黄瓜CDPK基因家族的鉴定与进化特征分析
2021-03-05王谢琴姚慧鹏
靳 燕,王谢琴,卿 华,马 政,姚慧鹏
(四川农业大学生命科学学院,四川雅安 625014)
黄瓜(Cucumis sativus L.)葫芦科一年生蔓生或攀缘草本植物,是我国的主要蔬菜作物之一,具有重要的医药、营养和食疗价值。随着人民生活水平的不断提高,对食品质量的追求逐步升高,因此,研究黄瓜抗逆育种对培育品质优良的黄瓜品种,进而推动黄瓜农业种植生产具有现实意义。
钙依赖性蛋白激酶(Calcium-dependent protein kinase,CDPKs)是一种直接利用ATP激活的蛋白质底物以及Ca2+作为信号的酶,主要存在于植物和原生动物中[1]。典型的CDPK分子由1条多肽链组成,从N端到C端存在4个功能区(结构域),从N端到C端依次为可变结构域(variable N-terminal domain,VNTD)、Ser/Thr蛋白激酶域(Ser/Thr protein kinase domain,PKD)、自抑制连接域(junction domain,JD)和C端具有EF手型结构的类钙调素调控结构域(CaM-like regulatory domain,CaMLD)[2]。CDPK 在植物体内广泛分布,并在植物碳/氮代谢、细胞骨架调节、气孔运动调节和生长发育调节等均发挥一定作用[3],另外,其在植物对抗生物胁迫与非生物胁迫中也起着重要作用[4-5]。CDPK基因作为古老的基因家族,在原生生物如刚地弓形虫[6](Toxoplasma gondii),早期低等植物如盐藻(Dunaliella salina)[7]中都有发现,因此CDPK基因是作为研究植物基因进化的理想对象。
本文用生物信息学方法,在黄瓜的全基因组范围内鉴定出17个CsCPKs基因并分析其结构、染色体分布、进化关系及功能域。以期为深入探索黄瓜CDPK蛋白生物学功能提供理论依据。
1 材料和方法
1.1 试验材料
从在线网站 Ensembl(http://plants.ensembl.org/index.html)中下载黄瓜基因组数据以及从Pfam数据库中下载 CDPK特征文件 PF00069(kinase)、PF00036(CaM-LD)和 PF13499(EF-hand),后 在Linux系统中利用进行hmmer搜索第一次鉴定基因家族(设定阈值为E<1e-5)。在此基础上选择匹配更好的候选序列,进行特殊隐马尔可夫模型的构建。然后进行第二次hmmer搜索鉴定(设定阈值为E<1e-5)。之后筛选数据借助在线网站SMARTSMART(http://smart.embl.de/)NCBI CDD(https://www.ncbi.nlm.nih.gov/cdd/)Pfam(http://pfam.xfam.org/)确认候选基因结构完整性和结构域保守性,剔除冗余序列得到17个CsCPKs序列。
1.2 试验方法
对鉴定出的所有基因序列进行结构分析,在GSDS(http://gsds.cbi.pku.edu.cn/)获得黄瓜 CDPK的内含子和外显子数量及位置、在软件Mapchart上绘制基因在染色体上的分布情况。通过软件CSSPalm 3.0预测黄瓜CDPK蛋白氮端结构域是否能进行十六烷酰化。在线软件MEME(http://meme-suite.org/)探测CDPK中保守的motifs,同时在InterProScan(http://www.ebi.ac.uk/interpro/)软件搜索不同的 motif所对应的不同功能[8]。利用下载好的无油樟(Amborella trichopoda)、拟南芥(Arabidopsis thaliana)和葡萄(Vitis vinifera)等植物和黄瓜的CDPK编码蛋白序列进行多序列比对后,利用MEGA7.0,采用邻接法(neighbor joining,NJ),校验参数 Bootstrap 重复1 000次,构建无根系统进化树,进而推测CDPK基因的进化过程。
2 结果与分析
2.1 黄瓜CDPK基因家族的鉴定及其基本特征研究
通过两次进行hmmer搜索鉴定,黄瓜基因组包含了17条CDPK基因,这17条CDPK基因都包含STKD结构域和CaM-LD结构域以及EF手结构(图1)。黄瓜CDPK基因特征表现和在7条染色体上的位置见图2、3。并根据黄瓜CDPK基因在染色体上的排列顺序,将17条黄瓜CDPK基因命名为CsCPK1-17。

图1 黄瓜CDPK基因结构域Figure 1 Cucumber CDPK gene domain
2.2 黄瓜CsCPKs的进化分析
将黄瓜CDPK与在NCBI下载的无油樟Atr-CPKs、拟南芥AtCPK和葡萄VvCPKs基因序列构建最大似然进化树,可以看到黄瓜CDPK基因家族的系统进化关系。这17条CsCPKs根据拟南芥CDPKs的分类可以分为 A、B、C和 D 4个亚家族,A1、A2、A3、B1、B2、B3、B4、C1、C2、C3、C4 和 D 12 个超亚家族,在4个超亚家族中,D亚家族所含有的黄瓜CDPK最少。同样的,也可以将葡萄VvCPKs分为4个亚家族,12个超亚家族(图2)。同源的CsCPK位于染色体 1、3、4、6、7 上,而染色体 2 上无同源CsCPKs,这可能是多倍体事件发生之后,来源同一染色体的序列发生了丢失(图3),导致出现了同源CsCPKs。
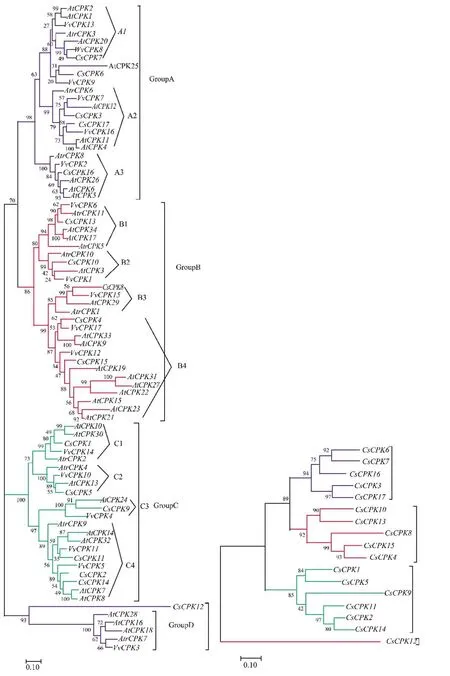
图2 拟南芥、无油樟、葡萄和黄瓜CDPK基因家族拓扑树Figure 2 Topological tree of CDPK gene family in Arabidopsis,oil-free camphor,grape and cucumber

图3 CDPK基因在黄瓜染色体上的分布Figure 3 Distribution of CDPK gene on cucumber chromosome
2.3 CsCPK中的结构差异
无论是在水稻还是在葡萄对CDPK的研究中,CDPKs的内含子对进化都十分重要。在水稻和葡萄中,D家族的内含子数量是最多的,这可能在进化过程中发生了内含子丢失事件,导致A、B和C家族的内含子数量减少。而在对CsCPKs的转录本结构分析来看,位于D家族的CsCPK的内含子数目与其他亚家族的内含子数目相差不大,都在6~8之间,证明黄瓜CDPK在进化过程中可能没有发生内含子丢失事件或者在此之前就早已发生了内含子的丢失(图 4,表 2)。

图4 17条CDPK基因内含子和外显子的基因结构Figure 4 Gene structure of 17 CDPK gene introns and exons

表1 CsCPKs的基因特征Table 1 Gene features of CsCPKs
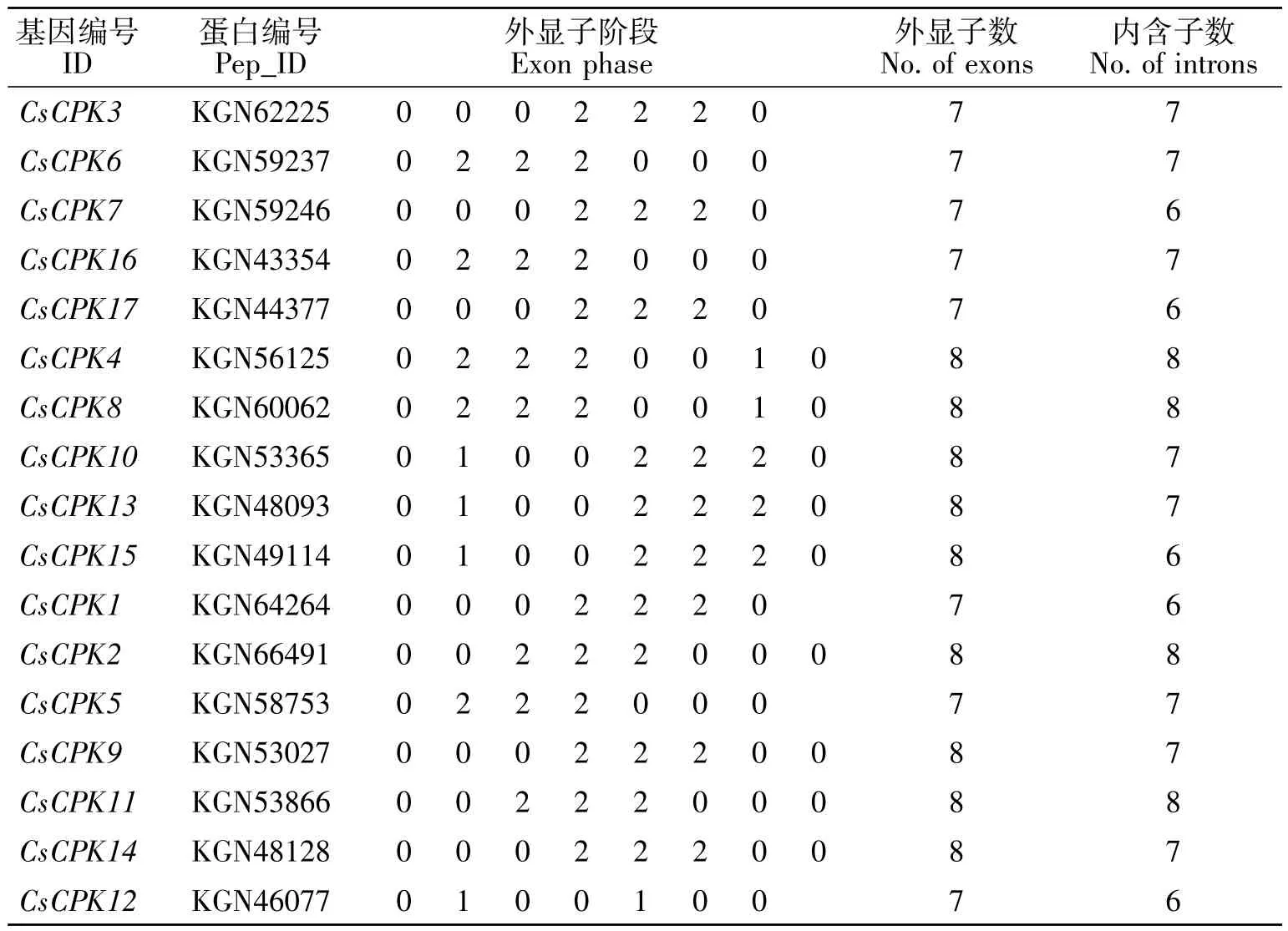
表2 17条CDPK基因内含子和外显子的数量Table 2 Number of introns and exons of 17 CDPK genes
根据在Css-palm3.0得到的结果表3来看,能进行棕榈酰化的CsCPKs除CsCPK3之外其余的CsCPKs都具有棕榈酰化位点,证明这些CsCPKs基因都与膜功能相关。李静等[9]研究中发现,在梨树和拟南芥CDPK的进化树中,PbCDPK5和PbCDPK7与AtCPK10和AtCPK30的亲缘关系最为接近,发现PbCDPK5和PbCDPK7与AtCPK10和AtCPK30一样都与逆境胁迫下的信号传导有关,证明了通过构建进化树进行对比分析可以预测一定的功能。而根据表2发现CsCPK9和VvCPK4最为接近,推测CsCPK9可能与VvCPK4一样在生殖和发育方面起作用。

表3 N端能进行十六烷酰化的CDPK基因Table 3 CDPK gene with N-terminal cetyl acylation
通过在MEME上对黄瓜CDPK蛋白质进行结构域扫描,搜索到了16个保守的motif。根据图5结果来看,所有的CsCPKs都具有motif1~10、motif12和motif14,这些motif构成了CDPK典型的结构域,而只有位于C亚家族的CsCPKs具有motif16(包括B家族的CsCPK15)和motif11,位于D亚家族的CsCPK12无 N 端的 motif3、motif13、motif14和 motif15,这可能是CsCPK12在进化过程中,N端发生了退化。根据在InterProScan搜索不同的motif所具有的不同功能,motif11和motif13与调节渗透性和盐度耐受性有关,motif15与植物生长发育有关,motif16与膜功能相关。经InterProScan对CsCPKs结构域搜索分析,所有的CsCPKs都具有ATP结合,Ca2+结合,蛋白激酶活性的分子功能,参与蛋白质磷酸化的生物过程,特有的motif决定了某些CsCPKs特有的生物学功能[8.10]。

图5 17条CsCPKs不同的motif组成Figure 5 17 CsCPKs composed of different motifs
3 讨论和结论
研究CDPKs的进化对了解植物抗逆作用机制有着重大的影响,此前F.Marianna等[11]分析到葡萄CDPKs基因分为4个亚家族,12个超亚家族,在进化的过程中发生了13次基因组加倍事件,3次大型全基因组复制事件。CDPK最早是存在于衣藻中,随着基因组的扩张,逐步出现在裸子植物,被子植物当中。不仅如此,叶水烽博士根据进化树分析,发现水稻中的CDPKs也分别处于4个亚家族当中,第四亚家族中的CDPKs的内含子个数明显比其他族的多,且衣藻的两个CDPK基因都位于这一组,可以得出第四亚家族是最早扩张出来的[12]。我们不仅可以通过与拟南芥葡萄构建进化树来分析黄瓜CDPK的进化情况,还可以通过内含子和外显子的分布情况来分析黄瓜CDPK基因进化的先后顺序。张进等[13]对杨树CDPK基因与拟南芥CDPK基因做同源性分析,也将杨树的CDPK基因分为4个家族,且第四家族的基因数最少。
研究CDPKs的结构和进化史对了解植物的功能有着重大的影响,Fasoli Marianna等[11]通过对17个葡萄CDPK基因进行motifs探测,发现催化区由motif6、motif3、motif1和motif4组成,连接区和结构区由motif2和motif5组成,可变区中只有A1家族含有motif9,其他的葡萄CDPK如VvCPK7等的可变区中未探测到motif,猜测VvCPK7可能发生了变性或存在不保守序列。不同的motifs对应不同的功能,这暗示了葡萄CDPK功能的多样性。J.Sheen[14]发现,在干旱和盐诱导条件下,拟南芥AtCPK1和AtCPK2的表达量显著增加,拟南芥AtCPK10和AtCPK30参与了对ABA和非生物胁迫的信号转导。在梨树和拟南芥CDPK的进化树的研究中,李静等[9]发现PbCDPK5和 PbCDPK7与AtCPK10和AtCPK30的亲缘关系最为接近,就猜测PbCDPK5和PbCDPK7与逆境胁迫下的信号传导有关。经过Q-PCR试验结果发现,在干旱处理8 h后,PbCDPK5和PbCDPK7含量升高,证明了通过构建进化树进行对比分析可以预测一定的功能。
由于最早分化出来的绿藻中很少有CDPKs基因,因此黄瓜CDPKs的形成必然经历了多次基因组扩张事件[15]。在植物的CDPKs进化史中,发生了3次大型基因组复制事件[10,16-17],分别是距今4亿年至3亿年前的ζ全基因翻倍,距今3亿年至2亿年前的ε全基因翻倍以及距今大约1亿1 700万年前的γ全基因翻倍。D家族的起源最早,最早为单一基因家族,后经ζ全基因翻倍出现了B1、B2、C1和C2超亚家族,ζ基因组扩张事件发生在维管植物出现之后,种子植物出现之前。ε全基因翻倍事件的发生导致了B3、B4、C3和C4超亚家族的产生,且ε全基因翻倍事件发生在种子植物出现之后,被子植物出现之前。γ全基因翻倍事件的发生导致了VvCPK9、13,VvCPK12、17,VvCPK5、11 的出现,通过黄瓜CDPK基因家族与葡萄CDPK基因家族构建的拓扑树来看,CsCPK6出现在VvCPK9之后,CsCPK4和CsCPK15 出现在 VvCPK12、17 之后,CsCPK2、11、14出现在VvCPK5、11之后,证明γ全基因翻倍事件产生了 CsCPK2、4、6、11、14 和 15(图 2)。在 3 次基因组扩张事件发生之后,无其他基因组扩张事件的发生,证明CsCPKs至少出现在1亿年前,而异源多倍体化事件发生在6 500万年前左右[18],CsCPKs的进化史并不支持异源多倍体事件促进CsCPK进化的理论,而支持由基因组扩张事件引起进化的理论。如果CsCPKs发生了异源多倍体化,那么CsCPKs肯定经过彻底的纯化选择作用。
CDPKs基因广泛存在于各种植物中,通过对黄瓜CDPKs的研究,发现黄瓜具有17条CDPKs,且在4个亚家族中均有分布,在目前对已测序的葫芦科植物的研究中,西瓜和甜瓜的CDPKs分别有22条和18条[9,19],黄瓜的CDPKs数目与甜瓜的最为接近,全基因组复制事件导致CDPK基因家族在不同的物种中数目出现差异[20]。
我们看到黄瓜的CDPK基因组出现在VvCDPK之前或在VvCDPK基因之后分成新基因,我们可以推测黄瓜的出现应该早于葡萄。随着对黄瓜的研究,我们可能会挖掘更多的黄瓜同源CDPK基因,并可能逐渐了解到CsCPKs基因是如何在逆境条件下响应的以及在黄瓜生长发育过程中是扮演的什么样的角色。
