晋西黄土区不同水文年土壤水分特征及其主要影响因子分析
2021-03-04孙占薇梅雪梅刘京晶王福星张金阁
孙占薇,马 岚,梅雪梅,刘京晶,王福星,张金阁,燕 琳
(1. 北京林业大学 水土保持学院,北京 100083;2. 北京林业大学 山西吉县森林生态系统国家野外科学观测研究站,山西 吉县 042200;3. 华北水利水电大学 水资源学院,河南 郑州 450046)
土壤水分作为植被生长的重要限制因素[1],对生态系统和水文过程起着重要作用。黄土高原地区覆盖的广袤而深厚的土层,为降水转化为土壤水提供了有利的条件。退耕还林(草)措施实施后,黄土区植被覆盖得到了极大改善,覆盖度由31.6%增加到59.6%[2],但该措施同时也加快了土壤的干燥化速度,使得植被缺水问题日益严峻[3]。因此,土壤水分的变化规律和影响因素成为了近些年黄土高原地区的研究热点问题。土壤在接受降水补给的同时,也因植被生长而消耗水分。已有研究表明:黄土高原地区人工植被下的土壤存在不同程度的水分亏缺现象[4]。不同水文年降水量及季节雨水分配的差异,使得不同时段土壤活跃层存在差异[5]。土壤储水能力受植被类型、地形和土壤理化性质等多因素的制约,这些环境因子通过交互作用直接或间接发挥着作用[6]。目前,关于黄土区土壤水分时空动态变化的研究成果较为丰富[7−10],但是大多数研究集中在某年的特定时间段,缺乏代表性,且侧重变化规律或单一影响因素的研究。本研究针对晋西黄土区具有代表性的人工刺槐Robinia pseudoacacia林、天然三角槭Acer buergerianum林和野艾蒿Artemisia lavandulaefolia草地,探究严重干旱年、平水年和中等湿润年的土壤水分变化规律、水分盈亏状况及环境因素对不同土层储水量的影响,为探索有利于区域土壤水分储存的植被类型提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于山西省临汾市吉县蔡家川流域 (36°14′~36°18′N,110°40′~110°48′E),地貌属于黄土高原残塬沟壑,海拔904~1 592 m,平均海拔1 168 m。该地属暖温带大陆性季风气候,多年平均降水量为571.0 mm,4−10月降水量占全年的90.5%左右,年平均气温10.0 ℃,潜在蒸发量1 729.0 mm。土壤类型为褐土。研究区内分布着近190余种木本植被,常见天然植被如辽东栎Quercus liaotungensis、山杨Populus davidiana、白桦Betula platyphylla、侧柏Platycladus orientalis、白皮松Pinus bungeana、黄刺梅Rosa xanthina林等;人工植被主要是刺槐、油松Pinus tabuliformis、侧柏林等。研究区地理位置见图1。
图1 研究区地理位置示意图Figure 1 Geographical location of studied area
1.2 样地设置
本研究在蔡家川流域内的嵌套小流域进行,以3种典型植被(人工刺槐林、天然三角槭林和野艾蒿草地)为研究对象,乔木林地采用20 m×20 m的样方,草地采用1 m×1 m的样方。样地基本情况见表1。

表1 样地基本情况Table 1 Basic information of experimental sample plots
1.3 方法
在群落调查的基础上,随机选择3个样地样点,采用土钻法对0~400 cm土层以20 cm为间隔采集土壤样品。将获得的土壤样品一份装入铝盒带回实验室,烘箱设定恒温105 ℃,烘干24 h,放置室温后测定土壤含水量,另一份用于测定土壤其他理化性质。采用环刀法对0~100 cm土层以10 cm为间隔测定土壤容重。采用MalvernSizer 2000超声波粒度分析仪对土壤粒径进行分析,采用重铬酸钾氧化-外加热法测定土壤有机质质量分数,采取环刀法测定0~100 cm土层土壤孔隙度,以上指标均设置3个重复。研究期内降水气象资料由观测点附近的气象站获得。土壤水分监测时间为2016、2017和2019年3 a的生长季初期(5月)、中期(7月)和末期(10月)。
1.4 数据处理
标准化降水指数(ISP)为MCKEE等[11]提出的评价旱涝程度的指数,其优点在于化解了降水时空分布差异大,难以直接相互比较的问题。标准化降水指数可以基于特定时间尺度计算降水量之和,基于降水量的Γ分布,并通过正态标准化得到[12]。假设某一时段的降水量为x(mm),则其Γ分布的概率密度函数为:

式(1)中:Γ指的是Gamma函数。

式(2)中:α、β分别代表形状参数和尺度参数(α>0,β>0)。用最大似然法估算α和β的最佳值。即:

式(3)~(5)中:n为计算序列的长度,为多年平均降水量。A代表观测点的数量。
根据ISP数值,MCKEE等[11]将水文年划分为7个大类,分别是ISP≤−2.00为极端干旱年,−2.00<ISP<−1.50为严重干旱年,−1.50≤ISP<−1.00为中度干旱年,−1.00≤ISP<1.00为平水年,1.00≤ISP<1.50为中度湿润年,1.50≤ISP<2.00为非常湿润年,ISP≥2.00为极端湿润年。
土壤储水量(SWS)的计算公式为:

式 (6)中:θi为第i层体积含水量 (cm3·cm−3),hi为土层厚度 (mm)。
土壤含水量标准差(DS)能够反映不同土层土壤水分变化情况。为了研究不同水文年活动层变化特征,采用土壤含水量标准差,将土壤剖面水分分布划分为3层,分别为土壤水分活跃层(DS≥1.5),土壤水分次活跃层(1.0≤DS<1.5)和土壤水分相对稳定层(DS<1.0)[13]。
为分析环境因子对土壤储水量的影响程度,通过冗余分析(RDA)探究环境因子对不同土层(0~100、100~200、200~300和300~400 cm)土壤储水量的影响,环境因子包括植被类型、不同水文年旱涝程度、坡度、坡向、土壤有机碳质量分数、黏粒含量、粉粒含量、砂粒含量、容重和毛管孔隙度。采用前向选择剔除部分冗余变量,并通过蒙特卡洛检验分析筛选出的环境因子与土壤储水量是否存在显著相关性。
本研究中方差分析、LSD多重比较采用SPSS 18.0,冗余分析(RDA)采用CANOCO 4.5,其他基础数据处理和绘图采用Excel和Origin。
2 结果与讨论
2.1 土壤水分概况
根据ISP对研究期内各时间段的干旱程度进行划分显示:2016、2017及2019年ISP分别为−0.33(平水年)、1.41(中等湿润年)以及−1.57(严重干旱年)。人工刺槐林、天然三角槭林和野艾蒿草地平均土壤含水量分别为8.36%~9.63%、10.01%~13.19%、15.43%~19.17%(表2)。3种植被类型0~100 cm土层土壤含水量差异显著(P<0.05),100 cm以下土层含水量差异不显著(P>0.05)。野艾蒿草地土壤含水量较高,天然林次之,而人工刺槐林最低。人工刺槐林冠层较高,地面灌草植被覆盖程度大于天然三角槭林,相应耗水量较大,导致人工林表层土壤水分含量较低。0~100 cm人工刺槐林、天然三角槭林和野艾蒿草地土壤含水量标准差分别为2.12、1.43和1.68,人工刺槐林变异程度最高,野艾蒿草地次之,天然三角槭林变异程度最低,100~400 cm土壤变异程度减小。不同植被类型土壤表层水分波动较大,而深层变幅平缓。该结论与汪星等[14]和徐志尧等[15]在对黄土高原半干旱区土壤水分动态变化研究结论一致,土壤含水量随着深度变化发生浮动,后趋于恒定。土壤表层受地表风力、辐射和降水的影响,容易产生波动,而深层干扰较小[16]。

表2 不同植被类型平均土壤含水量多重比较Table 2 Multiple comparisons of average soil water contents of different vegetation types
2.2 土壤水分垂直分布的年际差异
土壤水分垂直分布特征如图2所示。可以看出:生长季内,人工刺槐林和天然三角槭林的土壤含水量随着土层深度的增加而减小,而野艾蒿草地则相反。土壤活跃层变化更直观地反映土壤垂直剖面的年际差异。不同水文年0~400 cm活动程度划分结果见表3。对于严重干旱年,人工刺槐林、天然三角槭林和野艾蒿草地活跃层下边界分别为60、80、40 cm。而在平水年人工刺槐林及野艾蒿草地水分活跃层深度均不足50 cm,但天然三角槭林可达120 cm。在中等湿润年,人工刺槐林、天然三角槭林和野艾蒿草地水分活跃层分别可达140、180和140 cm。在所研究的水文年,天然三角槭林的土壤水分活跃层始终较深,而土壤活跃层反映了土壤受降水和蒸发等因素的影响程度[17]。这表明天然三角槭林土壤受外界因素干扰较强。人工刺槐林、天然三角槭林和野艾蒿草地活跃层深度从大到小均表现为中等湿润年、严重干旱年、平水年。次活跃层非普遍存在,天然三角槭林和野艾蒿草地次活跃层均出现在水分较为匮乏的干旱年,而人工刺槐林的次活跃层出现在平水年和中等湿润年。

图2 土壤剖面含水量变化Figure 2 Change of soil water contents with soil depth

表3 样地土壤水分活动层划分Table 3 Division of soil water active layers in sample plots
黄土高原地区降水对土壤补给的深度主要集中在100~300 cm,土壤水分活跃层主要集中在100 cm左右。随植被恢复年限的增加,活跃层深度也有所增加[18]。本研究表明:土壤活跃层深度最深可达200 cm左右。野艾蒿草地活跃层深度较浅,人工刺槐林和天然三角槭林活跃层较深,这主要是由于林地如刺槐等具有发达的根系,由主根和多层侧根构成,侧根丰富而发达[19],且随着林龄增长,根系生物量逐年增加,根系具有感知水分梯度的能力,其吸水作用将土壤水分向下牵引,而草地的根系分布较浅,对深层土壤水分难以起到作用。土壤水分活跃层深度在干旱年和湿润年大于平水年。这主要是由于黄土丘陵区干旱年生长季降水量缺乏,旱季和雨季降水的分布存在明显差异,导致了土壤水分和根系分布的季节性差异[20]。同时湿润年土壤水分在生长季整体性增加,雨季大量降水补充了植物生长消耗的水分,水分下渗程度也较高,生长季前后水分差异较大,从而使得湿润年土壤活跃层最深[21]。
2.3 土壤水分盈亏状况的年际差异
生长季前后土壤储水量差值的大小变化反映了年内土壤水分补给或消耗特征。人工刺槐林、天然三角槭林和野艾蒿草地生长季表层土壤储水量变化范围较深层土壤大,深层储水量变化接近于0(图3)。严重干旱年,生长季土壤储水量降低,表明由于降水量有限,不足以补充蒸散,不同植被类型均出现土壤水分亏缺的现象。人工刺槐林100~300 cm土壤水分亏缺程度较浅层(0~100 cm)和深层(300~400 cm)更为严重,天然三角槭林在0~200 cm水分亏缺最为严重,而在深层部分土壤得到了补给。对于野艾蒿草地,0~100 cm土层亏缺程度最高,随深度增加,亏缺程度减弱。人工刺槐林和天然三角槭林的浅层土壤水分损失量大于野艾蒿草地。平水年生长季,土壤水分的输入和输出达到平衡,各植被类型土壤储水量差异较小。中等湿润年生长季,各植被类型均表现出降水补给土壤水的现象。在0~100 cm土层中,野艾蒿草地的土壤水分补给量最高;在100~200 cm的土层中,天然三角槭林的土壤水补给量最高;在深层土层(200~400 cm)中,各植被类型土壤水分补给量较小,接近于0。

图3 不同植被类型土壤水分盈亏状况Figure 3 Status of soil water in different vegetation types
黄土高原土壤的蓄水能力受到土壤性质的影响,其入渗深度一般较浅,降水无法到达土壤深层,使得土壤蓄水能力在浅层和深层存在较大的差异[22]。本研究中,人工刺槐林中等湿润年的土壤水分补给量较小,严重干旱年水分亏缺情况较严重,且水分亏缺土层与天然三角槭林和野艾蒿草地不同,说明了人工刺槐林地的耗水模式与天然三角槭林和野艾蒿草地的耗水模式存在差异。这主要是因为人工刺槐林扎根较深,对深层水分的吸收大于其他林地,其深根系的分布,更是改变了土壤理化性质。水分的过度消耗极有可能会导致土壤出现干层,从而阻碍降水的垂直入渗进而导致该地区的地下水无法得到补给[23]。降水特征、气象因子、冠层结构和这些因素的相互作用影响了林冠降水的再分配[24]。与人工刺槐林相比,天然三角槭林枝叶茂盛,林冠结构更为扁平致密,该结构不利于穿透雨的形成[25]。野艾蒿草地深层土壤较难得到补给的原因是根系主要分布在浅层[26],表层土壤吸收了大部分降水,从而使得深层土壤水分很难通过降水入渗得到补给。
2.4 环境因子对土壤水分的影响
土壤储水能力与植被类型、地形、土壤理化性质、降水等因素息息相关。由图4可以看出:0~100 cm土壤有机碳质量分数和砂粒含量较高,随土层深度增加降低,黏粒含量和粉粒含量变化趋势相反,以上性质在100 cm以下土层相对稳定。因此将土壤有机碳、黏粒、粉粒和砂粒分为0~100 cm和100~400 cm等2个部分与土壤储水量进行分析。冗余分析(RDA)结果表明:环境因子解释了前4轴土壤储水量变化的93.6%,其中环境因子解释了前2轴土壤储水量变化的89.4%,因此前2轴很好地说明了环境因子对土壤储水量的影响。单个环境因子与前2轴的土壤储水量变化的相关系数见表4,环境因子与第1轴的相关系数更大,坡度、植被类型、100~400 cm黏粒含量、容重以及毛管孔隙度与第1轴均表现为极显著相关(P<0.01)。
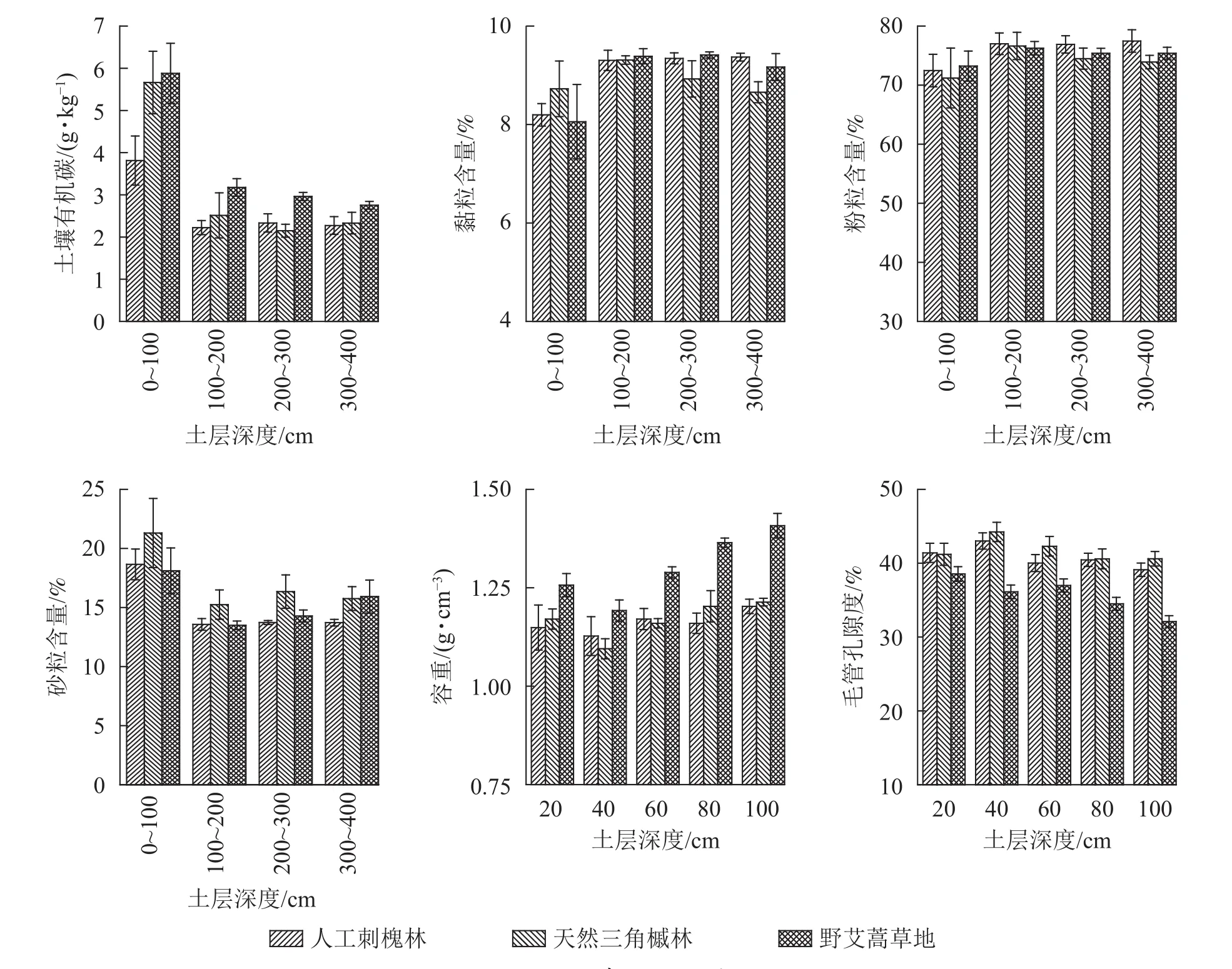
图4 土壤理化性质Figure 4 Physical and chemical properties of soil
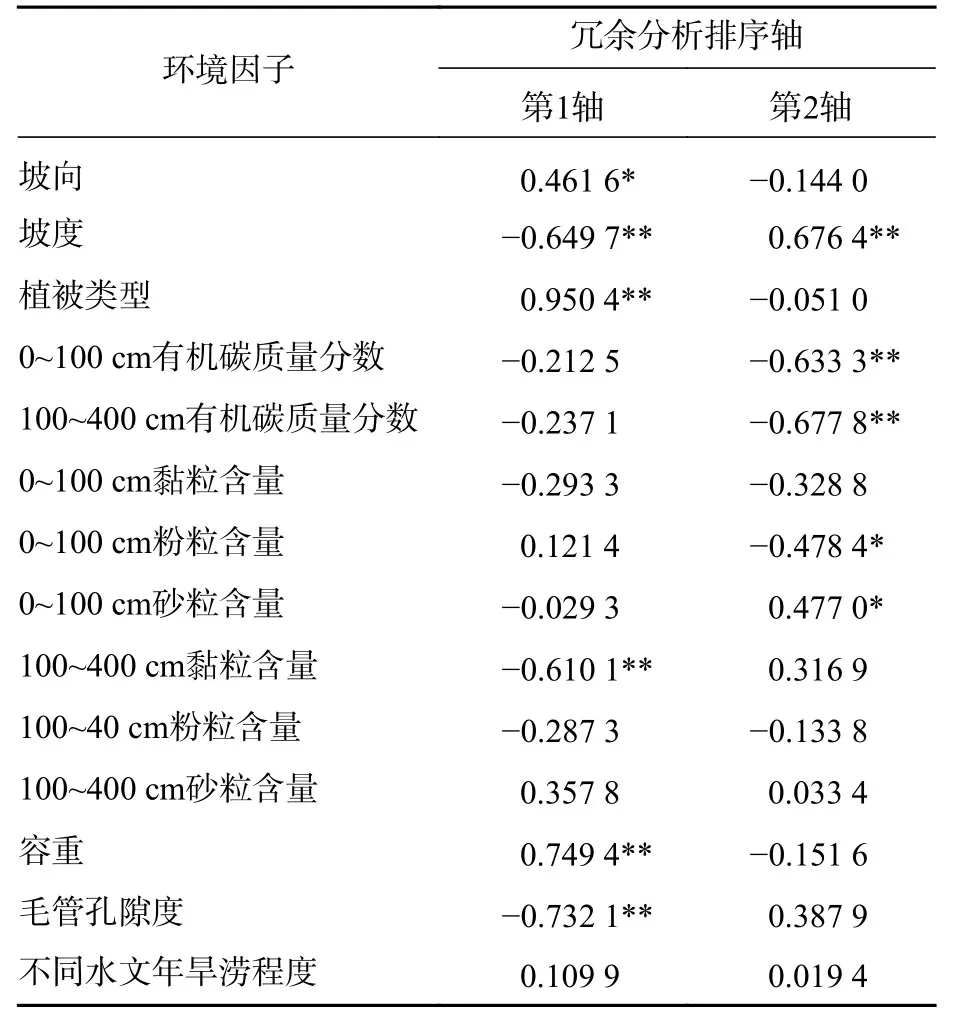
表4 环境因子与前 2 轴相关系数Table 4 Correlation coefficients between environmental factors and the first two axes
RDA排序图展示了不同土层深度土壤储水量与环境因子之间的关系(图5)。图5显示:植被类型、容重和坡向是影响0~100 cm土层土壤储水量的主要环境因子,容重和坡向是影响100~200 cm土层土壤储水量的主要环境因子,不同水文年旱涝程度和粉粒含量是影响200~300 cm土层土壤储水量的主要环境因子,300~400 cm土层土壤储水量主要是由土壤黏粒含量和毛管孔隙度控制。对影响土壤储水量的环境因子进行前向选择和蒙特卡洛检验结果显示(表5):主导土壤储水量的环境因子为植被类型、坡度、不同水文年旱涝程度以及土壤表层的黏粒含量(0~100 cm),其中植被类型、坡度与土壤储水量极显著相关(P<0.01),它们分别解释了土壤储水量66.4%和14.2%,而不同水文年旱涝程度和土壤表层粉粒含量与土壤储水量显著相关(P<0.05),它们分别解释了土壤储水量的3.3%和2.4%。

图5 土壤储水量与环境变量的冗余分析排序图Figure 5 RDA ranking chart of soil water storage and environmental variables

表5 环境因子的前向选择和蒙特卡洛检验Table 5 Forward selection and Monte Carlo test of environmental factors
不同植被类型的林冠层结构、凋落物以及根系等的差异影响了植被截留、蒸发和吸水,这些因素在不同时间尺度上影响着土壤水分的分布[27]。传统观点认为:旱季土壤理化性质主导了土壤水分的空间分布,雨季则主要受地形因素的影响[28]。土壤水分的主控因素在不同季节表现差异较大[29]。本研究区非生长季,坡向极大地影响着融雪程度[30],而在干旱生长季,研究区盛行西北风,坡向对浅层土壤储水量的影响极大[31]。黄土高原地区降水对土壤水分的补给程度可达1.6~2.3 m[32]。当出现低强度连续性降水时,深层土壤水可以得到一定程度的补给,可见在水分条件较好时段,降水可以补给2.0 m以下较深土层,因此不同水文年的旱涝程度(降水量分布情况)对200~400 cm土层储水量的影响作用不可轻视。土壤物理性质是土壤其他性质的结构性基础,对土壤入渗能力及持水量有极大影响[33],同时,深层土壤受蒸发等作用扰动较小,因而土壤粉粒含量、黏粒含量对200~400 cm土层储水量影响也极为显著。
结合以上分析认为:今后黄土高原地区的造林工程,不仅要考虑树种耐旱能力,更应充分调查恢复区的地形、土质条件及生长季降水的分配情况,采取因地制宜的原则,使生态环境得到可持续发展。
3 结论
人工刺槐林、天然三角槭林和野艾蒿草地0~400 cm土层土壤水分平均变化范围分别为8.36%~9.63%、10.01%~13.19%、15.43%~19.17%,野艾蒿草地0~100 cm土壤含水量显著高于人工刺槐林和天然三角槭林(P<0.05),100~400 cm土壤含水量差异不显著。
不同植被类型和不同水文年的土壤水分活跃层均有差异,严重干旱年人工刺槐林、天然三角槭林和野艾蒿草地土壤水分活跃层下边界分别为60、80、40 cm。在平水年人工刺槐林及野艾蒿草地土壤水分活跃层深度均不足50 cm,但天然三角槭林可达120 cm。在中等湿润年,人工刺槐林、天然林和野艾蒿草地土壤水分活跃层分别可达140、180和140 cm。
土壤盈亏状况年际差异较大:严重干旱年,人工刺槐林、天然三角槭林和野艾蒿草地土壤水分亏缺程度严重土层分别为100~300、0~200和0~100 cm;平水年土壤水分的输入和输出达到平衡;中等湿润年,人工刺槐林、天然三角槭林和野艾蒿草地分别在0~100、100~180和0~100 cm土层土壤水分补给量最高,而在深层土层(200~400 cm)中,各植被类型下补给量较小,接近于0。
不同深度土层土壤储水量变化受不同环境因子主导:0~100 cm土层主要受植被类型和土壤容重控制;100~200 cm土层主要受容重和坡向控制;区域干湿状况、土壤黏粒和粉粒含量是200~400 cm深土层的主导因子。今后,黄土高原地区的造林工程,不仅要考虑树种的耐旱能力,更应充分考虑地形、土质及生长季降水分配的影响。
