6种高寒禾草对干旱胁迫的生理响应及抗旱性评价
2021-03-02蔺亚平杨成行苏家豪包爱科
蔺亚平,杨成行,苏家豪,马 清,包爱科
(兰州大学草地农业科技学院, 甘肃 兰州 730000)
近年来,随着气候变暖和人口迅速增长,水资源短缺已成为全球面临的严峻挑战,干旱土地面积的逐年增加已成为世界各国关注的问题[1]。我国是受干旱影响最为严重的国家之一,干旱已成为制约我国农业生产和畜牧业发展的重要因素。水资源匮乏使我国西北高海拔地区农牧业面临严峻考验,不仅对农作物的存活、生长和生理生化代谢产生影响,而且伴随着草地沙化面积的不断扩大,草地逐渐减少,生态环境问题日益突出[2]。
对于我国北方高寒草甸草原植物,低温是影响其生长的重要因子,但水分的作用亦不可忽视,即使在降水多的地区也普遍存在季节性和非周期性干旱问题[3]。牧草抗旱性的强弱是维系草地资源可持续发展和改善生态条件的关键因素,近年来众多研究多用生长状况、相对含水量、叶绿素含量、渗透调节物质的积累、细胞膜透性、活性氧产生速率、抗氧化系统活性等指标评价牧草的抗旱性,取得了一系列进展。研究表明,干旱胁迫显著抑制了芒草(Miscanthus sinensis)[4]、黑麦草(Lolium perenne)[5]、披碱草(Elymus dahuricus)[6]的生长,使其生物量显著降低。干旱胁迫下,高粱(Sorghum bicolor)[7]、蒙古冰草(Agropyron mongolicum)、无芒雀麦(Bromus inermis)[8]等牧草幼苗相对含水量逐渐下降,细胞膜透性逐渐上升;狼尾草(Peuuisetum alopecuroides)、蓝羊茅(Festuca glauca‘Freddy’)[6]、 苏 丹 草 (Sorghum sudanense)[9]等牧草幼苗叶绿素含量均呈下降趋势,从而影响牧草的生长发育。随干旱胁迫时间延长,树锦鸡儿(Caragana arborescens)、砂生槐(Sophora moorcroftiana)[10]和紫花苜蓿(Medicago sativa)[11]幼苗叶片中H2O2和丙二醛(malondialdehyde, MDA)含量不断增加。干旱胁迫处理下草地早熟禾(Poa pratensis)[12]、猫尾草(Uraria crinita)和扁穗冰草(Agropyron cristatum)[13]叶片内超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase, CAT)和抗坏血酸过氧化物酶(ascorbate peroxidase, APX)活性显著增强。综上,干旱胁迫可通过影响牧草农艺性状、形态特征和生理生化等指标,从而严重影响其正常生长发育和产量形成。这些指标的变化在抗旱能力不同的牧草中存在着一定程度的差异,因此综合运用多种指标来衡量牧草的抗旱性能,将有望得到更为精确的结果。
禾本科牧草(简称禾草)是天然草地植被的重要组成成分,也是栽培草地的重要建植种,不仅在退耕还林还草、土壤改良、水土保持等生态恢复建设中发挥着重要作用,而且以产量高、品质优、适口性好、适应性广等特点,在我国草牧业发展中扮演着重要角色[3,14]。禾本科牧草种类繁多,在不同的气候环境下其抗性表现出较大差异。但目前我国高寒地区草地恢复在选择草种时,多考量其耐寒性,对于其抗旱性评价的研究较少。高寒地区草地退化类型多样,草地沙化是其中之一,且有逐年增加的趋势,在抗寒禾草中选择抗旱性强的品种将有助于高寒地区沙化草地的快速恢复。鉴于此,本研究以高寒地区草地恢复中主推的6 种禾草为材料进行盆栽干旱胁迫,分析其生物量、叶片水分状况、细胞膜透性、叶绿素含量、超氧化物歧化酶和过氧化物酶活性等指标对干旱胁迫的响应,并用隶属函数法综合评价6 种禾草的抗旱性,以期筛选出抗旱性强的禾草品种,为高寒地区沙化草地的恢复奠定基础。
1 材料与方法
1.1 植物材料
本研究所用6 个禾草品种均为青藏高原主推品种,种子由青海大学刘文辉研究员提供(表1)。

表1 试验品种Table 1 Experimental varieties
1.2 试验处理
采用盆栽试验,挑选籽粒饱满、无残缺的禾草种子,用10%高锰酸钾溶液消毒10 min,蒸馏水反复冲洗5~6 次,用滤纸吸干后,按照15 g·m-2的密度撒播,均匀地置于以蛭石为基质的塑料盆(15 cm ×17 cm)中,每种禾草种植15 盆。发芽后用1/2 Hoagland营养液每2 d 浇灌一次,培养室昼夜温度为(25 ± 2)℃/(23 ± 2) ℃,光照时间为16 h·d-1,空气相对湿度约为65%,光照强度约为800 μmol·(m2·s)-1。
所有材料培养1 个月后,每个品种选长势良好的12 盆植株,一次性浇透水,然后通过称重法进行控水处理。设置对照组及干旱处理组,对照组将基质含水量维持在70%饱和含水量,干旱处理组停止浇水,直至基质含水量降至30%饱和含水量,每2 d称重一次,当土壤含水量低于设置范围时,采用缓慢浇水法补充水分,对照组和处理组维持相应的饱和含水量,28 d 后取样测定相关形态生理指标,所有指标6 个重复。
1.3 测定指标及方法
地上部干重、叶片组织含水量均采用称重法测定[15];叶绿素含量测定采用乙醇丙酮混合液浸提法[16];细胞膜透性采用电导率法测定,以细胞膜外渗率表示细胞膜透性的变化[17];超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法测定[17];过氧化物酶(POD)活性采用愈创木酚法测定[17]。
1.4 抗旱性综合评价
采用隶属函数法[18]对6 种禾草苗期抗旱能力进行综合评价,以干旱胁迫下各指标的相对变化值为抗旱性评价值。计算公式如下:

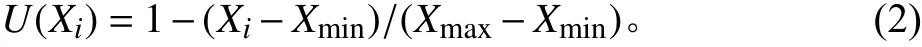
式中:U(Xi)为各个指标的隶属值;Xi表示第i个指标的相对变化值,Xi= (干旱胁迫处理的平均值-对照值)/对照值;Xmax表示所有品种第i个指标相对变化值的最大值;Xmin表示所有品种第i个指标相对变化值的最小值。如果i指标的相对变化值与植物的抗旱性呈正相关关系,用式(1);如果i指标的相对变化值与植物的抗旱性呈负相关关系,则用式(2)。对每种禾草各指标的抗旱隶属值进行累加,并计算出平均值,平均值越大,植物的抗旱性越强,平均值越小,抗旱性越弱[14]。
1.5 数据处理
用SPSS 26.0 软件对数据进行独立样本T检验,差异显著性水平设置为P< 0.05 和P< 0.01,数据以平均值 ± 标准误表示。
2 结果与分析
2.1 干旱胁迫下6 种禾草的形态变化
干旱处理28 d 后,6 种禾草的长势均明显弱于对照组,生长受到了抑制。相比而言,同德老芒麦、青牧1 号老芒麦和同德短芒披碱草长势显著好于青海扁茎早熟禾、青海冷地早熟禾和青海中华羊茅(图1)。

图1 干旱胁迫下6 种禾草的长势Figure 1 The growth phenotype of six varieties under drought stress
在干旱胁迫下,青海冷地早熟禾和青海中华羊茅的地上部干重与各自对照相比分别极显著下降了18.6%和37.6% (P> 0.01)。与对照相比,青海扁茎早熟禾、同德老芒麦、青牧1 号老芒麦和同德短芒披碱草在干旱胁迫下的地上部干重虽较对照呈现下降趋势,但并未达到显著差异(P> 0.05),其中青海扁茎早熟禾地上部干重下降率最小,为4.3% (表2)。
干旱胁迫下,青海冷地早熟禾和青海中华羊茅的叶片组织含水量较对照相比显著下降了3.6%和1.6% (P< 0.05);而青海扁茎早熟禾、同德老芒麦、青牧1 号老芒麦和同德短芒披碱草在干旱胁迫下叶片组织含水量变化不显著(P> 0.05),其中同德老芒麦的叶片组织含水量下降率最小,为0.1% (表2),说明这4 种禾草在干旱胁迫下能维持其体内正常的水分状态。

表2 干旱胁迫下6 种禾草的地上部干重和叶片组织含水量Table 2 The shoot dry weight and tissue water content of leaves of six varieties under drought stress
2.2 干旱胁迫对6 种禾草生理指标的影响
与对照相比,干旱胁迫下6 种禾草的叶绿素含量均有所下降,其中青海扁茎早熟禾和青海冷地早熟禾的叶绿素含量与对照相比差异不显著(P>0.05),而同德老芒麦、青牧1 号老芒麦、青海中华羊茅和同德短芒披碱草的叶绿素含量则比其各自对照分别显著或极显著下降了30.7%、23.3%、77.6%和34.6% (图2),可见干旱胁迫对这4 种禾草的叶绿素合成影响较大。

图2 干旱胁迫下6 种禾草的叶绿素含量、相对电导率、超氧化物歧化酶和过氧化物酶Figure 2 The chlorophyll content, relative electrical conductivity, superoxide dismutase (SOD) and peroxidase (POD) activity of six varieties under drought stress
在干旱胁迫下,6 种禾草的叶片相对电导率均有所增加,但青海扁茎早熟禾叶片相对电导率的增加幅度最大、数值也最高,高于其他5 种禾草;与各自对照相比,青海扁茎早熟禾和同德老芒麦的叶片相对电导率分别增加了180%和90%倍,差异达到极显著水平(P< 0.01),但干旱胁迫下青牧1 号老芒麦的叶片相对电导率值最低,为4.8%,且与对照相比,差异并未达到显著水平(P> 0.05) (图2),说明其细胞膜受损程度较轻。
干旱胁迫下,青海冷地早熟禾、同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的SOD 活性均有所增强,其中干旱胁迫下同德老芒麦的SOD 活性比对照增加了99.5%,差异达到显著水平(P< 0.05);而青海扁茎早熟禾和青海中华羊茅在干旱胁迫下SOD 活性有所降低,其中青海中华羊茅的SOD 活性最低(图2)。
干旱胁迫下6 种禾草的POD 活性均呈增加趋势,其中青牧1 号老芒麦、青海中华羊茅和同德短芒披碱草的POD 活性分别较其各自对照极显著增加了183.3%、47.0%和53.4% (P< 0.01);相比而言,无论在对照条件还是干旱胁迫下,同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的POD 活性高于青海扁茎早熟禾、青海冷地早熟禾和青海中华羊茅(图2)。
2.3 抗旱性综合评价
利用6 个生长和生理指标的相对变化率进行综合分析,分别计算各指标在不同品种中的隶属值,并以各品种的平均隶属值作为鉴定其抗旱性强弱的综合评价指标。结果显示,6 种禾草的隶属函数值存在一定差异,其平均值介于0.325~0.838。根据隶属函数值越大,抗旱性越强的原则,本研究中6 种禾草的抗旱性由强到弱依次为青牧1 号老芒麦、同德老芒麦、同德短芒披碱草、青海冷地早熟禾、青海扁茎早熟禾、青海中华羊茅(表3)。

表3 6 种禾草的抗旱性综合评价Table 3 Comprehensive evaluation of drought resistance among six varieties
3 讨论
干旱对牧草的危害是一个复杂的生理过程,而牧草抵抗逆境胁迫也是一个多系统的综合生理反应。研究表明,在干旱胁迫下,牧草通常通过改变其生长状态和调整各部分生物量以适应干旱环境,增强其生态适应性[19],主要表现为植株紧凑、低矮、叶片萎缩减少蒸发面积等来保证体内水分均衡。牧草生产的主要指标是生物产量,牧草在整个生长发育过程中会不断通过调整生物量的分配来适应和抵御外界环境的变化及不良影响,从而将伤害系数降到最低[20]。本研究表明,在干旱胁迫下,6 种禾草的生长受到了不同程度的抑制,长势均明显弱于对照组,地上部干重和叶片组织含水量与对照相比也有所降低,其中同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的长势明显优于青海扁茎早熟禾、青海冷地早熟禾和青海中华羊茅,表现出较强的抗旱性。这与闫天芳等[21]对西藏4 份野生披碱草属牧草幼苗的抗旱性研究以及郭郁频等[22]在不同早熟禾品种上的抗旱性研究得出的结论一致,说明在干旱胁迫下抗旱性较强的牧草能更好地维持体内的干物质含量,保持其体内正常的水分状态,以减轻干旱胁迫的伤害,从而保持较好的生长状况。
叶绿素含量是评价植物抗旱能力的重要指标。干旱胁迫会引起植物气孔的关闭,导致叶绿体超微结构和内囊体膜蛋白稳定性的改变,降低体细胞内叶绿素含量和光合碳循环中相关酶的活性,从而抑制光合作用[23]。叶绿素也会动态地调节比例,合理利用光能,以此反映牧草适应干旱胁迫的能力[24]。本研究中干旱胁迫下6 种禾草叶绿素含量的变化趋势与李京蓉等[24]对高寒牧区禾本科牧草抗旱性研究以及潘昕等[10]对青藏高原6 种植物抗旱性能分析的研究结果一致,即干旱胁迫导致禾草的叶绿素含量均有所下降,但下降趋势在不同品种间存在差异,这种差异可能与其自身的生物学特性和干旱胁迫发生的强度、时期或持续时间有关。本研究中,青海中华羊茅叶绿素含量的下降幅度最大,说明干旱胁迫对其叶绿素合成造成了较大破坏。
细胞膜是细胞维持胞内稳定代谢环境的必要屏障,其选择透过性可以调节物质的进出。干旱胁迫不仅会大幅降低叶片的含水量和叶绿素含量,使细胞膜渗透压上升,影响植物正常的生理活动,而且还会导致膜结构发生改变,使细胞受到严重损伤[25]。测定外渗液电导率的变化能够反映出质膜的受损程度和植物抗逆性的大小,通常认为,在持续干旱作用下电导率会逐渐上升,而上升的幅度有所不同,抗旱性较强的牧草品种其电导率上升幅度较小,抗旱性差的牧草品种其电导率上升幅度较大[26]。本研究结果表明,干旱胁迫下6 种禾草的相对电导率均有所增加,但青海冷地早熟禾、青牧1 号老芒麦、青海中华羊茅和同德短芒披碱草的相对电导率与对照相比差异并不显著,均维持在较稳定的水平,增幅较小;此外,无论在对照条件还是干旱胁迫下,同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的相对电导率低于其他品种。这说明青牧1 号老芒麦和同德短芒披碱草的细胞膜在干旱胁迫下受损程度较小,较为完整,有较强的防止膜脂过氧化作用的能力,对干旱胁迫的适应能力可能强于其他品种,这与王平等[27]在披碱草属牧草上的研究结果相类似。
SOD 和POD 是植物体内保护酶系统的重要组分,能有效清除自由基,降低膜质过氧化水平,从而保护细胞免受氧化损伤[28]。SOD 作为膜保护的第一道防线,催化超氧阴离子发生歧化反应,将毒性较强的超氧阴离子转化为毒性较轻的H2O2,POD 则是利用各种基质作为电子供体将H2O2还原为H2O[29-30]。在活性氧清除体系中SOD 是关键酶,随干旱胁迫强度的增加,SOD 活性呈先增加后降低的趋势[31],而POD 与SOD 活性在干旱胁迫下表现趋势相同,抗旱性强的品种,其根系POD 活性维持在较高的水平,且其SOD 活性的降低速率比抗性弱的草种缓慢[32]。本研究表明,干旱胁迫下,青海冷地早熟禾、同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的SOD 活性均有所增强;6 种禾草的POD 活性均呈现出增加的趋势,其中青牧1 号老芒麦、青海中华羊茅和同德短芒披碱草的POD 活性与对照相比差异均达到显著水平;且干旱胁迫下同德老芒麦、青牧1 号老芒麦和同德短芒披碱草的POD 活性高于青海扁茎早熟禾、青海冷地早熟禾和青海中华羊茅。这说明干旱胁迫下上述禾草会启动保护酶防御系统,增加体内POD 与SOD 活性,且这两种酶通过协同作用清除植物体内过剩的活性氧,从而减轻过氧化伤害,以维持细胞膜的完整性,使得同德老芒麦、青牧1 号老芒麦和同德短芒披碱草表现出较强的抗旱能力,这与8 种从美国引进的禾本科牧草保护酶活性与抗旱性研究结果[13]基本一致。
植物抗逆性的强弱是形态变异、生理变化、生化代谢和基因表达调控等多种因素综合作用的结果,抗旱能力是植物的一种复合性状[27]。干旱胁迫下植株的生长和生理生化指标的变化幅度因品种而异,不同指标反映出的结果不一致,无法以单一指标来反映植物的抗旱能力。因此,综合利用多个指标进行分析可提高植物抗旱性评价的准确性,近年来被众多研究者所采用。本研究采用禾草地上部干重、叶片组织含水量和生理指标的相对变化率,结合隶属函数法对6 种禾草的抗旱性进行了综合评价,消除了个别指标带来的片面性,使6 种禾草的抗旱差异具有可比性,从而提高了禾草抗旱鉴定结果的准确性和可靠性。分析得出本研究中青牧1 号老芒麦、同德老芒麦、同德短芒披碱草的抗旱性强于青海冷地早熟禾、青海扁茎早熟禾和青海中华羊茅,这与6 种禾草的形态观测和生理指标测定结果总体变化趋势相符。
4 结论
干旱胁迫下,6 种禾草其地上部生物量、叶片组织含水量和叶绿素含量均有所下降,细胞膜透性逐渐上升,除青海扁茎早熟禾和青海中华羊茅在干旱胁迫下SOD 活性有所降低外,其余禾草抗氧化酶活性升高,说明干旱胁迫能够诱导禾草体内抗氧化酶活性提高,保护细胞膜结构的完整性,增强禾草的抗旱性。但6 种禾草的抗旱性存在差异,同德老芒麦、青牧1 号老芒麦和同德短芒披碱草在干旱胁迫环境下能够维持较高的抗氧化酶活性,抗旱能力整体表现良好,可作为高寒地区草地恢复中主推的抗旱品种。
