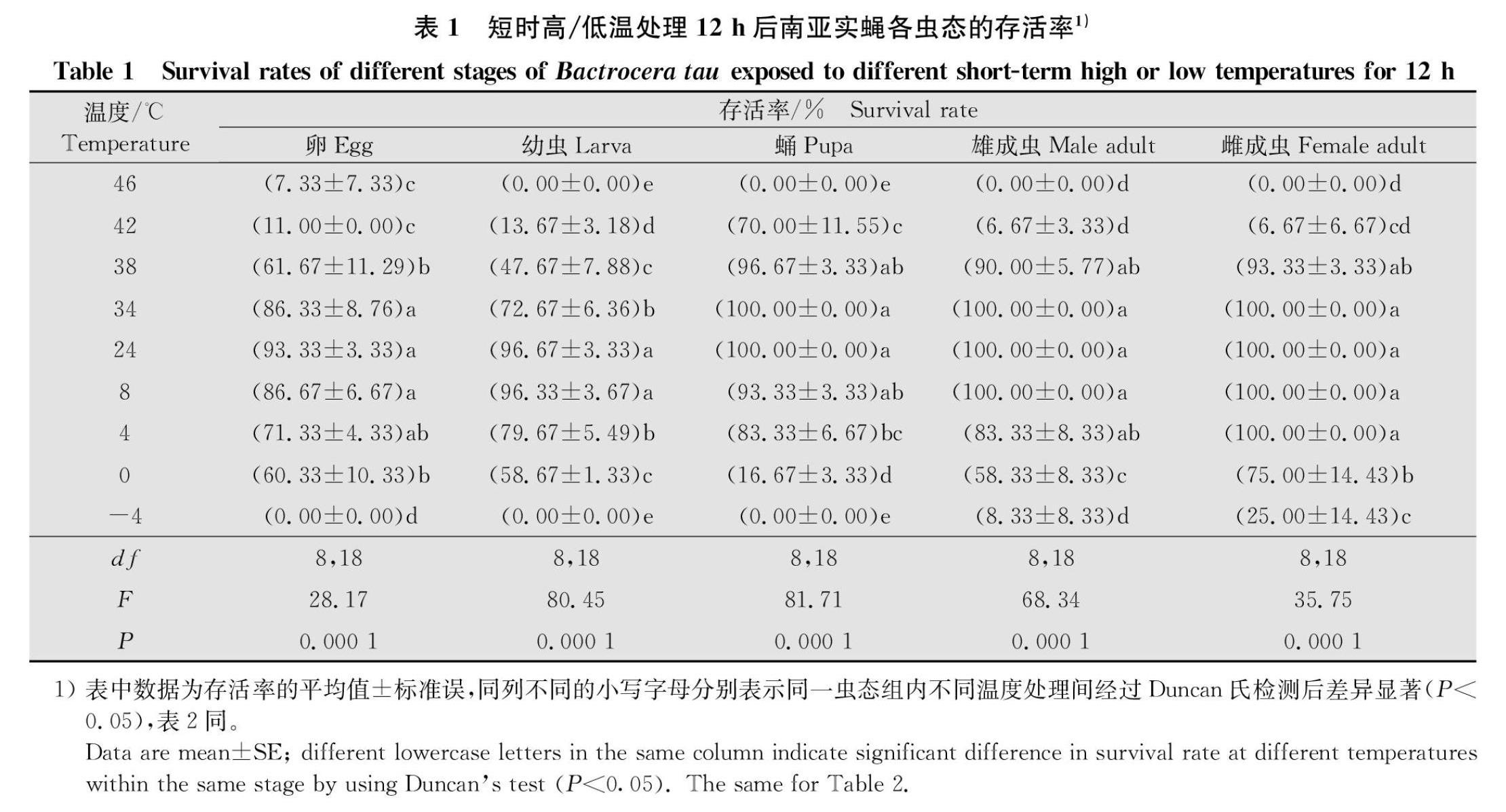
2.1 短时高/低温度处理对南亚实蝇不同虫态存活率的影响
不同短时高/低温度处理对不同南亚实蝇虫态的存活率有着显著的影响(卵:F8,18=28.17,P=0.000 1;幼虫:F8,18=80.45,P=0.000 1;蛹:F8,18=81.71,P=0.000 1;雌成虫:F8,18=35.75,P=0.000 1;雄成虫:F8,18=68.34,P=0.000 1)(表1)。短时高温处理后,卵、幼虫、蛹和雌雄成虫的存活率随着处理温度的升高逐渐降低,当处理温度上升至42℃时,蛹和雌雄成虫的存活率显著低于对照温度下的存活率(蛹:F1,4=7.808 0,P=0.049 1;雌:F1,4=196.000 0,P=0.000 2;雄:F1,4=784.000 0,P=0.000 1),且蛹的存活率仍达到70.00%,但雌雄成虫的存活率仅为6.67%;卵
在38℃和幼虫在34℃时的存活率分别
显著低于对照(卵:F1,4=7.977 0,P=0.047 6;幼虫:F1,4=11.172 0,P=0.028 8),且存活率分别为61.67%和72.67%。在短时低温处理后,随着处理温度的降低,卵、幼虫、蛹和雌雄成虫的存活率逐渐降低。当处理温度下将至0℃时,卵和雌雄成虫的存活率为60.33%,75.00%和58.33%,均显著低于对照温度下的存活率(卵:F1,4=9.238 0,P=0.038 4;雌:F1,4=8.308 0,P=0.044 9;雄:F1,4=25.000 0,P=0.007 5);在-4℃处理温度下,雄雌成虫仍有较低的存活率,存活率分别为8.33%和25.00%;4℃低温下,幼虫和蛹的存活率显著低于对照温度下的存活率(幼虫:F1,4=8.151 0,P=0.046 2;蛹F1,4=9.308 0,P=0.038 0)(表1),分别为79.67%和83.33%。综合不同短时高低温胁迫下南亚实蝇各虫态的存活率结果表明,随着处理温度的升高/降低,南亚实蝇各虫态的存活率均降低,在极端高温46℃下,幼虫、蛹和雌雄成虫均不能存活;在极端低温-4℃时,仅雌雄成虫有较低的存活率。
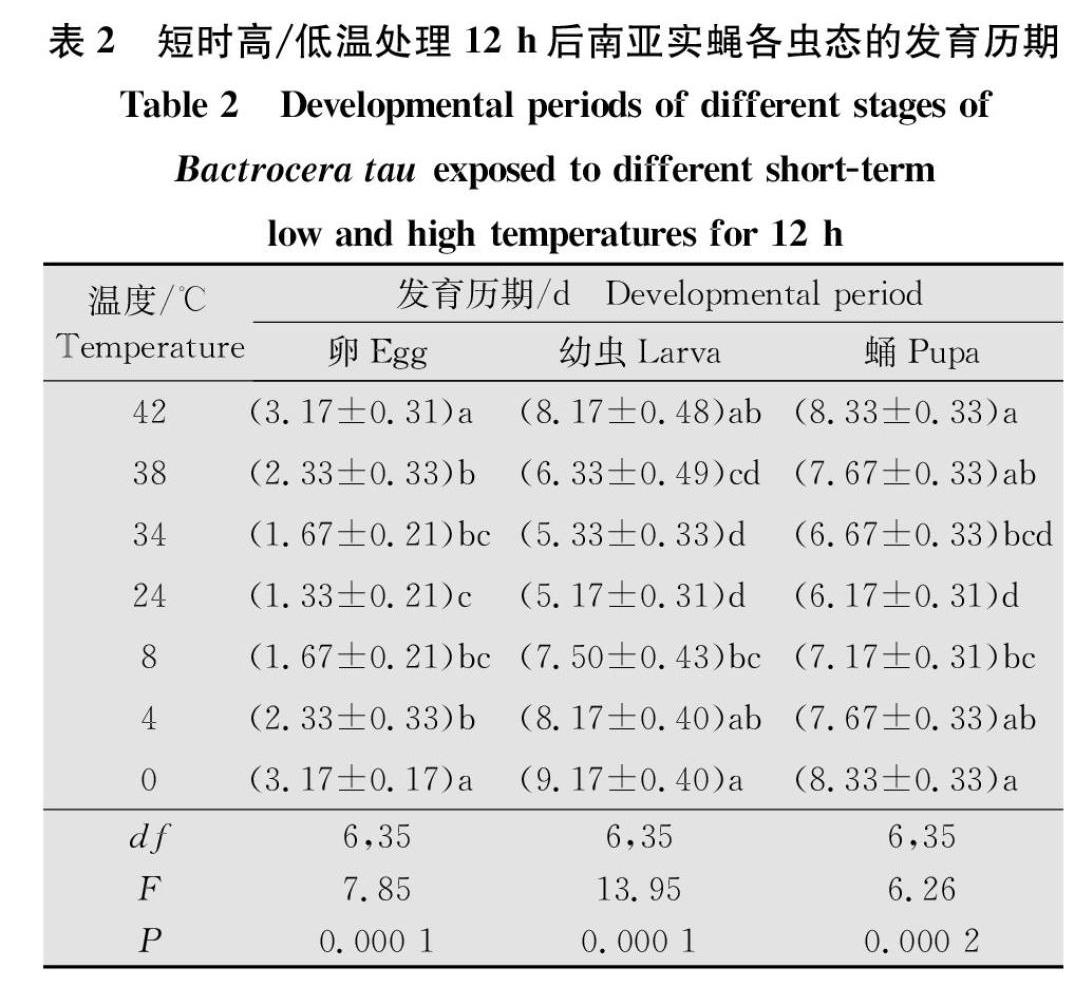
2.2 短时高/低温对南亚实蝇不同虫态发育历期的影响
不同的短时高/低温度处理南亚实蝇卵、幼虫和蛹,结果显示短时高/低温对不同虫态的发育历期有显著影响(卵:F6,35=7.85,P=0.000 1;幼虫:F6,35=13.95,P=0.000 1;蛹:F6,35=6.26,P=0.000 2)(表2)。随着处理温度的升高/降低,卵、幼虫和蛹的发育历期逐渐升高,其中,在对照温度下卵的发育历期最短,为1.33 d,處理温度为42℃和0℃时均为最长,达3.17 d;当处理温度上升至38℃或下降至4℃时,卵的发育历期都显著高于对照组(38℃:F1,10=6.429 0,P=0.029 6;4℃:F1,10=14.412 0,P=0.003 5);幼虫的发育历期在对照温度下最短,为5.17 d,42℃与0℃处理下的发育历期最长,分别达到8.17 d和9.17 d;当处理温度上升至42℃或下降至8℃时,该温度下幼虫的发育历期显著高于对照组(42℃:F1,10=27.931 0,P=0.000 4;8℃:F1,10=19.600 0,P=0.001 3);蛹的发育历期在对照温度下最短,为6.17 d,42℃与0℃处理下的发育历期最长,达8.33 d;当处理温度为38℃或8℃时,蛹的发育历期显著长于对照组(38℃:F1,10=10.946 0,P=0.007 9;8℃:F1,10=5.249 0,P=0.044 2)(表2)。综合不同短时高低温处理南亚实蝇各虫态的发育历期结果表明,在短时高温(38~42℃)和短时低温(0~8℃)处理时,随着温度的升高/降低,3种虫态的发育历期逐渐升高,且均在42℃和0℃时历期为最长。
2.3 短时高/低温对南亚实蝇成虫产卵前期的影响
分别在不同短时高/低温条件下进行处理,南亚实蝇成虫的产卵前期之间存在显著性差异(F5,12=12.320 0,P=0.000 2)(图1)。随着处理温度的升高/降低,产卵前期逐渐延长,其中,当处理温度上升至38℃和下降至4℃时的产卵前期显著高于对照组(38℃:F1,4=19.000 0,P=0.012 1;4℃:F1,4=16.071 0,P=0.016 0)(图1)。综合不同短时高/低温处理下南亚实蝇成虫的产卵前期结果表明,在短时高温处理后(34~38℃),成虫的产卵前期随着温度的升高逐渐延长,对照温度处理下产卵前期最短,同样的,在低温处理时(0~8℃)产卵前期随着温度的降低,逐渐延长,0℃处理下的产卵前期最长。
2.4 短时高/低温对南亚实蝇成虫单雌产卵量的影响
不同短时高/低温处理对南亚实蝇成虫的单雌产卵量有着显著的影响(F5,12=31.126 0,P=0.000 1)(图2)。其中,在短时高温处理后,随着处理温度的升高,单雌产卵量逐渐降低,当处理温度达到38℃时的单雌产卵量显著低于对照组(F1,4=73.144 0,P=0.001 0),且在38℃时的单雌产卵量最少;在短时低温处理后,随着处理温度的降低,单雌产卵量先增加后降低,8℃处理成虫后单雌产卵量显著高于对照组(F1,4=12.498 0,P=0.024 1),后随着处理溫度的降低,单雌产卵量逐渐减少,在0℃时最少。
2.5 短时高/低温对南亚实蝇成虫寿命的影响
成虫寿命也与温度密切相关,不同短时高/低温处理对南亚实蝇雌雄成虫的寿命有着显著的影响。分别在短时高/低温条件下处理,不同温度下同一性别成虫的寿命之间存在显著性差异(雌:F7,16=16.475 0,P=0.000 1;雄:F7,16=16.437 0,P=0.000 1)(图3)。其中,在短时高温处理后,雌雄成虫的寿命都随着处理温度的升高呈现先增加后减少的趋势,雌雄成虫的寿命在34℃时先增加,后随着处理温度的逐渐升高,寿命逐渐缩短,在42℃时最短,且在42℃时雌雄寿命均显著低于对照组的寿命(雌:F1,4=44.929 0,P=0.002 6;雄:F1,4=41.411 0,P=0.003 0);短时低温处理之后,雌雄成虫的寿命都随着处理温度的降低而缩短,当处理温度下降至4℃时,雌雄成虫的寿命显著低于对照组(雌:F1,4=8.278 0,P=0.045 1;雄:F1,4=8.123 0,P=0.046 4)。同一处理温度下雌成虫的寿命略长于雄成虫,但两者差异未达到显著水平(图3)。
2.6 短时高/低温对南亚实蝇后代雌性比的影响
分别在短时高/低温条件下处理成虫,南亚实蝇的后代雌雄性比之间存在显著性差异(F5,12=24.513 0,P=0.000 1)(图4)。随着处理温度的升高/降低,南亚实蝇后代雌性比逐渐增加,其中,当处理温度上升至34℃/下降至4℃时的后代雌性比显著高于对照组(34℃:F1,4=9.829 0,P=0.035 0;4℃:F1,4=8.887 0,P=0.040 7)。在高温处理后(34~38℃),后代性比随着温度的升高而增加,对照温度处理下最低,同样的,在低温处理时(0~8℃)后代性比随着温度的降低,逐渐升高。
3 结论与讨论
许多研究表明,温度对昆虫的生长发育起着至关重要的作用。每种昆虫都有其特定的温度适应范围,如果温度超过这种昆虫的耐受范围,它的生命活动就会受到抑制,严重的会导致昆虫死亡[31]。本研究结果表明,南亚实蝇各个虫态的存活率与温度密切相关,随着短时高温或低温处理中温度的升高或降低,各个虫态的存活率逐渐降低,当处理温度≥42℃时,除蛹以外的其余各虫态的存活率均低于50%,当处理温度≤-4℃时,除了雌雄成虫外,其余各虫态均死亡,且该极端温度下,雌成虫的存活率高于雄成虫。褚世海等的研究表明在35℃高温处理黑点切叶野螟Herpetogramma basalis幼虫16 h后,幼虫存活率小于25%,但随着处理温度的升高,幼虫的发育历期逐渐延长[32];瓜实蝇B.cucurbitae被短时高温处理后,各虫态存活率降低,发育历期延长,在24~42℃温度范围内,成虫产卵量逐随着短时处理温度的升高逐渐减少[28];随着处理温度从14℃降至2℃持续2 h,广聚萤叶甲Ophraella communa除蛹之外所有虫态的发育和存活都受到显著影响,随着处理温度的降低,成虫存活率显著降低[24];栎黄枯叶蛾Trabala vishnou gigantina越冬卵在被≤-30℃的低温处理后,无论处理时间的长短,均能导致越冬卵的存活率大幅度下降[33]。此外,在一定的温度范围内,意大利蝗Calliptamus italicus通过调节抗逆物质和保护酶的含量,能够在较高的温度下存活,但由于抗逆物质和保护酶的含量不同,雌性具有更快的反应速率和更强的耐高温性[34]。这与本文结果相一致。在初春和夏季,当南亚实蝇遭遇短时高低温后,对南亚实蝇各虫态的存活率均有着不利影响。
昆虫的不同发育阶段可以分为可活动阶段和非活动阶段,成虫和幼虫为可活动阶段,卵和蛹为非活动阶段,昆虫能够改变自己的行为和生理代谢,以避免环境温度升高时高温胁迫造成的损害,处于活动阶段的昆虫(成虫和幼虫)有可能逃离目前的有害环境,避免高温损害,然而,在类似情况下处于非活动阶段(卵和蛹)的昆虫只能通过提高其固有的耐热性来抵御不利影响[35]。因此,非活动阶段昆虫的耐热性通常高于活动阶段[3637]。不同的短时高/低温度胁迫下,南亚实蝇发育历期、产卵前期、单雌产卵量、寿命以及后代雌雄性比有着显著的差异,随着短时高/低温处理温度的升高和降低,发育历期、产卵前期和后代性比逐渐增加;随着高温处理温度的升高,雌雄成虫的寿命先增加后减少,雌成虫的单雌产卵量逐渐减少;随着低温处理温度的降低,成虫寿命逐渐缩短,但雌成虫的单雌产卵量呈现先增加后减少。该结果与其他研究结果相似,例如,Ma等的研究表明,当温度从29℃升高到34℃时,麦无网长管蚜Metopolophium dirhodum 的寿命和繁殖力均显著降低[38]。在43℃和45℃高温处理后,悬铃木方翅网蝽Corythucha ciliata的存活率、寿命和繁殖力均降低[39]。以往的研究认为,环境中过高或过低的温度都对昆虫的生殖产生不利影响[40]。但大多数结论均是建立在持续温度处理的研究结果上,本文则是研究短时温度变化对生长发育及繁殖的影响,与恒温处理的结果有差异,相对较高的短时低温(8℃)处理成虫12 h后,对单雌产卵量有显著促进作用,但并不是所有短时低温温度处理均对单雌产卵量有促进作用,过低或过高的短时温度对其单雌产卵量仍然会造成损伤。从能量角度分析,大多昆虫在遭遇低温后会将更多的能量用于生殖过程,从而降低存活率[41]。昆虫的产卵不仅仅与温度有关,还与湿度有着密切的关系,于莹等的研究表明在高温干旱条件下,褐飞虱Nilaparvata lugens雌成虫体重、单雌产卵量和卵孵化率均下降[42];杨帅等的研究表明,咖啡豆象Araecerus fasciculatus在相对湿度为90%时其成虫的寿命和单雌产卵量最高[43];草地螟Loxostege sticticalis在条件为21~22℃和RH60%~80%下,草地螟存活率高,生殖能力强[44]。以上研究表明相对高湿的环境对昆虫种群的发展也有影响,因此考虑温度条件的同时也应考虑湿度条件。本研究中,34℃的短时高温会使得雌雄成虫寿命略有延长,但与24℃下寿命无显著差异,随着处理温度继续的上升,则寿命缩短。其他研究也与该结果相似,郭苏帆的研究表明,韭菜迟眼蕈蚊Bradysia odoriphaga和异迟眼蕈蚊B.difformis成虫的寿命随着处理温度(24~34℃)的升高呈现出先增加后减少[45]。虽然本研究结果反映出短时高温和低温对南亚实蝇的繁殖和发育存在着不利的影响,但相对较低的高温处理对雌雄成虫的寿命有一定的延长作用,相对较高的低温处理对繁殖力有一定的促进作用,因此在南亚实蝇发生的地区,在初春出现短时低温后,应对南亚实蝇种群采取适当的控制措施。本研究系统地研究了短时高/低温胁迫对南亚实蝇生长发育和繁殖的影响,但短时低温和短时高温胁迫后昆虫体内的生理活性物质的变化有待于进一步研究。
参考文献
[1] WALKER F. List of the specimens of dipterous insects in collection of the British Museum, part4 [M]. London: British Museum, 1849: 6891172.
[2] SINGH S K, KUMAR D, RAMAMURTHY V V. Biology of Bactrocera (Zeugodacus) tau (Walker) (Diptera: Tephritidae) [J]. Entomological Research, 2010, 40(5): 259263.
[3] 汪兴鉴. 东亚地区双翅目实蝇科昆虫[J]. 动物分类学报, 1996, 21(S1): 1338.
[4] 李小珍. 南亚果实蝇种群特征及其对食料和热胁迫的生理调控机制[D]. 重庆: 西南大学, 2007.
[5] 毛紅彦, 丁华锋, 焦永吉, 等. 2013-2017年河南省南亚果实蝇种群动态监测[J]. 植物检疫, 2020, 34(1): 8284.
[6] 周锁奎, 李广学, 邱仲华, 等. 南亚寡鬃实蝇生物学特性观察及防治研究[J]. 植物保护, 1993, 19(5): 1112.
[7] HASYIM A, MURYATI, KOGEL W J. Population fluctuation of adult males of the fruit fly, Bactrocera tau Walker (Diptera: Tephritidae) in passion fruit orchards in relation to abiotic factors and sanitation [J]. Indonesian Journal of Agricultural Science, 2008, 9(1): 2933.
[8] 张艳. 海南省三种热带果园实蝇害虫种群动态及诱捕防控技术研究[D]. 海口: 海南大学, 2013.
[9] BAIMAI V, PHINCHONGSAKULDIT J, SUMRANDEE C, et al. Cytological evidence for a complex of species within the taxon Bactrocera tau (Diptera: Tephritidae) in Thailand [J]. Biological Journal of the Linnean Society, 2000, 69(3): 399409.
[10]张艳, 陈俊谕. 南亚果实蝇国内研究进展[J]. 热带农业科学, 2018, 38(11): 7077.
[11]林明光, 汪兴鉴, 曾玲, 等. 海南果蔬实蝇种类、地理分布及危害调查[J]. 植物检疫, 2013, 27(5): 8589.
[12]JAMNONGLUK W, BAIMAI V, KITTAYAPONG P. Molecular phylogeny of tephritid fruit flies in the Bactrocera tau complex using the mitochondrial COI sequences [J]. Genome, 2003, 46(1): 112118.
[13]KITTHAWEE S, DUJARDIN J P. The geometric approach to explore the Bactrocera tau complex (Diptera: Tephritidae) in Thailand [J]. Zoology, 2010, 113(4): 243249.
[14]韦淑丹. 广西南宁市瓜类实蝇的发生为害及瓜实蝇生物学和生态学特性[D]. 南宁: 广西大学, 2011.
[15]陈瑜, 马春森. 气候变暖对昆虫影响研究进展[J]. 生态学报, 2010, 30(8): 21592172.
[16]马春森, 马罡, 赵飞. 气候变暖对麦蚜的影响[J]. 应用昆虫学报, 2014, 51(6): 14351443.
[17]王艳敏, 仵均祥, 万方浩, 等. 昆虫对极端高低温胁迫的响应研究[J]. 环境昆虫学报, 2010, 32(2): 250255.
[18]MEEHL G A. More intense, more frequent, and longer lasting heat waves in the 21st century [J]. Science, 2004, 305(5686): 994997.
[19]HANSEN J, SATO M, RUEDY R. Perception of climate change [J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(37): 1472614727.
[20]SENEVIRATNE S I, DONAT M G, MUELLER B, et al. No pause in the increase of hot temperature extremes [J]. Nature Climate Change, 2014, 4(3): 161163.
[21]EASTERLING D R. Climate extremes: observations, modeling, and impacts [J]. Science, 2000, 289(5487): 20682074.
[22]刘超. 短时高温对亚洲玉米螟的影响[D]. 沈阳: 沈阳农业大学, 2016.
[23]徐天杨. 高温对玉米螟赤眼蜂生存和繁殖的影响[D]. 沈阳: 沈阳农业大学, 2017.
[24]罗敏, 郭建英, 周忠实, 等. 短时低温胁迫对广聚萤叶甲发育和生殖的影响[J]. 昆虫学报, 2011, 54(1): 7682.
[25]陈申芝, 张合伦, 高强, 等. 低温胁迫对东亚飞蝗存活及其生命特征的影响[J]. 亚热带农业研究, 2009, 5(4): 261263.
[26]刘丽红. 温度及补充营养对南亚实蝇Bactrocera tau(Walker)种群增长的影响[D]. 重庆: 西南农业大学, 2005.
[27]杨福田, 王庆云, 秦卓, 等. 4种杀虫剂对甜菜龟叶甲幼虫毒杀效果初探[J]. 中国植保导刊, 2020, 40(5): 8384.
[28]顾祥鹏, 黄禹禹, 张金永, 等. 短时高温胁迫对瓜实蝇生长发育及繁殖的影响[J]. 环境昆虫学报, 2020, 42(2): 391399.
[29]范秀娟, 陈丹, 孙志娟, 等. 短时高温对烟蚜生长发育、繁殖和取食行为的影响[J]. 昆虫学报, 2014, 57(10): 11881197.
[30]欧阳芳, 戈峰. 昆虫抗冻耐寒能力的测定与分析方法[J]. 应用昆虫学报, 2014, 51(6): 16461652.
[31]陈红松. 广聚萤叶甲对异常高温的生殖与生理代谢响应[D]. 长沙: 湖南农业大学, 2012.
[32]褚世海, 丛胜波, 李儒海, 等. 短时高温处理对黑点切叶野螟生长发育和繁殖的影响[J]. 福建农业学报, 2019, 34(11): 12881293.
[33]刘永华, 阎雄飞, 陆鹏飞, 等. 低温胁迫对栎黄枯叶蛾越冬卵发育和存活的影响[J]. 西北农林科技大学学报(自然科学版), 2018, 46(10): 126130.
[34]李爽, 王冬梅, 李娟, 等. 雌雄意大利蝗耐高温差异及其生理生化响应对策[J]. 应用昆虫学报, 2015, 52(4): 960967.
[35]HUEY R B, HERTZ P E, SINERVO B. Behavioral drive versus behavioural inertia in evolution: a null model approach [J]. The American Naturalist, 2003, 161(3): 357366.
[36]MARAIS E, CHOWN S L. Beneficial acclimation and the Bogert effect [J]. Ecology Letters, 2008, 11(10): 10271036.
[37]MITCHELL K A, SINCLAIR B J, TERBLANCHE J S. Ontogenetic variation in cold tolerance plasticity in Drosophila: is the Bogert effect bogus? [J]. Naturwissenschaften, 2013, 100(3): 281284.
[38]MA Chunsen, HAU B, POEHLING H. Effects of pattern and timing of high temperature exposure on reproduction of the rose grain aphid, Metopolophium dirhodum [J]. Entomologia Experimentalis et Applicata, 2003, 110(1): 6571.
[39]JU Ruiting, GAO Lei, ZHOU Xuhui, et al. Tolerance to high temperature extremes in an invasive lace bug, Corythucha ciliata (Hemiptera: Tingidae), in subtropical China [J/OL]. PLoS ONE, 2013, 8: e54372. DOI: 10.1371/journal.pone.0054372.
[40]劉安然. 温度对昆虫繁殖力的影响及其机制初探[J]. 江西农业, 2020(6): 105107.
[41]PAPAJ D R. Ovarian dynamics and host use [J]. Annual Review of Entomology, 2000, 45(1): 423448.
[42]于莹, 徐红星, 郑许松, 等. 在高温下干旱胁迫对褐飞虱生态适应性的影响[J]. 植物保护学报, 2013, 40(3): 193199.
[43]杨帅, 张涛, 高玉林, 等. 相对湿度对咖啡豆象生长发育、繁殖及种群增长的影响[J]. 应用昆虫学报, 2016, 53(1): 121127.
[44]罗礼智, 程云霞, 唐继洪, 等. 温湿度是影响草地螟发生为害规律的关键因子[J]. 植物保护, 2016, 42(4): 18.
[45]郭苏帆. 高温对韭菜迟眼蕈蚊和异迟眼蕈蚊生长发育及生理代谢的影响[D]. 兰州: 甘肃农业大学, 2018.
(责任编辑:田 喆)
