诺丽果原粉对绒山羊体外瘤胃发酵功能的影响
2021-03-03张清月梁晓帅李胤豪马国强郭晓宇赵艳丽闫素梅
张清月 梁晓帅 李胤豪 马国强 郭晓宇 赵艳丽 闫素梅
(内蒙古农业大学动物科学学院,内蒙古自治区动物营养与饲料科学重点实验室,呼和浩特 010018)
内蒙古绒山羊作为世界著名的绒肉兼用型品种,其肉质细嫩、膻味小且营养价值高。但由于天然草场的营养物质季节性不平衡以及我国禁牧力度的不断加强,绒山羊的养殖逐渐由放牧转变为舍饲和半舍饲养殖。然而,集约化养殖易使绒山羊产生氧化应激,甚至引起营养代谢病,进而降低其生产性能。抗生素在解决上述问题时也带来了食品安全隐患,且饲料中的全面禁抗使得养殖产业中替抗产品的开发尤为需要。诺丽(MorindacitrifoliaL.,药名海巴戟)在我国东南部地区种植广泛,其果实含有丰富的营养物质和生物学活性物质,主要为多酚类、糖苷类和生物碱等,具有抗菌、消炎和提高免疫力等多种功效[1],极具开发为绿色饲料添加剂及新型替抗产品的前景。目前,国内外对于诺丽果的研究主要集中在其功能性成分的提取和分析上,以及以小鼠[2]、鸡[3]和犊牛[4]为模型研究其药理作用,或是研究其工业废弃物对奶山羊体外发酵的影响[5]。然而,将诺丽果作为饲料添加剂应用于绒山羊等反刍动物养殖领域的研究目前还未见报道。瘤胃作为反刍动物最重要的消化器官,其功能对于营养物质和能量的获取具有重要意义。但活体试验动物数量大、试验周期长且成本高,研究工作常受到限制。而体外瘤胃发酵可在离体环境下模拟瘤胃内的消化过程,且更为省时省力。因此,本试验利用体外瘤胃发酵结合多项组合效应值(MFAEI),研究饲粮中添加不同剂量的诺丽果原粉对绒山羊瘤胃发酵功能的调控作用,以期为诺丽果资源在饲料领域的综合利用提供基础,也为绒山羊养殖中替抗产品的开发提供新思路。
1 材料与方法
1.1 试验原料
诺丽果风干片由海南省五指山市诺丽果基地提供,于65 ℃烘干,粉碎后过40目筛制成诺丽果原粉。
3.系统内普法与社会普法不平衡。各级各部门在落实责任制过程中,存在系统内普法与社会普法不能并重的情况。许多单位积极开展社会普法,紧密结合本部门法律法规颁布实施纪念日、宣传日、纪念周等重要普法节点,组织开展各类有声有色的普法活动,但却忽视对系统内执法人员和工作人员的普法,执法工作人员法律素养不够高,法治意识不够强,开展“谁执法谁普法”的广度与深度不够,实效性欠缺。
1.2 试验动物和基础饲粮
选取3只体况相近、年龄为2.5岁的健康绒山羊羯羊(品系为内蒙古阿尔巴斯白绒山羊)作为瘤胃液供体羊。每天于09:00和15:00对供体羊进行饲喂,供体羊自由饮水,基础饲粮参照我国《肉羊饲养标准》(NY/T 816—2004)配制,基础饲粮组成及营养水平见表1。
于刚性轮,其中在空载工况中尤为明显,脱轨系数减小43.84%,超限时间减少26.09%。直线轨道上弹性轮和刚性轮的脱轨系数均未超过限值,弹性轮的最大值略小。
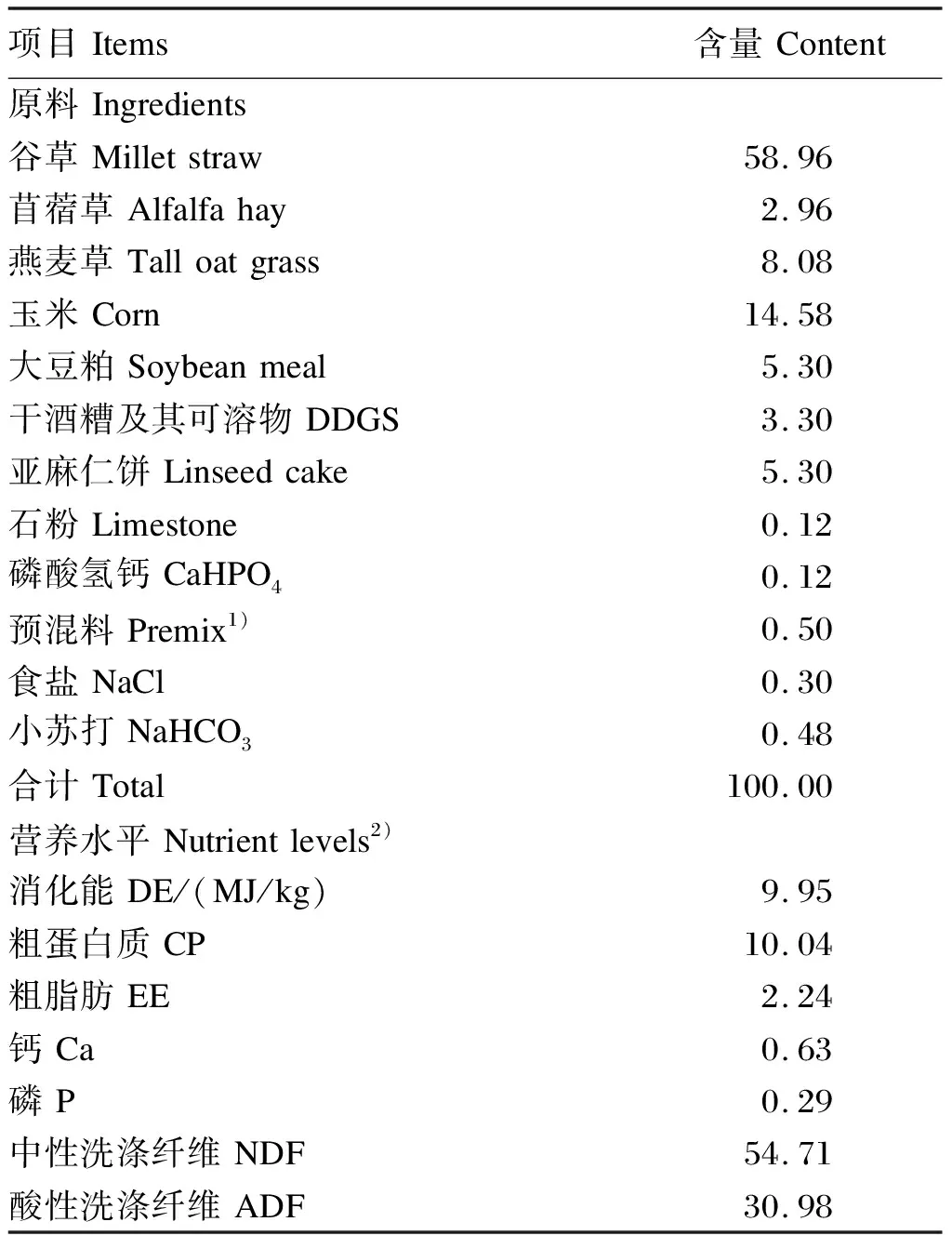
表1 基础饲粮组成及营养水平(风干基础)
1.3 试验设计
参考Menke等[7]的方法配制缓冲液,称取1 g发酵底物放入高压灭菌的发酵瓶中,于39 ℃预热。于晨饲前经口腔采集3只绒山羊的瘤胃液,混合后经1层纱布过滤至已预热达39 ℃并通有二氧化碳(CO2)的保温瓶中。将瘤胃液与缓冲液按1∶2的比例混合,并分装至上述玻璃瓶中,培养体系为60 mL,该步骤全程通CO2进行。将发酵瓶密封后于39 ℃气浴摇床培养,转速为120 r/min。培养时间分别为3、6、9、12和24 h,到达各时间点后,迅速将相应的培养瓶取出,冰浴终止发酵,并取样测定各项指标。
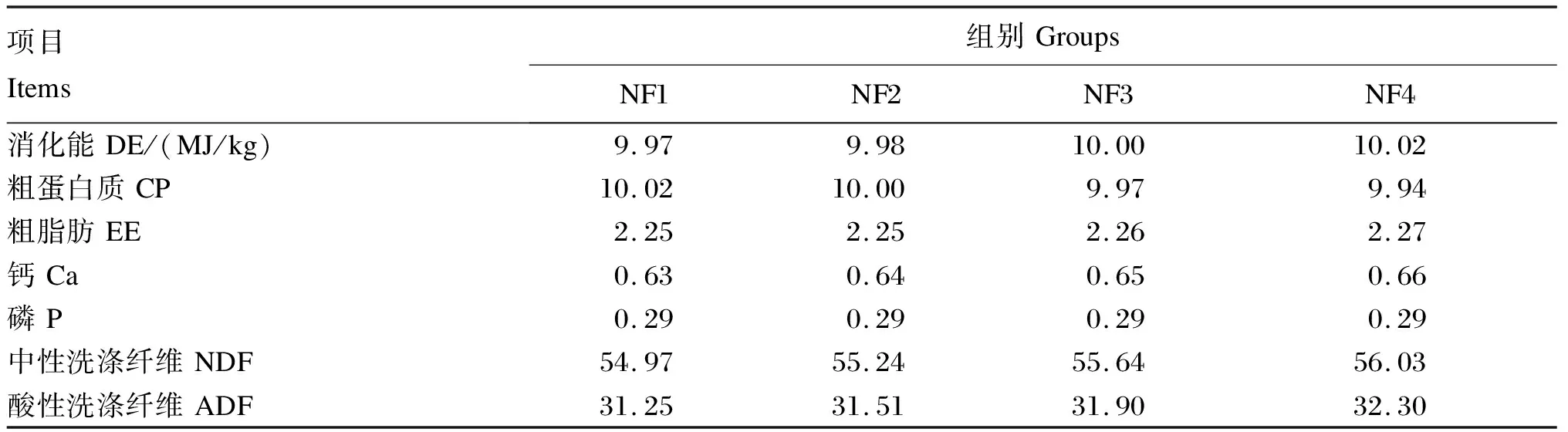
表2 NF1~NF4组试验饲粮的营养水平(风干基础)
1.4 体外培养方法
试验采用完全随机区组设计,设5个诺丽果原粉添加剂量,试验饲粮分别以0(对照组,CON组)、1.0%(NF1组)、2.0%(NF2组)、3.5%(NF3组)和5.0%(NF4组)的诺丽果原粉等比例替代基础饲粮中的玉米;并设5个培养时间点,分别为3、6、9、12和24 h;每个培养时间点设6个重复(即每个添加剂量有30个发酵瓶)。其中,CON组的发酵底物为基础饲粮,营养水平与瘤胃液供体羊基础饲粮的营养水平相同;NF1~NF4组试验饲粮的营养水平见表2。
1.5 测定指标与方法
粗蛋白质(CP)含量的测定参照《饲料中粗蛋白的测定 凯氏定氮法》(GB/T 6432—2018);粗脂肪(EE)含量的测定参照《饲料中粗脂肪的测定》(GB/T 6433—2006);钙含量的测定参照《饲料中钙的测定》(GB/T 6436—2018),采用高锰酸钾法;磷含量的测定参照《饲料中总磷的测定 分光光度法》(GB/T 6437—2018);中性洗涤纤维和酸性洗涤纤维含量的测定参考Van Soest等[8]的方法,采用ANKOM纤维分析仪;pH用CT-6022型手持便携式pH计测定;氨态氮(NH3-N)浓度的测定采用比色法[9];菌体蛋白(BCP)浓度的测定采用考马斯亮兰法[10];原虫数量的测定参考冯仰廉[11]的方法;挥发性脂肪酸(VFA)浓度的测定采用气相色谱法[12],以二乙基丁酸为内标,并计算总挥发性脂肪酸(TVFA)浓度;产气量采用ANKOM RFS体外产气系统进行测定;参考卢德勋[13]的计算方法,对单项组合效应值(SFAEI)及MFAEI进行计算,MFAEI是不同组合(处理)的各指标SFAEI之和,SFAEI的计算公式如下式:
用染色序列对P进行着色,实际上是对m(2n+1)+2n-1条边进行着色,而图4中色集合的个数有个,根据上述染色算法,当k是奇数时,有当k是偶数时,有因此恒成立,此时求得最小的整数k满足⎤。
1.6 统计分析
由表3可知,添加不同剂量的诺丽果原粉均显著降低了体外瘤胃发酵pH、NH3-N浓度和原虫数量(P<0.05);NF3组NH3-N浓度最低,显著低于其他各组(P<0.05);NF3组原虫数量也最低,显著低于NF1组(P<0.05)。随诺丽果原粉添加剂量的增加,NH3-N浓度和原虫数量均呈显著的一次线性和二次曲线下降(P<0.05),而pH也呈显著的一次线性和二次曲线下降(P<0.05)。添加不同剂量的诺丽果原粉显著提高了BCP浓度(P<0.05);其中,NF3和NF4组BCP浓度显著高于NF1组(P<0.05);NF3组BCP浓度最高,显著高于NF2组(P<0.05)。NF3和NF4组产气量显著高于CON组(P<0.05),且以NF3组最高。随诺丽果原粉添加剂量的增加,BCP浓度和产气量均呈显著的一次线性升高(P<0.05)。发酵时间对pH、NH3-N浓度、BCP浓度、原虫数量和产气量均有显著影响(P<0.05)。随着培养时间的延长,pH呈下降趋势;产气量呈上升趋势;NH3-N浓度、BCP浓度和原虫数量则呈先升高再下降的趋势,并在培养12 h达最高值。添加剂量与培养时间的交互作用对原虫数量有显著影响(P<0.05),培养3和6 h时各组原虫数量较少,培养12 h时CON组的原虫数量最多。
式中:m为各培养时间点(m=1,2,3,4,5);n为培养时间点的个数(n=5);A1m为组合前(对照组)各单一指标不同培养时间点的数值;A2m为组合后(试验组)各单一指标不同培养时间点的数值。
由表4可知,NF2和NF3组乙酸浓度显著高于CON组(P<0.05),且以NF3组最高,显著高于NF1和NF4组(P<0.05);NF1~NF4组丙酸、丁酸、异丁酸、戊酸和TVFA浓度显著高于CON组(P<0.05),乙酸/丙酸显著低于CON组(P<0.05)。NF3组丁酸、戊酸和TVFA浓度最高,其中,NF3组丁酸和TVFA浓度显著高于其他组(P<0.05),而戊酸浓度显著高于NF1组(P<0.05);NF2~NF4组异戊酸浓度显著高于CON组(P<0.05)。随着诺丽果原粉剂量的增加,乙酸和异丁酸浓度呈显著的二次曲线升高(P<0.05),乙酸/丙酸呈显著的二次曲线下降(P<0.05);戊酸和异戊酸浓度呈显著的一次线性升高(P<0.05),而丙酸、丁酸和TVFA浓度呈显著的一次线性和二次曲线升高(P<0.05)。发酵时间对乙酸、丙酸、丁酸、异丁酸、戊酸、异戊酸、TVFA浓度和乙酸/丙酸有显著影响(P<0.05)。随培养时间的延长,除乙酸/丙酸呈先下降、再趋于平缓、之后再下降的趋势;其他指标均呈上升趋势。添加剂量与培养时间的交互作用对TVFA浓度有显著影响(P<0.05),培养24 h时NF3组的TVFA浓度最高,培养3 h时各组的TVFA浓度较低。
2 结 果
2.1 诺丽果原粉对体外瘤胃发酵pH、NH3-N浓度、BCP浓度、原虫数量和产气量的影响
使用SAS 8.1对数据进行统计分析,采用MIXED模型进行方差分析和回归分析,混合模型包括诺丽果原粉的添加剂量、培养时间、添加剂量×培养时间的交互作用以及发酵瓶的随机效应。P<0.05表示差异显著。
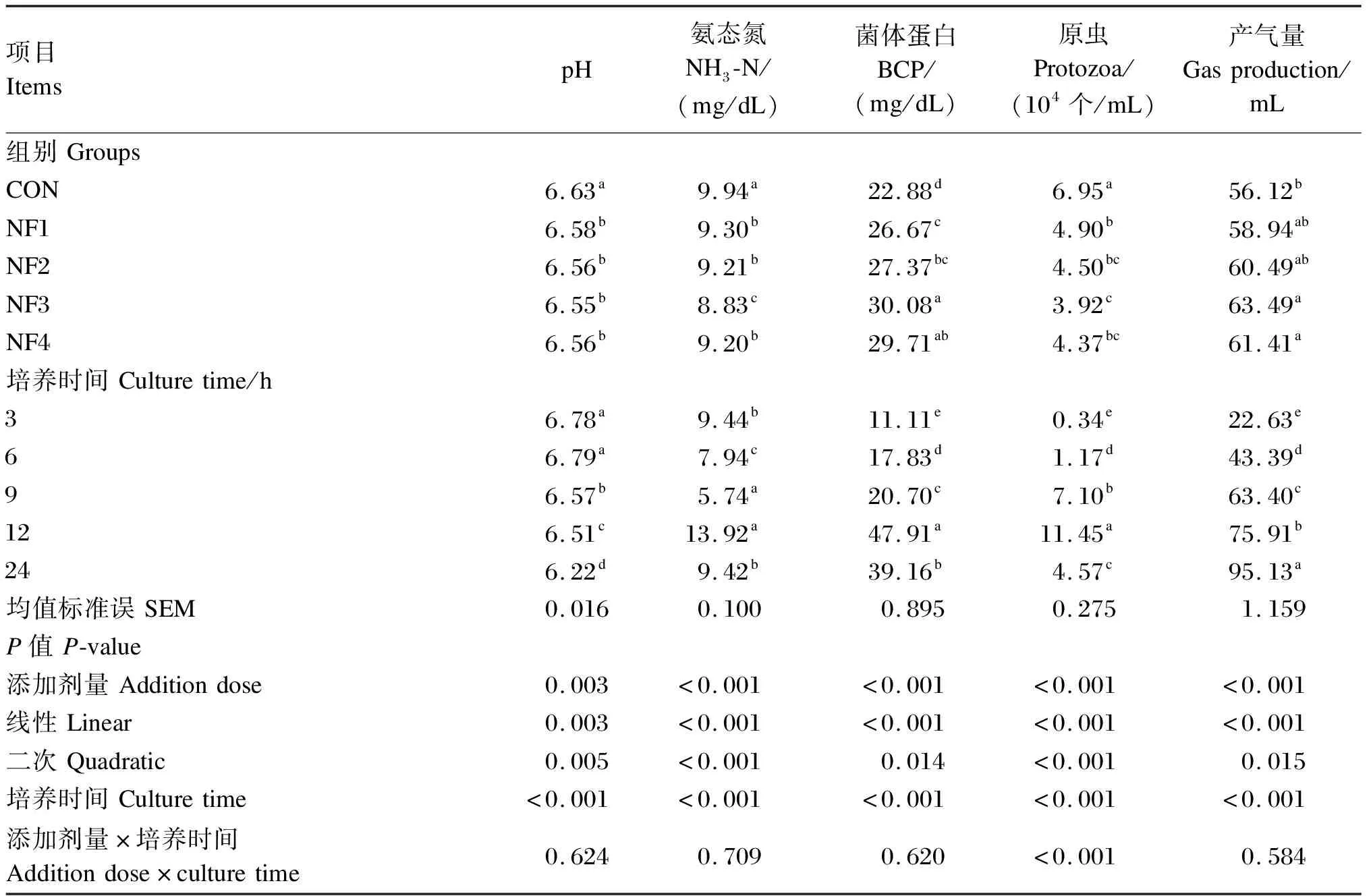
表3 诺丽果原粉对体外瘤胃发酵pH、NH3-N浓度、BCP浓度、原虫数量和产气量的影响
2.2 诺丽果原粉对瘤胃体外发酵VFA浓度的影响
3.死亡体重。据笔者的资料分析,仔猪初生重0.5 kg以下,哺乳期间死亡占死亡总数的80%以上,0.6~1.0 kg占13%,1.1 kg以上占6%。可见,仔猪初生重越小,死亡率越高。
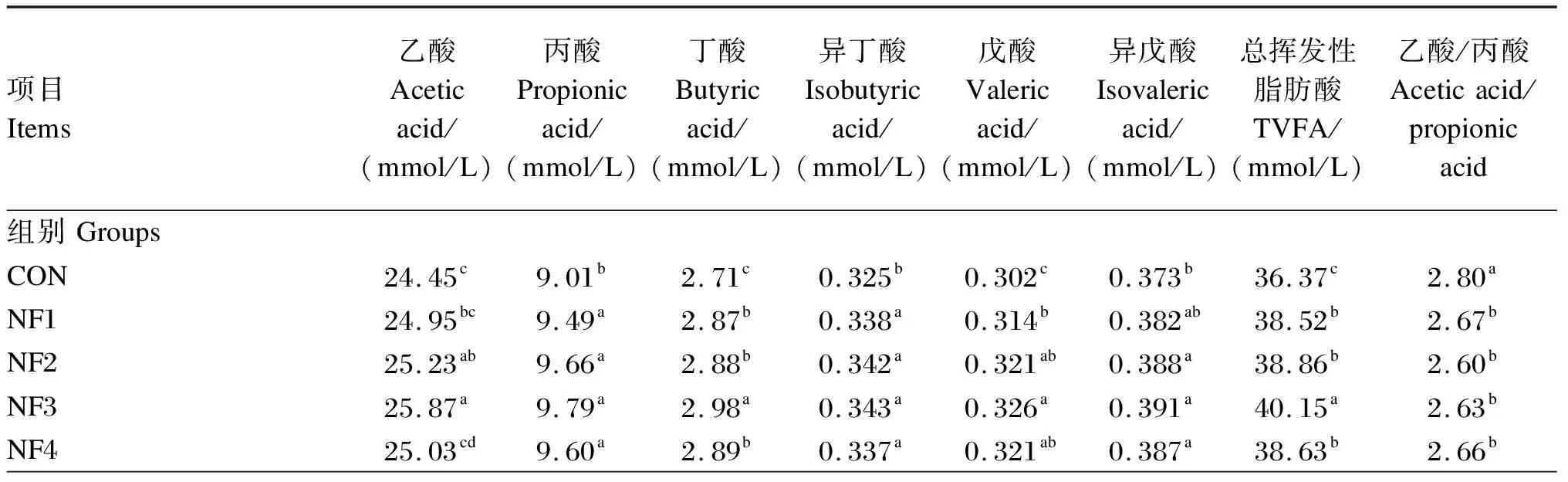
表4 诺丽果原粉对瘤胃体外发酵VFA浓度的影响
2.3 诺丽果原粉对瘤胃体外发酵MFAEI的影响
由表5可知,添加诺丽果原粉后,BCP浓度、TVFA浓度和产气量的SFAEI均显著升高(P<0.05);且NF3和NF4组BCP浓度的SFAEI显著高于NF1和NF2组(P<0.05);而TVFA浓度和产气量的SFAEI均以NF3组最高,且NF3组TVFA浓度的SFAEI显著高于其他各组(P<0.05),而产气量的SFAEI显著高于NF1和NF2组(P<0.05)。BCP浓度、TVFA浓度和产气量的SFAEI均随诺丽果原粉添加剂量的增加呈显著的线性和二次曲线升高(P<0.05)。并且,NF1~NF4组的MFAEI显著升高;以NF3组最高,显著高于其他各组(P<0.05);以NF4组次高,显著高于NF1和NF2组(P<0.05)。MFAEI随诺丽果原粉添加剂量的增加呈显著的一次线性和二次曲线升高(P<0.05)。
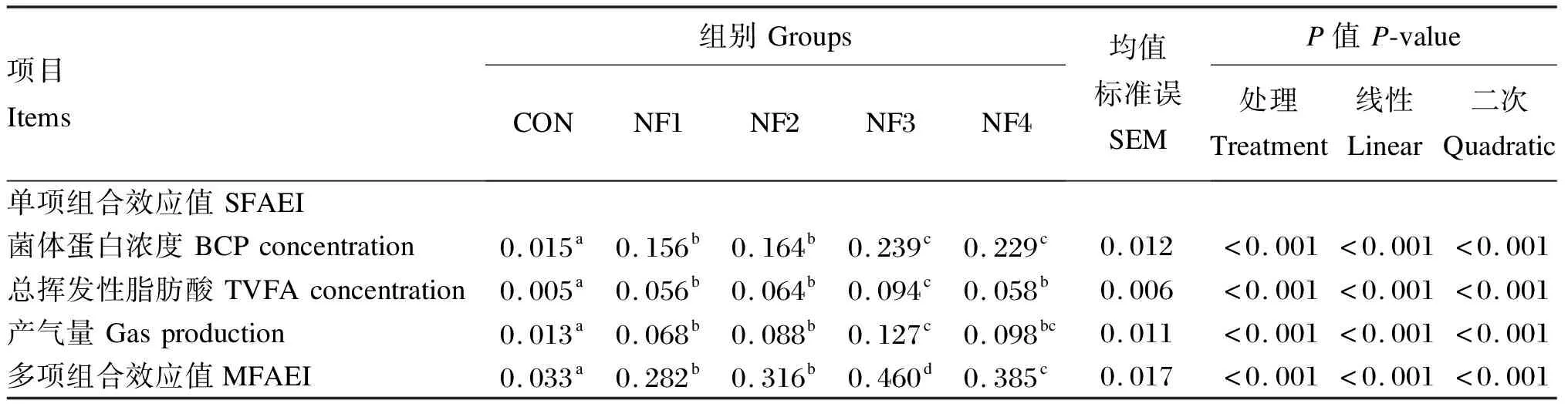
表5 诺丽果原粉对瘤胃体外发酵MFAEI的影响
3 讨 论
瘤胃液中的BCP浓度是综合评价蛋白质利用效率及微生物种群数量的重要指标,而NH3-N是合成BCP的主要氮源。因而在一定范围内NH3-N浓度的降低和BCP浓度的升高暗示着瘤胃内蛋白质合成效率的提高。所以,原虫的存在增加了氮在瘤胃内的周转和消耗,去原虫更有利于提高反刍动物的氮利用效率[14]。合成优质BCP也需要适宜的pH维持瘤胃发酵稳态,pH在6.0~7.0之间利于BCP的合成[15]。在本试验中,添加诺丽果原粉后pH虽下降,但仍在BCP合成的适宜范围内;而NH3-N浓度和原虫数量的下降以及BCP浓度的增加,则表明添加诺丽果原粉可以促进体外瘤胃发酵的氮降解并提高机体对氮的利用效率。关于本试验中诺丽果原粉的降原虫数量功能,有研究表明每100 g成熟诺丽果中的皂苷含量约为236.0 mg[16],且皂苷类物质可通过破坏原虫细胞膜杀死原虫[17],因而诺丽果原粉的降原虫作用可能与其含有皂苷类物质有关。
产气量是间接评定饲料样品营养价值及其在瘤胃内发酵状况的指标[18];而VFA是反刍动物的主要能量来源,也是瘤胃微生物重要的碳架来源,其浓度与组成是反映瘤胃消化及其代谢能力的重要指标[19]。本试验中,产气量和VFA浓度显著升高,说明添加诺丽果原粉可显著促进饲料样品体外瘤胃发酵及其营养物质代谢;而乙酸/丙酸显著下降,则说明诺丽果原粉可优化VFA组成,提高能量利用效率,为BCP合成提供充足碳源。分析本试验中pH降低的原因,可能与诺丽果原粉中有机酸含量较高[20]或培养液中VFA浓度增加有关;也可能是因为培养液中原虫数量的减少,致使原虫因储存淀粉而稳定pH的作用减弱[21],进而导致体外发酵pH的下降。诺丽果促进瘤胃发酵的机理尚不清楚,这可能与诺丽果中含有多糖、酚类等活性物质有关[22]。目前尚未见这方面的研究报道,但关于其他多糖的体内外研究表明,黄芪多糖[23]、苦瓜多糖[24]、香菇多糖或木聚糖[25]等均可促进瘤胃发酵。因而,可能是诺丽果多糖调控了瘤胃微生物多样性及其代谢产物,进而影响了瘤胃发酵,需要进一步研究探讨。
MFAEI可反映饲料间的整体互作,其数值越大,对应的饲料组合越有利于瘤胃发酵以及营养组分的利用。因此,可利用MFAEI判断饲料的适宜配比。本试验的各指标回归分析结果表明,随诺丽果原粉添加剂量的增加,pH、BCP浓度、产气量和异戊酸浓度呈显著的一次线性变化趋势,其他指标呈显著的二次曲线剂量依赖效应,且极值均出现在NF3组。虽然MFAEI的一次线性和二次曲线升高趋势均显著,但以NF3组最高,NF4组次高,且二者差异显著。这说明诺丽果原粉的添加剂量为3.5%时,促进瘤胃发酵的效果较好,而当添加剂量增至5.0%时,促进瘤胃发酵的效果减弱。MFAEI对诺丽果添加剂量的二次曲线响应是可以预见的,因为高剂量的诺丽果所具有的抗菌性更强,因而可能会使其促瘤胃发酵作用减弱。类似的,无籽诺丽果渣在体外发酵中促TVFA生成的作用与其添加剂量呈显著的二次曲线升高趋势,最佳添加剂量为10%~15%,而添加20%~25%时TVFA浓度与对照组无显著差异[5]。饲粮中添加沙棘果渣可促进体外瘤胃发酵,但添加24%时的TVFA浓度显著低于添加16%时[26]。目前,关于诺丽果促进瘤胃发酵的机理尚不清楚,今后需进一步从其多糖、酚类物质等方面,研究诺丽果促瘤胃发酵的功效物质,并结合微生物多样性等多组学联合分析技术深入探究诺丽果促瘤胃发酵的机理。
4 结 论
饲粮中添加诺丽果原粉可促进阿尔巴斯白绒山羊的体外瘤胃发酵功能,并降低原虫数量,且均在添加剂量为3.5%时效果较好。
