不同水平脂肪酶对罗非鱼生长性能、形体指数及血清生化、免疫指标的影响
2021-03-09作者龚俊勇吴伟坚钟国防
◆作者:龚俊勇吴伟坚钟国防
◆单位:1.深圳绿微康动物营养有限公司;2.上海海洋大学遗传与育种
脂肪在鱼类生长中具有十分重要的功能,可为其生长提供能量,能量为碳水化合物的2.25倍,是鱼类正常生长发育所必需的营养因子,还可提供必需脂肪酸;同时,可节约蛋白和降低氮排泄(Takeuchi T等,1978)。当饲料中的脂肪含量不足或者缺乏时,可能导致鱼类代谢紊乱,引发脂溶性维生素缺乏。当日粮脂肪含量过高,则会导致鱼体脂肪过量沉积,鱼体抗病力下降,产生营养性脂肪肝,影响鱼体对蛋白质的消化吸收与利用,从而影响鱼类的生长(王爱民等,2014)。然而,在实际养殖中由于过度添加低蛋白高能量日粮,导致鱼体中脂肪过度蓄积,严重制约鱼类养殖业的健康发展。
脂肪酶是脂肪消化利用最基本的酶,可将脂肪分解成甘油二酯、单甘脂、甘油和脂肪酸,提高饲料中油脂的消化吸收率,促进鱼类健康,减少脂肪肝的发生率。肉仔鸡日粮中添加脂肪酶,可提高其对脂肪的消化和黄羽肉鸡对脂质的代谢(叶慧等,2013)。在草鱼高脂肪水平日粮中额外添加适量脂肪酶可提高其生长性能及改善肠道健康(Liu S等,2016)。本试验在罗非鱼低脂日粮中添加不同水平脂肪酶,研究其对罗非鱼生长性能、形体指数及血清生化、免疫指标的影响,探讨其是否可达到高脂肪组水平性能,为脂肪酶在鱼类养殖中的应用及提高鱼体脂肪消化提供实践依据。
1 材料与方法
1.1 试验材料
选用上海海洋大学鱼类遗传与育种实验室培育的同批吉富罗非鱼为试验用鱼,规格整齐,健康无损害,平均体重为(14.32±0.15)g。
1.2 试验时间
2018年8月1日到10月7日,为期68d。
1.3 饲养试验
1.3.1 试验日粮
日粮组成及营养水平见表1。
1.3.2试验设计
试验选用480尾罗非鱼,随机分为6个组,每个组4个重复,每个重复20尾。正对照组饲喂基础日粮(脂肪含量高达8.6%),负对照组基础日粮降低能量50 kcal/kg,试验1-4组在负对照组基础上分别添加50、100、200、400 g/t脂肪酶。
1.3.3 饲养管理
养殖期间,每两周称重1次,调整投饲量,日投喂率为体重的3%~5%。每天投喂2次,投喂时间分别为7:00和17:00。每天早上清除箱内粪便,养殖期间水温控制为24.0±0.5℃,pH值为7.0±0.5,溶解氧不低于6.0 mg/L,氨氮控制在0.03 mg/L以下。
1.4 样品采集和分析
试验结束后禁食24 h,每组随机称鱼10尾,取血样、内脏及肝脏并称重。计算存活率(SR)、增重率(WGR)、饲料转化率(FCR)、特定生长率(SGR)、肥满度(CF)、肝体比(HIS)和脏体比(VSI)等。
同时,每缸中取5尾鱼从尾静脉抽取血液置于5 mL采血管中,待血液凝固后,4000 r/min(4℃)离心10 min,分离血清保存于-80℃备用。生理生化指标血清胆固醇、甘油三酯和尿素氮等采用日立7600全自动生化分析仪测定,测定试剂购自罗氏公司。谷草转氨酶(GPT)、谷丙转氨酶(GOT)、血清超氧化物歧化酶(T-SOD)、过氧化氢酶(CAT)、溶菌酶、补体C3和IgM,均采用南京建成生物公司的试剂盒按照说明书进行测定。
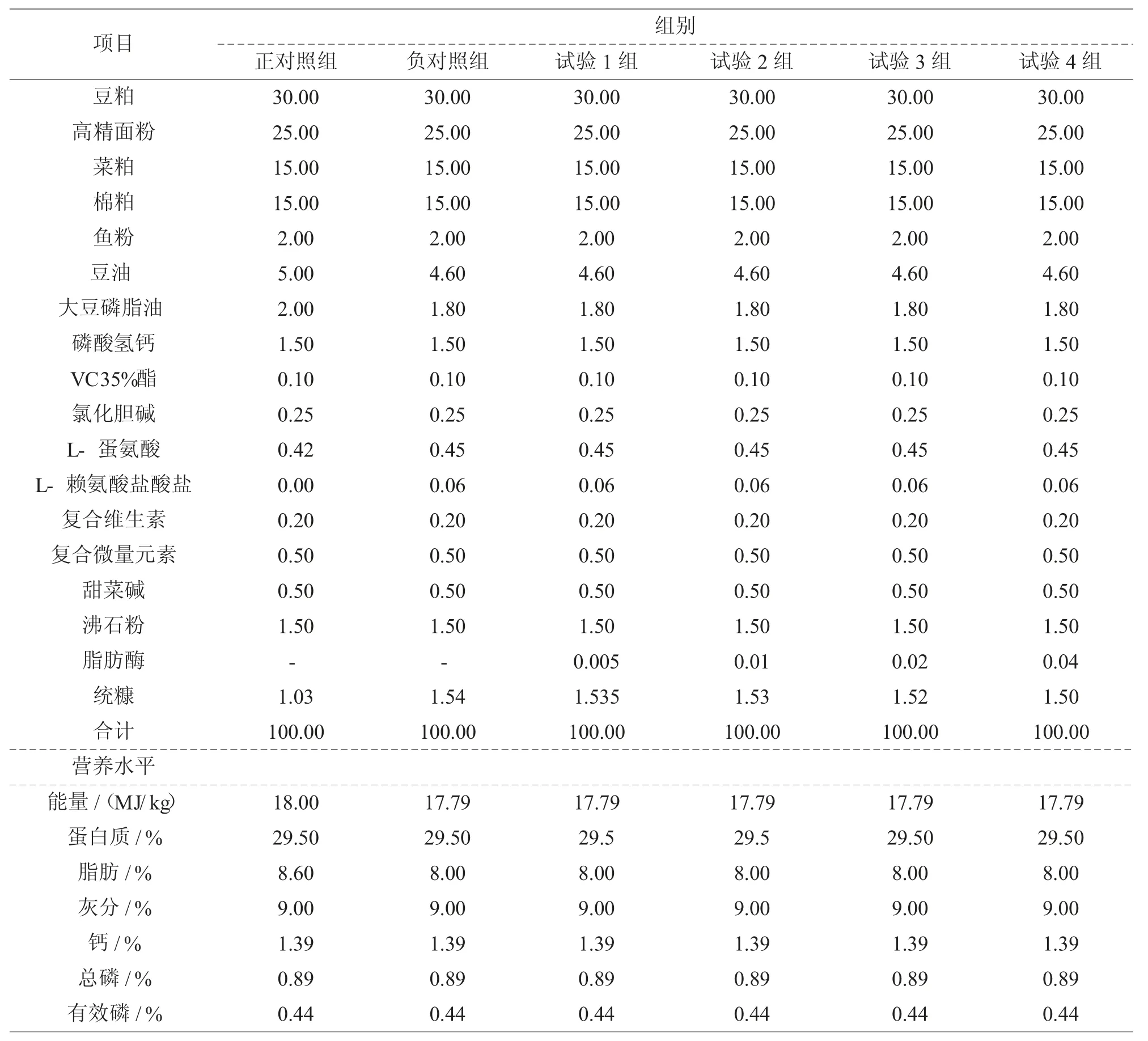
表1 罗非鱼饲料配方及营养水平

表2 添加不同水平脂肪酶对罗非鱼生长性能的影响
存活率(%)=100×末尾数/初始尾数
增重率(%)=100×(末均重-初均重)/初均重
饲料转化率(%)=消耗饲料量/增重
特定生长率(%/d)=100×[ln(末均重)-ln(初均重)]/饲养天数
肥满度(g/cm3)=100×(末均重-初均重)/体长3
1.5 数据处理
用SPSS 17.0处理数据,用单因子方差分析(one-way ANOVA,LSD)和Duncan氏多重比较检验统计结果。所有结果使用“平均值±标准差”(mean±SD)表示。以P<0.05为显著水平。
2 试验结果
2.1 添加脂肪酶对罗非鱼生长性能的影响
由表2可知,与负对照组比较,各试验组增重率分别提高6.97%、15.70%、11.41%、10.63%,且添加量为100、200及400 g/t试验组增重率显著高于负对照组(P<0.05);各试验组与正对照组增重率没有显著性差异(P>0.05)。与负对照组比较,各试验组FCR均有一定程度降低,但差异不显著(P>0.05)。脂肪酶的不同添加量对特定生长率及存活率无显著影响(P>0.05)。
2.2 添加脂肪酶对罗非鱼形体指数的影响
由表3可知,与负对照组比较,试验2、3组肥满度差异显著(P<0.05),组间无显著差异(P>0.05)。与负对照组比较,试验3组与负对照组差异显著(P<0.05),组间无显著差异(P>0.05);与负对照组比较,各试验组脏体比分别降低2.85%、9.54%、10.25%、8.29%,其中试验2、3、4组与负对照组差异显著(P<0.05);与正对照组比较,各试验组脏体比有降低的趋势,均差异不显著(P>0.05)。
2.3 添加脂肪酶对罗非鱼血清生化指标的影响
从表4可知:与负对照组比较,各试验组甘油三酯降低明显,均差异显著(P<0.05),组间无显著差异(P>0.05)。与负对照组比较,添加脂肪酶可以降低罗非鱼血清低密度脂蛋白含量,其中添加量为100、200 g/t试验组达到差异显著水平(P<0.05)。添加量为50、100、200 g/t试验组显著高于负对照组(P<0.05),且组间无显著差异(P>0.05)。
从表5可知:添加量为100、200、400 g/t试验组GPT显著低于负对照组(P<0.05),且组间无显著差异。与正对照组比较,各试验组GPT有降低趋势,但差异不显著(P>0.05)。与负对照组比较,100、200 g/t试验组GOT差
异显著(P<0.05);与正对照组比较,00、400 g/t试验组差异显著(P<0.05)。尿素氮、血糖含量各组间无显著影响(P>0.05)。
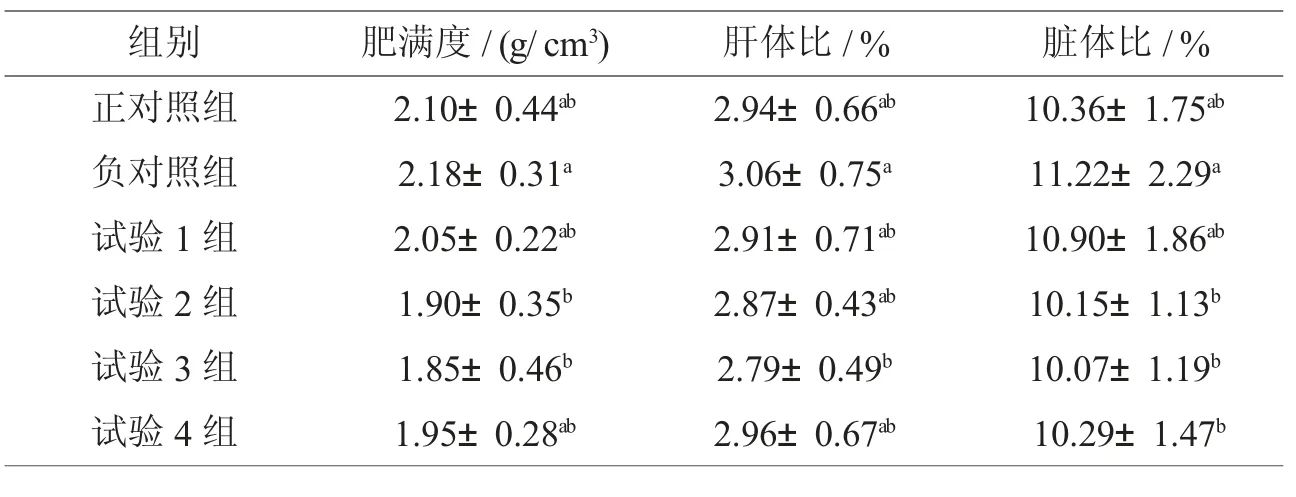
表3 添加不同水平脂肪酶对罗非鱼形体指数的影响
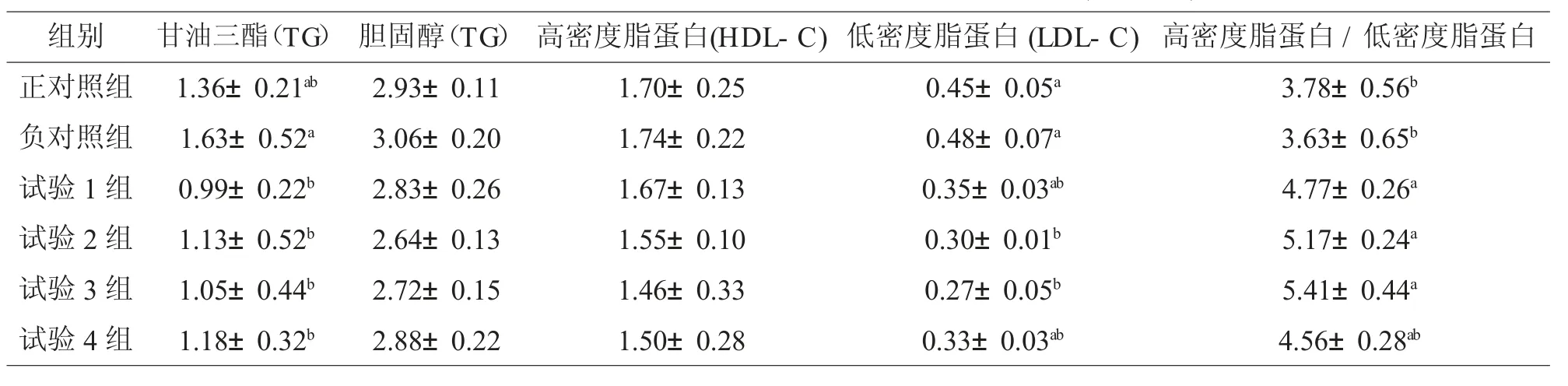
表4 添加不同水平脂肪酶对罗非鱼血清生化指标的影响(m mol/L)

表5 添加不同水平脂肪酶对罗非鱼血清生化指标的影响
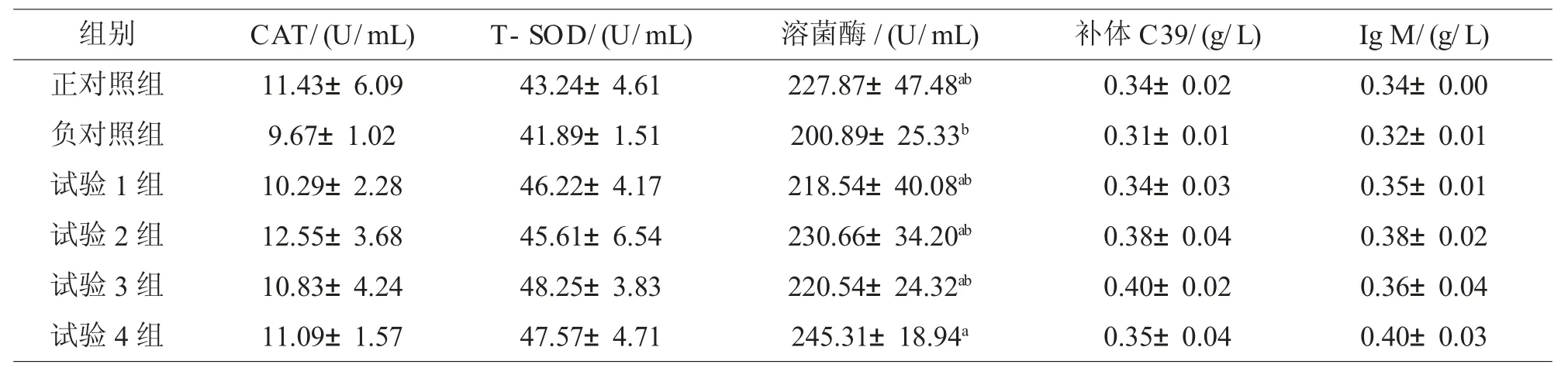
表6 添加不同水平脂肪酶对血清抗氧化免疫指标的影响
2.4 添加脂肪酶对罗非鱼抗氧化免疫指标的影响
从表6可知:与负对照组比较,添加不同水平脂肪酶后罗非鱼血清抗氧化指标CAT、T-SOD和溶菌酶酶活及免疫指标补体C3及Ig M含量均有不同程度的提高,均差异不显著(P>0.05),但400 g/t试验组溶菌酶提高22.11%,与负对照组差异显著(P<0.05)。
3 讨论
3.1 脂肪酶对罗非鱼生长性能的影响
脂肪酶在畜禽方面的研究应用表明,肉鸡日粮中添加脂肪酶可显著提高日增重,料肉比降低(叶慧等,2013)。水产方面,Troels等(2001)研究报道将脂肪酶添加于饲料中应用于虹鳟和金头笛鲷,结果表明,添加脂肪酶均可提高鱼类对脂肪的利用率。Dierick等(2002)研究指出,畜禽和水产中添加脂肪酶一定程度上可以提高生长。猪日粮中添加0.1%微生物脂肪酶,日增重率显著提高14.3%(P<0.01),饲料转化率提高5.4%。在对鳗鱼的研究中,添加0.2%脂肪酶显著提高鳗鱼日增重28.53%~71.38%、饲料转化率提高16.90%~26.99%,饵料系数降低14.69%~20.98%。王国霞等(2017)研究结果显示,在低蛋白高脂肪日粮中添加脂肪酶,花鲈生长性能有所降低。Liu等(2016)在低蛋白高脂肪中适量的脂肪酶(1193 U/kg,2560 U/kg),草鱼的生长性能和肠道免疫力得到提高。谷金皇等(2010)研究表明,添加100、300、500 mg/kg脂肪酶组分别比对照组增重率提高3.86%(P>0.05)、9.29%(P<0.05)、6.75%(P>0.05),饲料系数分别降低3.17%(P>0.05)、6.35%(P<0.05)、5.29%(P>0.05)。Silva等(2007)在大盖巨脂鲤日粮中添加0.2%脂肪酶,饲养期38d,较对照组增重率提高55.1%(P<0.05)。Nunes等(2006)在巨骨舌鱼(Arapaimagigas)日粮中添加0、0.05%、0.1%、0.2%脂肪酶,增重率提高29.3%、14.88%、65.9%,蛋白质效率也有不同程度提高。本试验结果显示:在罗非鱼基础日粮中添加不同水平脂肪酶,均可提高增重率,当添加水平为100、200、400 g/t时,增重率显著高于对照组,且FCR均有不同程度降低,差异不显著(P>0.05)。与之前的研究报道的结果相一致,但是也有相关研究指出,添加脂肪酶并不一定可以改善生长性能。X.Meng等(2004)在小麦基础日粮中添加脂肪源、糖酶、脂肪酶,结果发现添加脂肪酶对肉鸡的生长和养分的利用率均无显著提高。Martin等(1998)在含米糠的家禽饲料中混合添加溶血卵磷脂和脂肪酶,生长性能并未得到改善。
3.2 脂肪酶对罗非鱼形体指数的影响
肥满度和形体指数常作为衡量鱼类经济价值的重要指标之一。甘辉等(2009)研究指出,奥尼罗非鱼幼鱼的肥满度随饲料脂肪水平升高呈先上升后下降的变化趋势,且在饲料脂肪水平达到4%时最高,吉富罗非鱼的肥满度在饲料脂肪水平为5.69%时达最高,但当饲料脂肪水平为5.69%~9.64%时无明显差异。而脏体比常用来评价动物摄入能量的分配状况,肝体比用来衡量试验过程中肝脏功能的好坏。添加脂肪酶可促进鱼类对脂肪的利用,减少脂肪过度沉积,提高肥满度,降低脏体比和肝体比。朱龙等(2017)研究指出,添加胆汁酸作为乳化剂可显著提高肥满度(P<0.05),可显著降低脏体比、肝体比和脂肪沉积率(P<0.05)。这说明胆汁酸和脂肪酶均可作为乳化剂,可改善加州鲈的营养状况,减少内脏能量沉积,增加加州鲈的可使用的比例,提高其营养价值。原因可能是:胆汁酸作为乳化剂可改善动物肠道和肝胰脏健康(梁铁军,2007),消化吸收功能强;其次,胆汁酸可促进多余脂肪的分解,提高饲料脂肪利用率,减少脂肪在腹腔、肝脏组织中的不断累积,从而达到改善酮体品质的目的。但是也有些研究得到不同结果,Troels(2001)等研究在虹鳟饲料添加0.2%和0.02%微生物脂肪酶,仅鱼体成分中单不饱和脂肪酸的含量得到提高,而生长性能、内脏指数、肝胰脏指数和其他营养成分均无显著影响。
3.3 脂肪酶对罗非鱼血清生化的影响
血液中的TG、TC、LDL-C和HDL-C等水平的变化,可以反映出机体的脂类代谢状况和健康水平(邹思明,2005)。血清中TG和TC水平是反映肝脏脂肪代谢的重要指标(Krajnovic等,1992),血清中LDL-C水平升高,HDL-C水平下降,有利于清除组织和血清中胆固醇(王清滨,2014)。本试验中添加脂肪酶后血 清 中 TG、LDL-C及HDL-C/LDL-C水平降低,表明脂肪酶在一定程度上可降低罗非鱼油脂水平,这与饲料胆汁酸降低大菱鲆血清中TG、TC、LDL-C和HDL-C的水平(孙建珍,2014),以及草鱼血清中TC和HDL-C的水平的结果类似(陈昊杰,2016)。这表明,添加脂肪酶可加强罗非鱼的脂肪代谢,加强脂肪动员,促进机体对脂肪的利用,抑制脂肪过度沉积,促进机体对脂肪的利用,从而降低了血液中TG、LDL-C含量。叶慧等(2013)等研究指出,添加脂肪酶可以降低后期黄羽肉公鸡血清中总胆固醇、低密度脂蛋白胆固醇和高密度脂蛋白胆固醇的含量。谷金皇等(2010)研究指出,添加不同剂量脂肪酶可降低血清TG、TC、LDL-C含量,其中添加100、300、500 mg/kg组差异显著(P<0.05),300 mg/kg组的血清TG、LDL-C含量最低(P<0.05)。苏继影(2006)研究发现,添加脂肪酶可提高肝脏丙酮含量,促进肝脏进行脂肪代谢。
血清转氨酶是反映肝功能的主要敏感指标之一,血清GOT升高,说明肝细胞受到损伤的可能性增加,血清GPT升高意味着肝脏发生炎症的可能性增加(刘含亮等,2012)。谷金皇等(2010)研究结果显示,与对照组比较,添加500 mg/kg脂肪酶GOT和GPT显著降低(P<0.05),可能原因:添加脂肪酶可能主要通过促进肝脏脂类代谢实现,可一定程度上提高鱼体的非特异性免疫功能。王国霞等(2017)研究结果表明,在花鲈低脂饲料中添加脂肪酶后,血清GOT活性显著降低,血清抗氧化能力升高,分析可能的原因是:脂肪消化产生脂肪酸,适宜脂肪酸利于鱼的生长和机体健康,但是一旦产生过量脂肪酸,会引起鱼体的炎症反应,导致抗氧化指标升高。
3.4 脂肪酶对罗非鱼免疫指标的影响
SOD可将超氧阴离子(O2-)歧化为过氧化氢(H2O2)和氧气(O2),从而避免自由基对机体造成的氧化损伤(Fang Y Z等,2002)。溶菌酶是吞噬细胞杀菌的物质基础,主要针对革兰氏阳性菌发挥作用,溶菌酶活力越大,其溶解细胞的能力越强。IgM是动物机体发育过程中最早合成和分泌的抗体。补体系统中的C3是抗体的补充成分,参与机体的非特异性免疫反应。本试验结果显示:罗非鱼低脂日粮中增加不同水平脂肪酶,均可一定程度提高CAT、SOD、溶菌酶、补体及IgM水平,与高脂日粮水平差异不显著(P>0.05)。这说明草鱼中添加脂肪酶可一定程度提高免疫特性,可促进肝脏脂类代谢,在一定程度上提高鱼体的免疫功能,减少对肝脏的损坏。这与部分学者研究一致。谷金皇等(2010)试验结果显示,瓦氏黄颡鱼中添加脂肪酶后,溶菌酶、SOD活性均显著增强(P<0.05)。赵盼月(2019)研究指出,在欧洲鳗鲡幼鱼饲料中添加胆汁酸作为乳化剂可提高血清中IgM和C3的水平,说明胆汁酸可提高机体免疫功能,分析可能的原因是:胆汁酸和脂肪酶同作为分解脂肪类的物质,可提高机体免疫功能,及肝脏抗氧化能力,同时降低机体内对细胞内毒素的吸收。
4 结论
罗非鱼低脂肪日粮中添加不同水平脂肪酶,可改善生长性能及降低肝体指数。其中,脂肪酶添加量为100 g/t时,增重率最高,饲料系数最低;添加量为200 g/t时,对罗非鱼脏体比和肝体比的改善作用最明显。本试验条件下,在15g左右幼鱼低脂日粮中添加脂肪酶100~200 g/t对罗非鱼生长性能、肝体指数及血清生化、免疫指标的改善效果最佳。
