广州市农贸市场猪肉源伦敦沙门菌的流行情况及耐药性分析
2021-02-27王兰茜张丽娜张建民瞿孝云
高 远,王兰茜,张丽娜,符 颖,张建民,廖 明,瞿孝云
(华南农业大学兽医学院,人兽共患病防控制剂国家地方联合工程实验室,广东省动物源性人兽共患病预防与控制重点实验室,农业部人畜共患病重点实验室,广州 510642)
沙门菌(Salmonella)为革兰阴性杆菌,是一种重要的人兽共患致病菌,其分布广泛,种类繁多,不仅可以感染多种畜禽动物,甚至可以通过食品引起人类的食物中毒。每年感染沙门菌的病例超过9 300万例,死亡达15.5万人,其中,85%都与食物有关[1]。现今,沙门菌感染已成为国际上重要的公共卫生问题[2-5]。
迄今为止,沙门菌在世界范围内已经发现并鉴定出超过2 600多种血清型,绝大部分能导致人类和动物发病[6]。我国约有75%食源性疾病的暴发是由沙门菌引起的,其中,90%的与食物中毒和摄入污染畜禽肉及其相关制品有关[7]。广东省是畜牧生产和消费大省,猪肉也是沙门菌的主要贮库,畜禽产品的安全尤其是猪肉的安全直接关系到消费者健康[8]。伦敦沙门菌在20世纪40年代被发现,目前,成为沙门菌中排名前十的优势血清型,但在食品流通环节尤其是农贸市场中猪肉源菌株的研究鲜有报道。国内研究者曾在新生儿和食物中毒病人中有过报道,新加坡的研究在1名儿童体内检出多重耐药伦敦沙门菌[9-12]。因此,本研究旨在调查2016年5—10月广州农贸市场猪肉中伦敦沙门菌的流行与耐药情况,为临床中合理应用抗菌药物以及保障食品安全提供数据支撑。
1 材料与方法
1.1 主要材料
1.1.1 菌株来源 2016年5—10月,从广州市农贸市场采集分离伦敦沙门菌。药敏试验质控菌株为大肠埃希菌ATCC25922,购自中国兽医药品监察所。
1.1.2 主要试剂 木糖-赖氨酸-硫酸四癸钠琼脂(XLT4 Agar)、LB(Luria Bertani)营养琼脂、LB 肉汤购自广东环凯微生物科技有限公司;DNA Marker DL1000,DNA Marker DL2000,rTaqDNA聚合酶(Premix Taq)购自日本TaKaRa公司;Goldview核酸染料,购自美国Biotium公司。

1.2 方法
1.2.1 沙门菌的分离鉴定 将在农贸市场不同档口随机采集的猪肉样品装入无菌采样袋,在8 h内低温运送至实验室。将所有猪肉样品按每份25 g加入灭菌BPW 225 mL充分揉搓洗涤2 min后,置于37 ℃摇床中以100 r·min-1振荡6~8 h。取预增菌液体1 mL,转移至10 mL TTB内进行选择性增菌,温度为42 ℃,时间为18~24 h。再用接种环蘸取TTB增菌液划线接种于XLT4琼脂平板,37 ℃ 培养18~24 h。选取疑似沙门菌菌落(典型形态:色泽黑亮、周围有白色透明环)接种于沙门菌显色培养基上进行纯化,取纯化后的疑似菌落用沙门菌特异性基因invA进行聚合酶链式反应(PCR)鉴定,引物详见表2[13-14]。

表2 试验中所用引物的相关信息
1.2.2 血清学鉴定 按照泰国S&A公司提供的沙门菌血清诊断操作步骤对菌株进行血清型鉴定,查阅S&A公司沙门菌抗血清诊断附录,根据测定得到的抗原式确定沙门菌的血清型[12]。
1.2.3 药物敏感性试验 使用K-B纸片扩散法对伦敦沙门菌进行抗菌药物敏感性试验,并根据CLSI(美国临床实验室标准化委员会)推荐的操作规程对结果进行解释。使用大肠杆菌ATCC 25922作为质控菌株。药敏试验判定标准见表1。
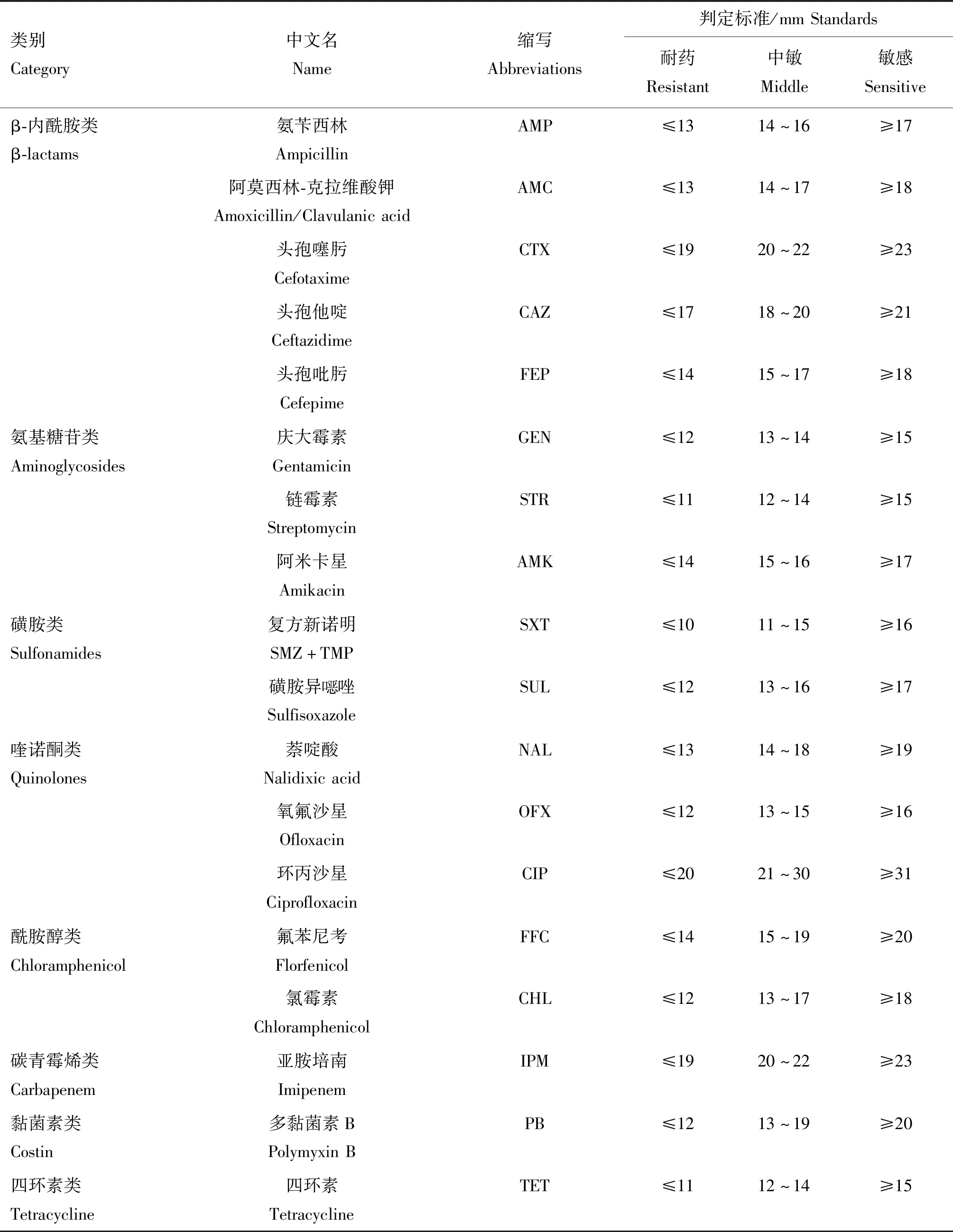
表1 药敏纸片种类及判定标准
1.2.4 耐药基因检测 利用水煮法提取28株伦敦沙门菌基因组DNA。选取如下基因:氨基糖苷类耐药基因aadA2、aadB、strA;酰胺醇类耐药基因aphAI-IAB、cat1b、cat2b、floR;磺胺类耐药基因sul1、sul2;四环素耐药基因tetA、tetB;选择β内酰胺类、碳青霉烯类耐药基因blaTEM、blaCTM、blaOXA、blaNDM;选择多黏菌素类耐药基因mcr-1;选择喹诺酮类耐药基因qnrA、qnrB、qnrC、qnrD、qnrS、oqxAB、aac(6’)-Ib-cr[15-17]。扩增后的产物进行琼脂糖凝胶电泳检测,结果为阳性的PCR产物直接送广州市艾基生物技术有限公司测序。测序结果通过NCBI网站(https://www.ncbi.nlm.nih.gov)进行Blast分析,与GenBank上对应基因序列进行比对。
2 结 果
2.1 分离鉴定及血清型鉴定
从广州市5个农贸市场采集到猪肉样品198份,经分离鉴定后得到147份沙门菌阳性样品,阳性率为74.2%。通过血清型鉴定,共鉴定出17种血清型,其中,主要为鼠伤寒(34/147,23.1%)、罗森(34/147,23.1%)、德比(31/147,21.1%)和伦敦(28/147,19.0%),详见表3。
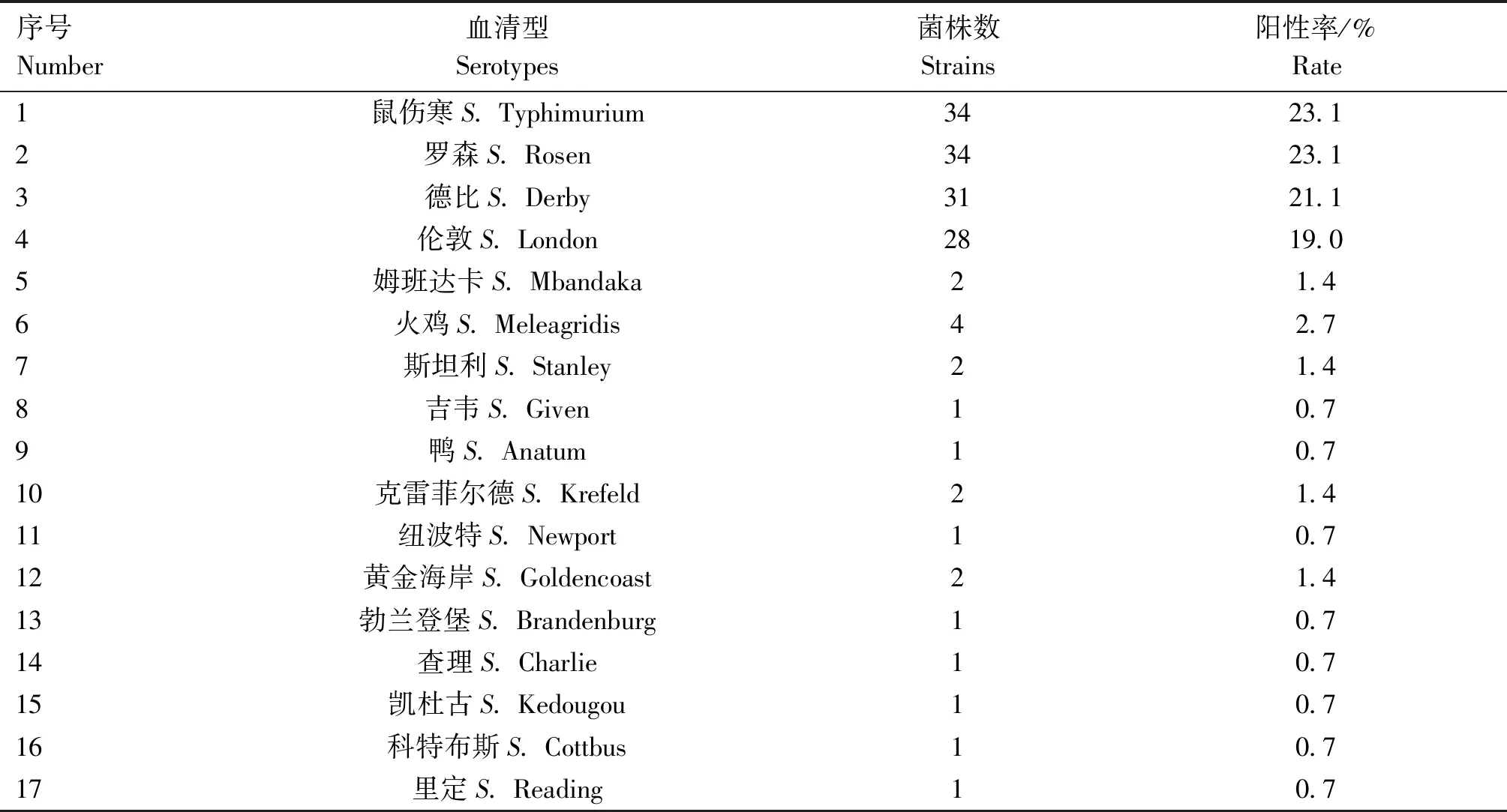
表3 沙门菌血清型鉴定结果
2.2 药敏试验

表4 28株伦敦沙门菌耐药情况
多重耐药率为71.4%(耐3类及其以上的抗菌药物)。28株伦敦沙门菌共存在9种耐药谱型,其中AMP+CN+S+SXT+SUL+FFC+C+TE为优势谱型(28.6%),其次是AMP+CN+S+SXT+SUL+TE(25.0%)。有2株菌对9种药物耐药,耐药谱型分别为AMP+CN+S+SXT+SUL+FFC+C+NA+TE(3.6%)和AMP+CN+S+SXT+SUL+FFC+C+PB+TE(3.6%),详见表5。

表5 28株伦敦沙门菌耐药谱
2.3 耐药基因检测
通过对28株伦敦沙门菌进行耐药基因检测发现,β-内酰胺类类耐药基因blaTEM的检出率为10.7%(3/28),未检出blaCTX-M、blaCTX-M耐药基因;氨基糖苷类耐药基因aadB的检出率为100.0%(28/28),未检出aadA1;酰胺醇类药物耐药基因cat1b、floR的检出率为39.2%(11/28),未检出cat2b;磺胺类耐药基因sul1、sul2的检出率均为60.7%(17/28);四环素类耐药基因tetA、tetB的检出率均为78.6%(22/28);喹诺酮类耐药基因qnrA检出率为3.6%(1/28),oqxAB检出率为10.7%(3/28),aac(6’)-Ib-cr检出率为7.1%(2/28);黏菌素类耐药基因mcr-1检出率为3.6%(1/28);未检出碳青霉烯类耐药基因blaNDM,详见表6。

表6 28株伦敦沙门菌耐药基因检出结果
3 讨 论
沙门菌可以通过食物链进入人体从而引起人食物中毒等症状。伦敦血清型不常见,1980年曾在匈牙利引起局部流行,2000年前后,韩国感染病例的增多趋势且出现产超广谱β内酰胺酶菌株[18-19]。国内关于伦敦沙门菌的报道主要集中在食物中毒的案例,特别是免疫功能不完善的新生儿和有慢性疾病的较易发病[20-21]。本研究发现在广州农贸市场售卖的猪肉中,伦敦沙门菌分离率高达19%,仅次于常见的鼠伤寒、罗森和德比血清型,为第四优势血清型。这提示广州市售猪肉中伦敦沙门菌污染较为严重,需及时对广州市农贸市场猪肉中伦敦沙门菌的流行与耐药情况进行检测与监测,这对广州地区的食品安全与公共卫生具有重要意义。
本研究在广州农贸市场中检出的高耐药率伦敦沙门菌的现象应引起重视。抗菌药物在畜禽养殖过程中的广泛使用及滥用,导致多重耐药菌株愈来愈多[28-29]。笔者发现伦敦沙门菌多重耐药率达到71.4%,其中优势耐药谱型为AMP-CN-S-SXT-SUL-FFC-C-TE,甚至有10株菌对5类以上的抗菌药物耐药,仅次于之前报道的多重耐药鼠伤寒沙门菌[30]。
在抗菌药物的作用压力下,食源性耐药沙门菌不断出现,细菌耐药机制复杂,但耐药表型与耐药基因之间往往存在着密切联系[31]。28株伦敦沙门菌氨基糖苷类耐药基因aadB检出率为100.0%,高携带率可能是导致沙门菌对庆大霉素耐药的主要原因。78.6%的菌株同时携带tetA基因和tetB耐药基因,tetA基因携带率小幅增加,tetB基因携带率明显增加,说明这两种耐药基因可使伦敦沙门菌对四环素耐药性加强[31]。OqxAB基因是编码多重耐药外排泵的常见基因,检出率为10.7%,aac(6’)-Ib-cr基因可以编码灭活酶,检出率为7.1%,qnr家族的基因qnrA、qnrB、qnrC、qnrD等能保护拓扑异构酶Ⅳ和促旋酶来降低药物敏感性,以上基因单独存在时可介导低水平的喹诺酮类药物耐药[32]。由于只有1株菌对传统药物萘啶酸耐药,所以喹诺酮类耐药基因检出率均较低。ESBLs基因是由质粒介导的水解酶,可以水解氨曲南等单环β-内酰胺类抗生素以及头孢他啶和头孢噻肟等第3代头孢菌素[33]。伦敦沙门菌对头孢类抗菌药物全部敏感,未检出blaCTX-M和blaOXA,而blaTEM检出率为10.7%,可能部分菌株携带该耐药基因导致其对氨苄西林产生了耐药现象。mcr-1作为介导的黏菌素耐药基因,很少在沙门菌中检出,本研究发现有1株沙门菌携带mcr-1且对多黏菌素耐药,其是否为质粒介导的耐药性亟待人们进一步研究[34]。以上结果提示,在动物饲养过程中,抗菌药物的不合理使用可能是导致细菌对氨基糖苷类、四环素等常见抗生素产生高度耐药的重要原因。喹诺酮类和头孢类药物对沙门菌有很好的抗菌作用,可作为临床用药的选择依据。
本研究可为畜禽养殖临床用药提供理论指导,为防控食品安全做好预警工作,加强市售猪肉的安全管控,对保障公共卫生安全及其人类健康具有重要意义。
4 结 论
