γ-聚谷氨酸对小尾寒羊瘤胃发酵及菌群结构的影响
2021-03-01曹琪娜张艳梅敖长金张腾龙张兴夫赵亚波齐景伟
曹琪娜,张艳梅,敖长金,张腾龙,张兴夫,白 晨,赵亚波,齐景伟
(内蒙古自治区动物营养与饲料科学重点实验室,内蒙古农业大学动物科学学院,内蒙古呼和浩特 010018)
γ-聚谷氨酸(Poly-γ-glutamate,γ-PGA)是由D-谷氨酸和L-谷氨酸通过γ-谷氨酰键结合形成的一种可生物降解的水溶性阴离子聚合物[1],由传统发酵食品(日本纳豆等)中的芽孢杆菌类(主要为枯草芽孢杆菌、地衣芽孢杆菌)发酵而成,其分子中富含-COOH、-CO-、-NH-等多种活性基团,电荷密度高,具有优良的保水性、可食用性、缓释性和生物相容性等,降解后为小分子氨基酸,不会带来二次污染,是一种新型高分子材料[2]。作为一种可食用、无毒无害的生物聚合物,γ-PGA 及其衍生物被广泛应用于食物增稠剂、保湿剂、苦味消除剂、冷冻保护剂、药物载体、重金属吸收剂和动物饲料添加剂等物质中。研究指出,作为食品补充剂,γ-PGA 在体内可以通过增加钙的溶解度和肠道吸收来有效地提高钙的生物利用度,从而减少人体的骨质流失[3]。添加γ-PGA于动物饲粮中可促进机体对钙等矿物质的吸收,有利于降低饲料中矿物质的添加量,并能增加蛋壳强度和减少动物体内脂肪的沉积[4]。Jin 等[5]研究发现,口服γ-PGA可有助于将小鼠肠道微生物菌群调节为益生菌来增加小鼠肠道内乳酸菌的丰度,同时减少梭菌数量,具有稳定小鼠肠道微生物和增加肠道微生物组成多样性的作用。γ-PGA 作为饲料添加剂在反刍动物上的研究鲜有报道。因此,本试验通过16S rDNA 高通量测序分析技术,探讨饲粮中添加γ-PGA 对小尾寒羊瘤胃菌群结构的影响,为其作为饲料添加剂的开发利用提供理论依据。
1 材料与方法
1.1 试验动物的选择与饲养管理 本试验选取检疫合格、体重为(37.1±0.5)kg 的3 月龄小尾寒羊公羊24只,采用全混合饲粮(TMR),日饲喂2 次(07:00、17:00),自由采食,全天自由饮水。
1.2 试验药品与仪器设备γ-聚谷氨酸购买自西安瑞盈生物科技有限公司,分子质量为20 ku 的冷冻干燥粉,纯度为99%。DNA 提取试剂盒(南京建成生物工程研究所)、pH 计(贝尔分析仪器有限公司,BPH-7200)、全自动酶标仪(美国BIOTEK 公司,Synergy HT 型)、瘤胃液采样器(上海硅狄科学仪器有限公司,GCYQ-1-B)及气相色谱仪(美国Agilent 公司,6890N)。
1.3 试验设计与基础饲粮 采用单因素完全随机试验设计,将小尾寒羊随机分成对照组和试验组,试验预试期14 d,正试期7 d。对照组饲喂基础饲粮,试验组在基础饲粮中添加20 g/(只·d)γ-PGA。基础饲粮参考《肉羊饲养标准》(NY/T816-2004)配制,其中NaHCO3按20 g/(只· d)添加,基础饲粮组成及营养成分见表1。
1.4 瘤胃发酵参数测定
1.4.1 瘤胃液的采集与处理 在正试期第7 天晨饲2 h后通过瘤胃液采样器采集试验羊的瘤胃液,并将采集到的瘤胃液用4 层纱布过滤(在实验室内进行)后先用pH 计测定瘤胃液pH,再将滤液等体积充分混匀后转入冻存管后立即投入液氮罐中临时保存,再将其转移到实验室后放入-80℃冰箱冻存备用。对照组和试验组每组共有6 个样品,将样品送至上海桑格信息技术有限公司进行分析。
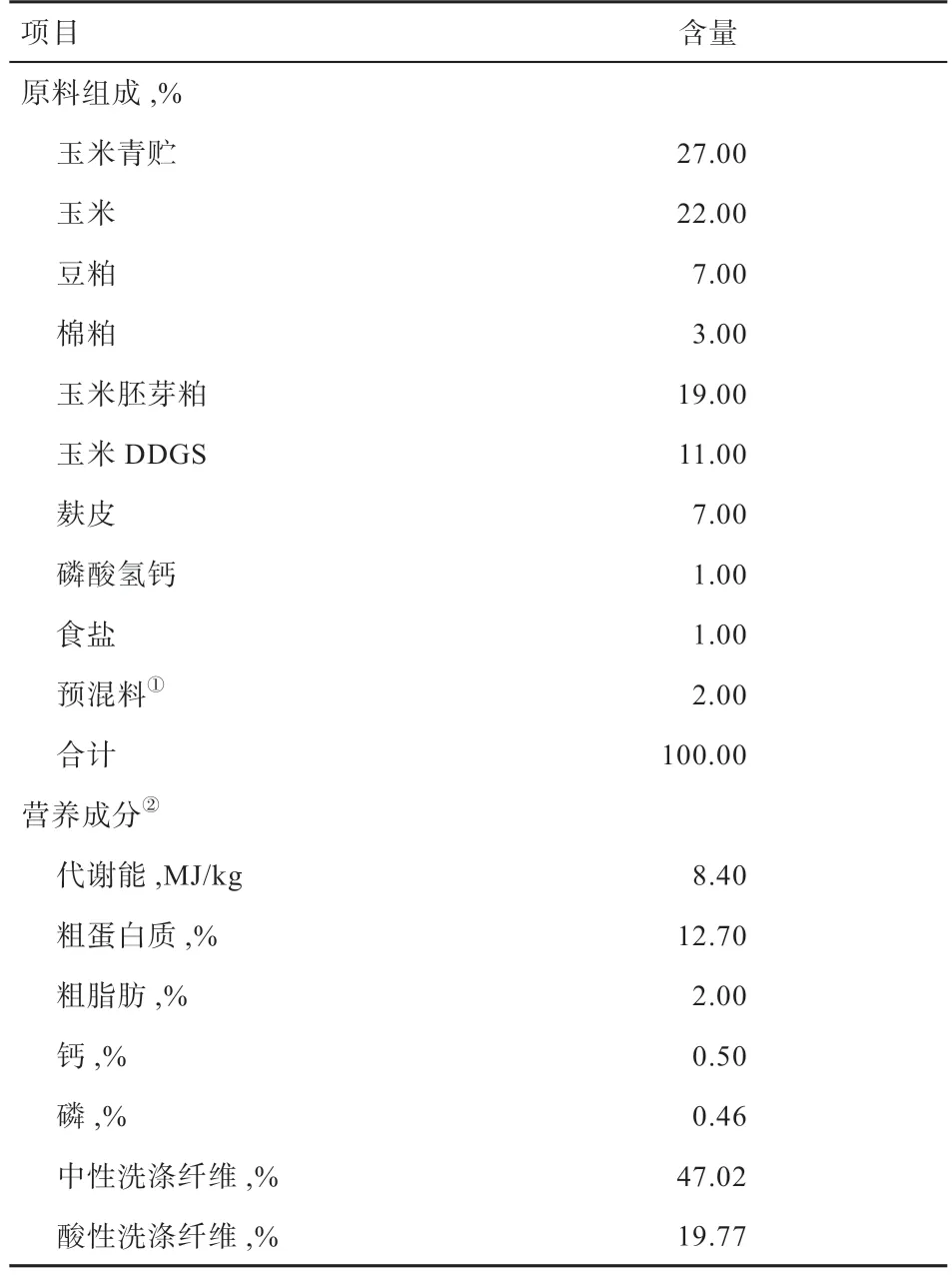
表1 基础饲粮组成及营养成分(干物质基础)
1.4.2 测定指标 粗蛋白质(CP)、粗脂肪(EE)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)、钙(Ca)、磷(P)及代谢能(ME)参照张丽英等[6]的方法测定,氨态氮(NH3-N)参照冯宗慈等[7]的方法测定,微生物蛋白(MCP)和挥发性脂肪酸(VFA)均参照周奕毅[8]的方法(气相色谱仪法)测定。
1.5 DNA 样品提取、扩增和Miseq 测序
1.5.1 DNA 的提取 使用DNA 提取试剂盒并按照试剂盒说明书严格操作,提取出样品中的DNA。
1.5.2 PCR 扩增 样品DNA 扩增目的片段为V3+V4 区,其引物为338F-806R,引物序列为338F:5'-ACTCCTA CGGGAGGCAGCAG-3'、806R:5'-GGACTACHVGGG TWTCTAAT-3'。
1.5.3 Miseq 测序 由上海桑格信息技术有限公司使用Miseq 2500 PE 250 平台上机测序样品Miseq。
1.6 统计分析 使用SAS 9.0 软件进行统计分析,采用t检验方法分析处理试验数据,分析结果以平均值±标准差表示,P<0.05 定义为差异显著。
2 结果与分析
2.1γ-PGA 对小尾寒羊瘤胃发酵参数的影响 由表2 可知,对照组与试验组小尾寒羊的瘤胃液pH、丙酸、丁酸、乙酸/丙酸及MCP 间无显著差异。试验组小尾寒羊的瘤胃液NH3-N(P<0.01)、乙酸(P<0.05)浓度均高于对照组。
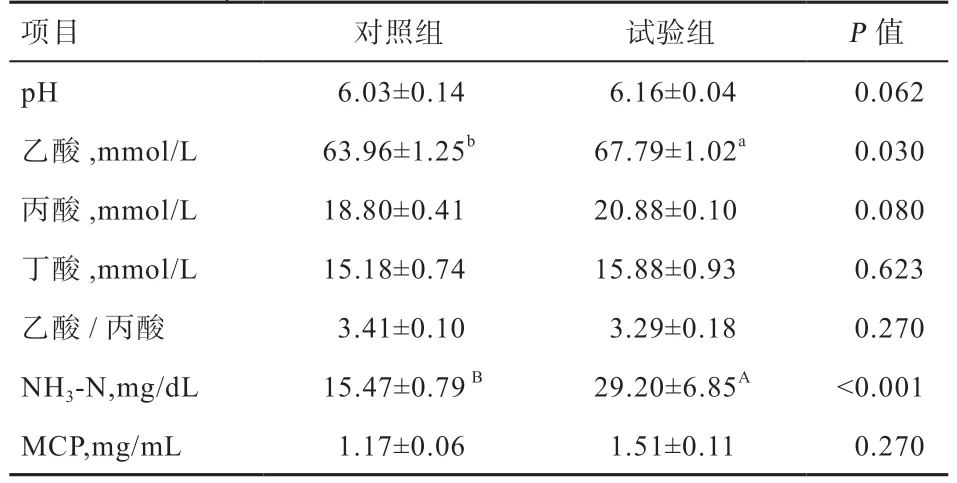
表2 γ-PGA 对小尾寒羊瘤胃发酵参数的影响
2.2 总DNA 质检结果 12 个瘤胃液样品中总DNA 浓度、纯度及质量均符合 Illumina Miseq 平台测序要求,可进一步用于后续试验。
2.3 16S rDNA 基因v3+v4 区域扩增结果 由图1 可知,2 组试验羊瘤胃液样品中的16S rDNA 基因V3+V4 区扩增条带大小正确,浓度适宜且条带单一,可用于后续测序试验。
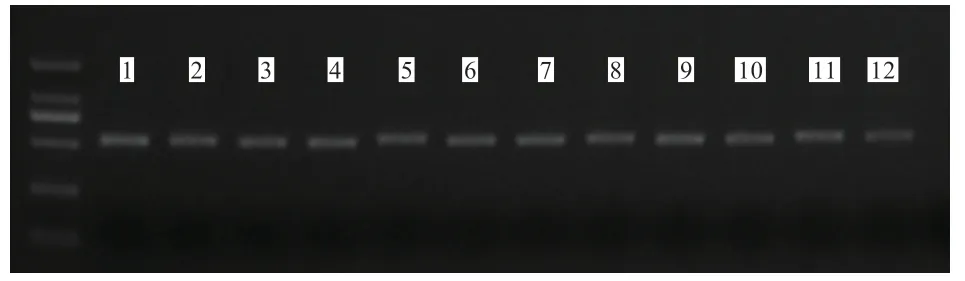
图1 试验样品16S rDNA 基因V3+V4 区域的扩增结果
2.4 操作分类单位(OTU)数量 通过Illumina Miseq测序结束后,OTU 划分样品的序列,并根据不同样品的聚类信息绘制韦恩图(Venn graph)。试验组样品获得97 个,对照组样品获得86 个,2 组间共享的OTU为596 个,饲粮添加γ-PGA 后小尾寒羊瘤胃菌群多样性升高。
2.5 OTU 稀释曲线和 OTU Shannon 稀释曲线 本试验通过获取的OTU 数据,利用随机抽取的序列数及OUT数目去建立曲线,得出每组样品的稀释曲线,用以表明样品测序出的数据量能否反映出瘤胃液中的物种多样性。由图3、4 可知,当测序深度较小时,OTU 数目发生剧烈变化,并随着测序深度的增加而增高。测序深度达到15 000reads 数时,稀释曲线趋于稳定,说明测序深度已覆盖到瘤胃液样品中所有的菌群物种中,测序结果可靠。测序深度达5 000 reads 时,Shannon 曲线趋于饱和状态,说明此测序量足够,可以进行样品菌群多样性分析。
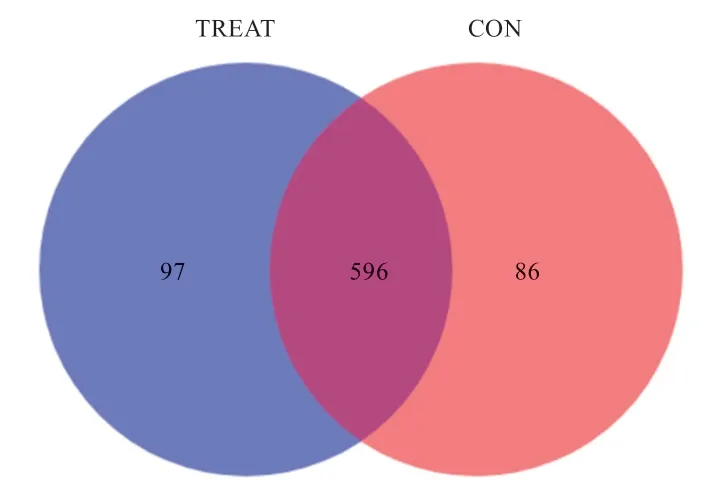
图2 小尾寒羊瘤胃液菌群韦恩图
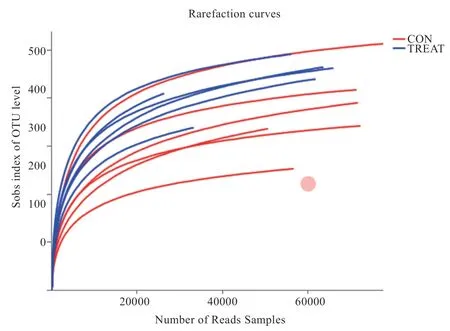
图3 小尾寒羊瘤胃细菌OTU 稀释曲线
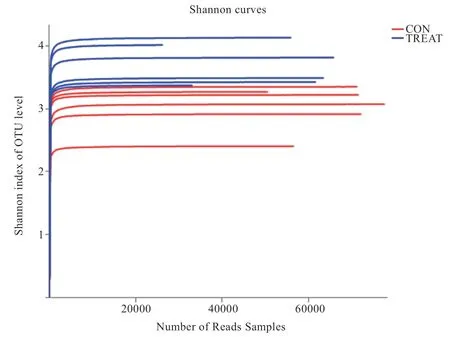
图4 小尾寒羊瘤胃细菌OTU Shannon 稀释曲线
2.6 OTU Alpha 多样性分析 基于OUT 的结果,计算出每组样品的Alpha 多样性。由表3 可知,试验组Chao-1 指数高于对照组,说明饲粮添加γ-PGA 使小尾寒羊瘤胃液中菌群丰富度升高,但差异不显著。试验组的Shannon 指数高于对照组(P<0.01),说明饲粮添加γ-PGA 能够提高小尾寒羊瘤胃菌群多样性。2 组覆盖率均大于99%,说明采集的样品足以反映瘤胃菌群情况。

表3 样品Alpha 多样性指数
2.7 瘤胃菌群结构分析
2.7.1 门水平 由表4、图5 可知,小尾寒羊瘤胃液中总共涉及17 个菌门,主要包括拟杆菌门(Bacteroidetes)、互养菌门(Synergistetes)、厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、螺旋菌门(Spirochaetae)、蓝藻门(Cyanobacteria)、疣微菌门(Verrucomicrobia)、丝状杆菌门(Fibrobacteres)及软壁菌门(Tenericutes)等,其中所含的主要细菌门即优势菌门为拟杆菌门、厚壁菌门及互养菌门,占总细菌门比例的90% 以上。试验组小尾寒羊瘤胃液中拟杆菌门的相对丰度高于对照组,厚壁菌门的相对丰度低于对照组,互养菌门的相对丰度高于对照组,但差异均不显著。与对照组相比,饲粮添加γ-PGA 可降低小尾寒羊瘤胃液中软壁菌门、螺旋菌门、未注释细菌门及蓝藻门的相对丰度(P<0.01),增加放线菌门的相对丰度(P<0.01)。
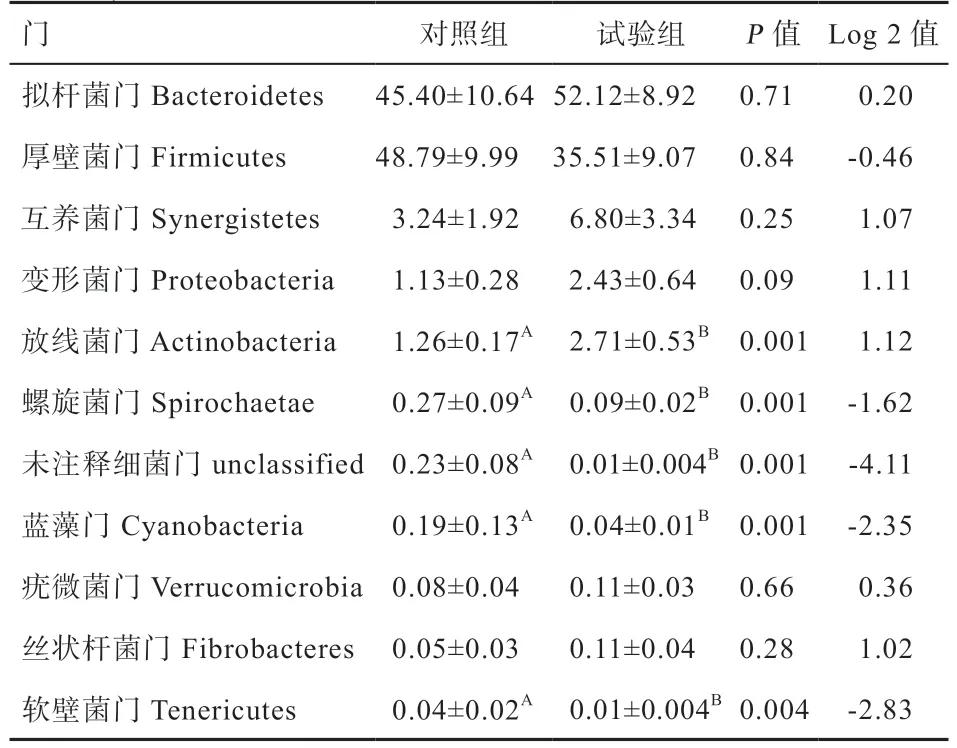
表4 γ-PGA 对小尾寒羊瘤胃菌群门水平上相对丰度的影响 %
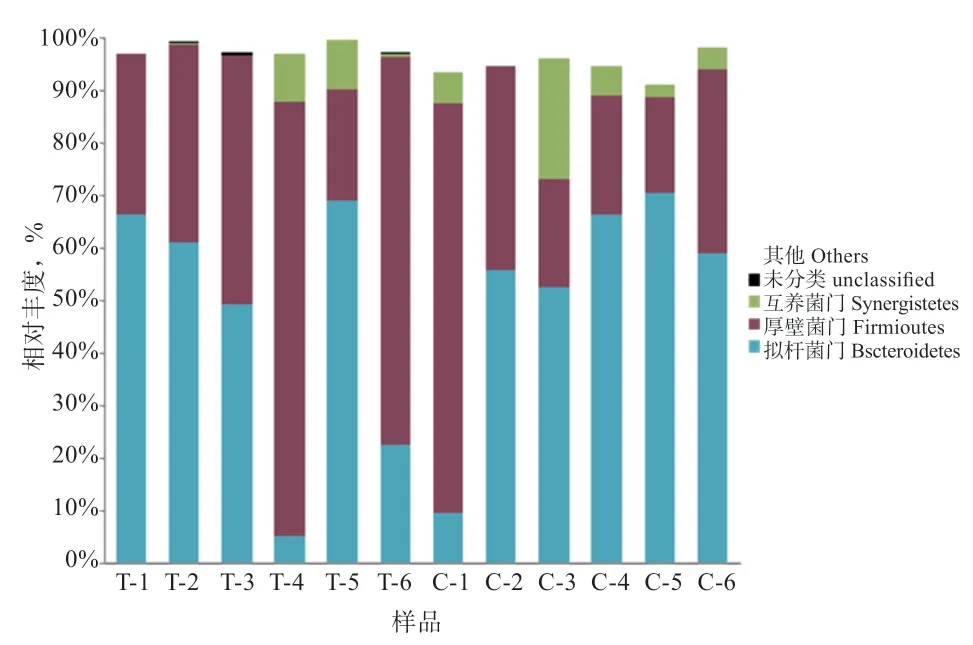
图5 门水平上瘤胃菌群分布图
2.7.2 属水平 在本试验中小尾寒羊瘤胃液菌群总共涉及188 个属种。由表5、图6 可知,相对丰度较高的29 个属种占各样品中总菌属的90% 以上,其中所包含的6 个菌属的相对丰度较高,分别是普雷沃氏菌属(Prevotella)、瘤胃球菌属(Ruminococcus)、奎因氏菌属(Quinella)、Fretibacterium、Prevotellaceae_UCG-003及解琥珀酸菌属(Succiniclasticum)。试验组普雷沃式菌属的相对丰度高于对照组,对照组瘤胃球菌属的相对丰度高于试验组,但差异均不显著。试验组小尾寒羊瘤胃液中解琥珀酸菌属(Succiniclasticum)的相对丰度高于对照组(P<0.01)。
2.7.3 组间差异显著性检验 由图7 可知,试验组小尾寒羊瘤胃液中解琥珀酸菌属的相对丰度高于对照组(P<0.01)。
3 讨 论
3.1γ-PGA 对小尾寒羊瘤胃发酵参数的影响 瘤胃pH是反映反刍动物瘤胃内环境的重要指标之一,其高低可直接影响瘤胃微生物的生长代谢[9],并可反映反刍动物瘤胃内的发酵及其有机酸的代谢状况。研究指出,瘤胃液pH 的正常范围是5.5~7.5,对其影响最大的主要是饲粮结构和营养水平[10]。本试验结果显示,2 组试验羊瘤胃液的pH 差异不显著,且均在正常范围内,说明饲喂γ-PGA 后小尾寒羊的瘤胃发酵和代谢均属正常。瘤胃微生物通过发酵饲料中的碳水化合物产生挥发性脂肪酸(VFA),并在此发酵过程中释放出的能量供微生物生长繁殖利用。VFA 是反刍动物主要的能量来源物之一,其合成受到饲粮结构、品质及瘤胃微生物活性等的调控,其中乙酸、丙酸和丁酸占总VFA 的95%以上[11]。本试验中,2 组饲粮经瘤胃微生物发酵后的主要产物为乙酸,并且试验组小尾寒羊瘤胃中的乙酸含量显著高于对照组,说明饲粮中添加γ-PGA 能够提高粗纤维的降解率,其可能是通过提高纤维分解菌(解琥珀酸菌属)的丰度所致。γ-PGA 分子链中含有大量的羧基[12],可能也是引起试验组乙酸升高的原因,有待进一步研究分析。2 组试验羊瘤胃丙酸和丁酸含量没有显著差异。研究指出,乙酸/丙酸越高,奶牛的氢浓度和产甲烷菌数越高,其拟杆菌门和厚壁菌门的丰度越低[13]。本试验发现,试验组瘤胃乙酸/丙酸低于对照组,这可能是饲喂γ-PGA 后小尾寒羊拟杆菌门和互养菌门的相对丰度有所提高的原因。
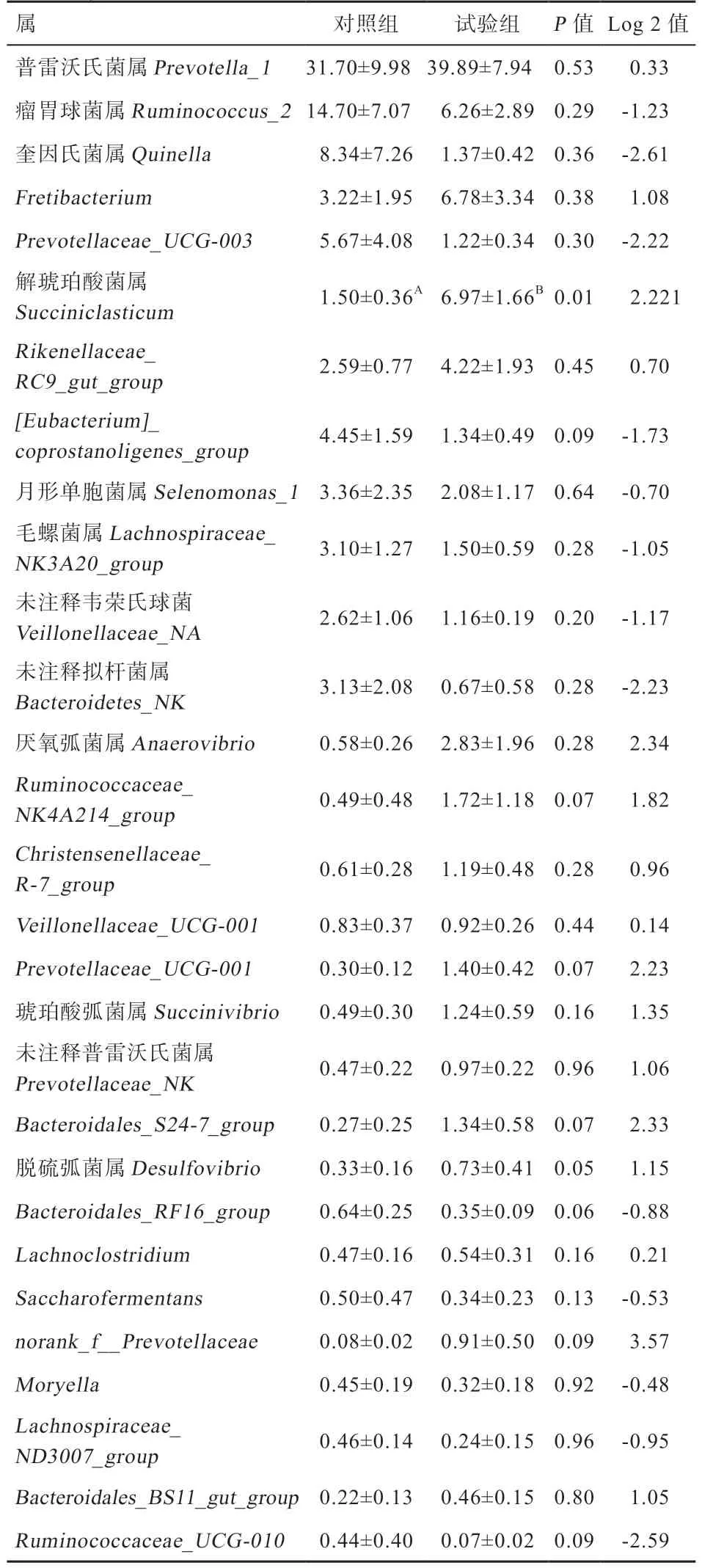
表5 γ-PGA 对小尾寒羊瘤胃菌群属水平上相对丰度的影响
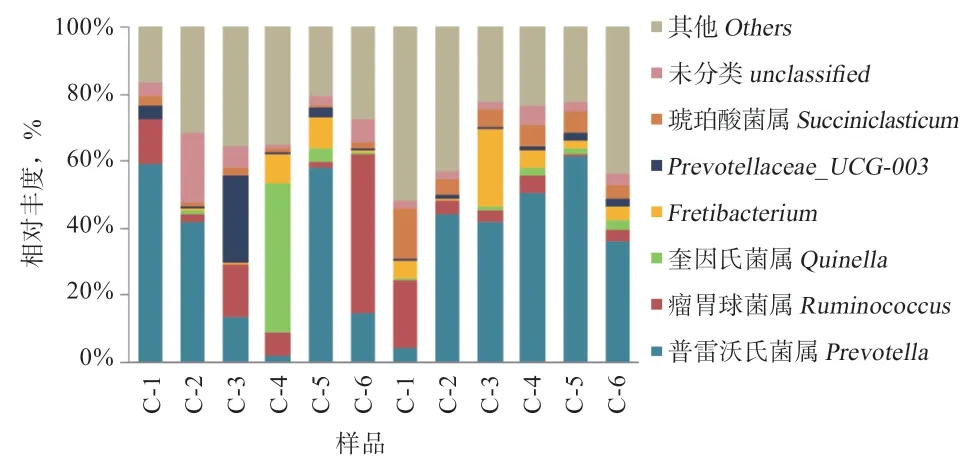
图6 属水平上瘤胃菌群分布图
反刍动物瘤胃中的NH3-N 主要来源于食糜中蛋白质的降解,是大部分微生物生长繁殖所需的原料,因此可用NH3-N 浓度反映瘤胃内微生物的活力[14]。研究表明,NH3-N 浓度在6~30 mg/dL 时,最适宜微生物的生长繁殖[15]。反刍动物瘤胃微生物经过复杂的发酵过程合成MCP,NH3-N 是瘤胃微生物合成MCP 的原料,过低及过高的NH3-N 浓度均不利于MCP 合成。研究指出,日粮添加谷氨酸渣可降低瘤胃NH3-N,提高MCP 含量[16]。本试验结果显示,试验组小尾寒羊瘤胃NH3-N 浓度极显著高于对照组,并且浓度在6~30 mg/dL,其可能是由于γ-PGA 特异性地增加了瘤胃内蛋白分解菌(如普雷沃式菌属)的数量而造成的。试验组MCP 含量略高于对照组,说明饲粮中添加γ-PGA 有利于小尾寒羊瘤胃MCP 合成。
3.2γ-PGA 对小尾寒羊瘤胃菌群结构的影响 瘤胃是反刍动物特有的消化器官,因含有丰富的微生物菌群而具有超强的降解纤维物质的能力,能将高纤维素物质转化为动物性蛋白。瘤胃微生物主要包括细菌、真菌和原虫,在消化饲料的过程中三者共同协作完成碳水化合物和蛋白质等的分解,并具有调控反刍动物的生产效率及健康状况等功能[17],细菌是瘤胃内种类和数目最多且最为复杂的微生物。反刍动物瘤胃内的细菌主要分布于拟杆菌门和硬壁菌门中,对瘤胃内容物的发酵过程具有重要的作用[18]。本试验结果表明,饲粮添加γ-PGA 可极显著提高小尾寒羊瘤胃菌群多样性。曾燕[19]研究指出,拟杆菌门和厚壁菌门是绵羊胃肠道中主要的优势菌门。本试验结果显示,饲粮中添加γ-PGA 后,在小尾寒羊瘤胃液菌群的门水平上,拟杆菌门和互养菌门的相对丰度分别提高了6.3%和3.6%。互养菌属中所属的各类菌种均具有降解氨基酸的能力。研究指出,从羊的反刍食物中分离出的互养菌属能以精氨酸和组氨酸作为底物,降解饲粮中存在的毒素,帮助宿主动物生存[20]。本试验中,饲粮中添加γ-PGA 可极显著降低小尾寒羊瘤胃液中软壁菌门、螺旋菌门、未注释细菌门及蓝藻门的相对丰度,极显著增加放线菌门的相对丰度;饲粮添加γ-PGA 后,小尾寒羊瘤胃液中普雷沃式菌属的相对丰度提高了8.2%。普雷沃式菌属为厌氧的革兰氏阴性菌,是瘤胃内重要的淀粉降解菌,还可以降解多种代谢产物如蛋白质、多糖及小肽等。
反刍动物的饲料成分主要以粗饲料为主,瘤胃中的纤维降解菌可以降解粗饲料中的纤维物质,将其转化为营养物质而满足宿主动物的能量需求[21]。产琥珀酸拟杆菌、黄色瘤胃球菌及白色瘤胃球菌是瘤胃内的主要纤维降解菌[22],其中解琥珀酸菌属中的产琥珀酸拟杆菌为厌氧型革兰氏阴性菌,具有降解黄白瘤胃球菌所不能降解的纤维素的能力[23],并且还具有较强的耐抗生素能力。本试验结果表明,饲粮添加γ-PGA 后极显著提高了小尾寒羊瘤胃液内解琥珀酸菌属的相对丰度。
4 结 论
本研究结果表明,饲粮中添加γ-PGA 可极显著提高小尾寒羊瘤胃乙酸、NH3-N 及菌群多样性;γ-PGA可在门水平上极显著降低小尾寒羊瘤胃液中软壁菌门、螺旋菌门、未注释细菌门及蓝藻门的相对丰度,极显著增加放线菌门的相对丰度,可提高拟杆菌门和互养菌门的相对丰度;γ-PGA 可在属水平上极显著提高小尾寒羊瘤胃内解琥珀酸菌属的相对丰度,也可提高普雷沃式菌属的相对丰度但差异不显著。综合分析,γ-PGA 具有优化小尾寒羊瘤胃微生态环境的作用,能改善瘤胃的发酵,提高饲料利用率。
