基于生物信息学方法的精神分裂症炎症机制研究
2021-02-23田海佳刘洪杨
田海佳,刘洪杨
精神分裂症是一种病因未明的重性精神障碍,全球约1%的人罹患此病[1]。该病起病隐匿,易复发,多呈慢性病程,患者的社会功能受损,给患者家庭以及社会都带了极大的负担。然而目前精神分裂症的诊断仍依靠症状学,缺乏客观生物学诊断标志,且病理生理机制仍未阐明,导致其治疗并未取得突破性进展。越来越多的研究表明免疫系统与炎症反应影响着大脑,且与情绪、认知以及行为的变化有关[2-3]。丹麦一项随访研究发现患有自身免疫性疾病的人群精神分裂症的发病风险增加29%,曾因感染而住院的人群精神分裂症的患病风险增加60%[4]。此外,Li等[5]发现精神分裂症患者携带的T细胞受体库与正常人有显著差异,且有暴力倾向与无暴力倾向者之间也存在差异。上述研究均提示免疫炎症或许参与了精神分裂症的病理生理机制。而Notter等[6]在动物实验中发现精神分裂症样行为与炎症有关,且有内皮细胞的参与;进一步的转化研究中也发现了精神分裂症与炎症之间的关联。
背外侧前额叶皮质(dorsolateral prefrontal cortex,DLPFC)参与人的认知、情绪等过程,且DLPFC的代谢紊乱与首发精神分裂症患者的认知功能缺损有关[7]。因此DLPFC常常成为精神分裂症的重点研究脑区。为进一步探索精神分裂症的炎症机制,本研究通过GEO数据库GSE12679基因芯片中精神分裂症患者和正常对照者DLPFC的内皮细胞转录数据进行分析,探讨炎症与精神分裂症的相关性。
1 资料和方法
1.1 资料
从美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)的GEO数据库获取GSE12679基因芯片数据,该基因芯片数据为人类尸检大脑DLPFC的内皮细胞和神经元的转录组测序结果,内皮细胞和神经元由通过激光捕获显微切割技术获得。样本测序由GPL570平台(Affymetrix公司的Human Genome U133 Plus 2.0芯片)进行分析。在该基因芯片数据中选取11例精神分裂症患者和5名健康对照的大脑DLPFC内皮细胞数据进行后续分析。
1.2 方法
1.2.1 临床资料收集 收集精神分裂症组与健康对照组样本的年龄、大脑组织pH值以及死亡时间。
1.2.2 差异表达基因 利用GEO在线工具GEO2R将对两组样本进行差异表达基因的筛选。以P<0.05以及|logFC|>1为标准筛选差异表达基因。
1.2.3 基因本体(gene ontology,GO)功能注释富集分析 生物学信息注释数据库 (database for annotation,visualization and integrated discovery,DAVID)是生物信息学领域使用广泛的免费分析软件,整合了生物学数据和分析工具。本研究采用DAVID的GO功能对差异表达基因的细胞成分、生物过程和分子功能3个方面进行分析。
1.2.4 京都基因与基因组百科全书(KEGG)通路富集分析 KEGG通路数据库主要包含涵盖通道在内的目前分子互动网络的信息,是生物信息学领域最常用的数据库之一。本研究采用DAVID数据库中的KEGG通路富集分析模块对差异表达基因进行通路富集分析。
1.2.5 统计学方法 利用SPSS 24.0统计软件对精神分裂症组与健康对照组的年龄、大脑组织pH值以及死亡事件进行独立样本t检验,P<0.05为差异具有统计学意义。
2 结果
2.1 样本临床资料比较
精神分裂症组年龄(42.7±7.8)岁;大脑组织pH(6.6±0.2);死亡时间(20.3±10.4)年。健康对照组年龄(43.2±10.6)岁;大脑组织pH(6.5±0.3);死亡时间(29.6±10.1)年。两组年龄、大脑组织与死亡时间差异无统计学意义(P均>0.05)。
2.2 差异表达基因筛选
经GEO在线工具GEO2R分析,从GSE12679基因芯片数据中找出差异表达基因389个,其中上调145个,下调244个,包括了大量涉及炎症反应的基因,如单核细胞趋化蛋白2(C-C motif chemokine ligand 8,CCL8)基因、CXC趋化因子配体(C-X-C motif chemokine ligand,CXCL)基因等。差异表达基因中前20个基因见表1。
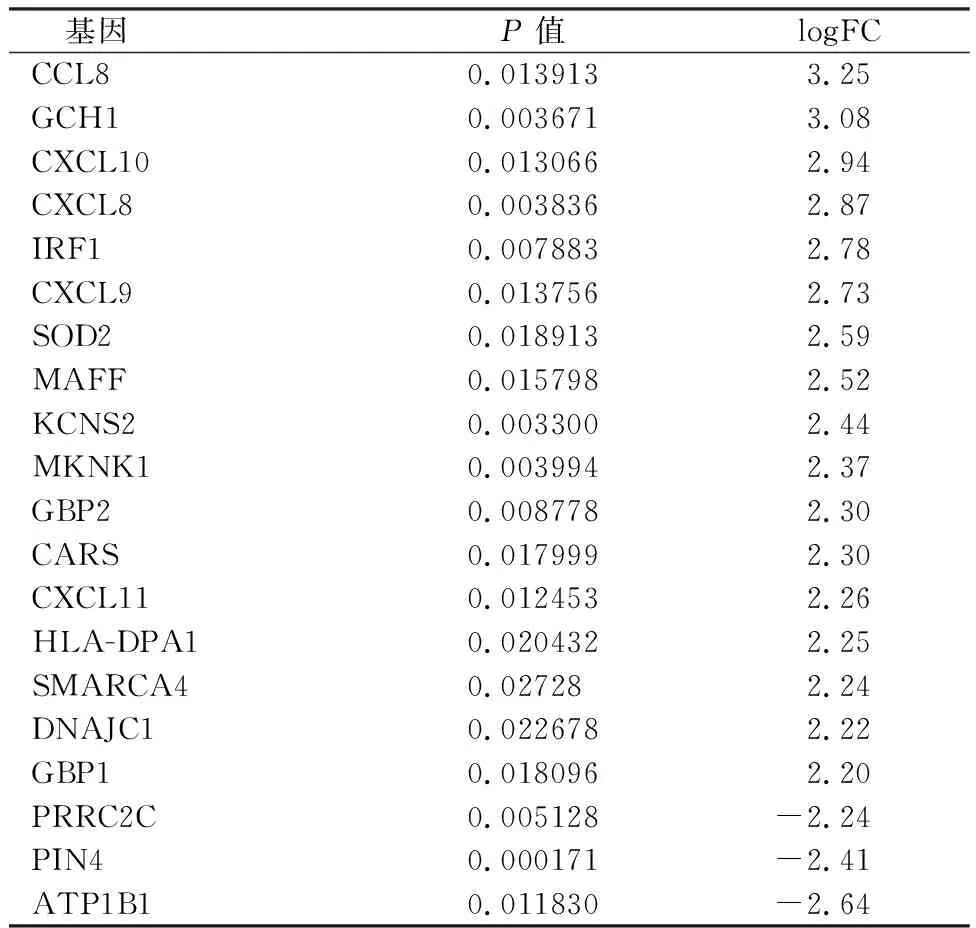
表1 前20个差异表达基因
2.3 差异表达基因GO功能富集分析
DAVID在线数据库对GO功能富集分析发现,差异表达基因富集于65个GO条目。对差异表达基因的注释分为细胞成分、生物过程和分子功能,对这3部分进行分析。结果显示,细胞成分中高尔基复合体、细胞质、细胞核、细胞膜等均有富集;在分子功能方面,蛋白结合、趋化因子活性、XCXR3趋化因子受体结合、RNA结合酶、GTP酶活性等均有富集;在生物过程方面,信号转导、免疫反应、干扰素介导的信号通路、对病毒的防御反应、趋化因子介导的信号通路、神经系统的发育等过程均有富集。见图1。
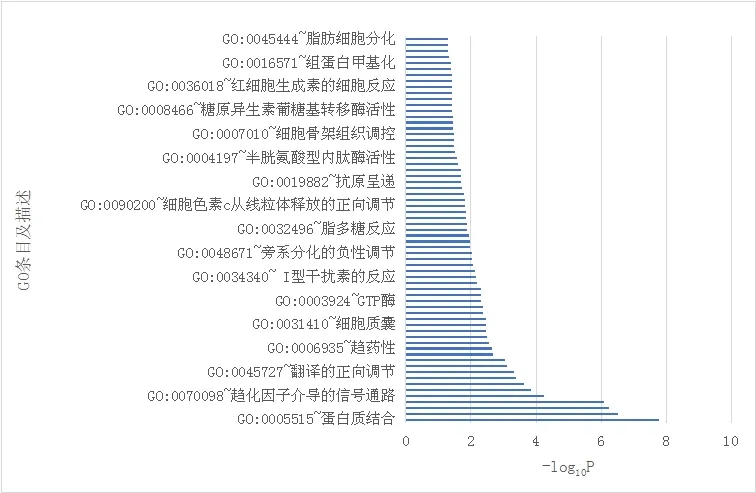
图1 差异表达基因GO功能富集分析
2.4 差异表达基因KEGG通路富集分析
通过DAVID在线数据库进行KEGG通路富集分析,结果发现差异表达基因主要富集于免疫、炎症相关等17条信号通路,包括甲型流感病毒A通路、肿瘤坏死因子信号通路、Toll样受体信号通路、RIG-I样受体信号通路、NOD样受体信号通路趋化因子信号通路等,暗示了炎症机制在精神分裂症中的作用。见图2。
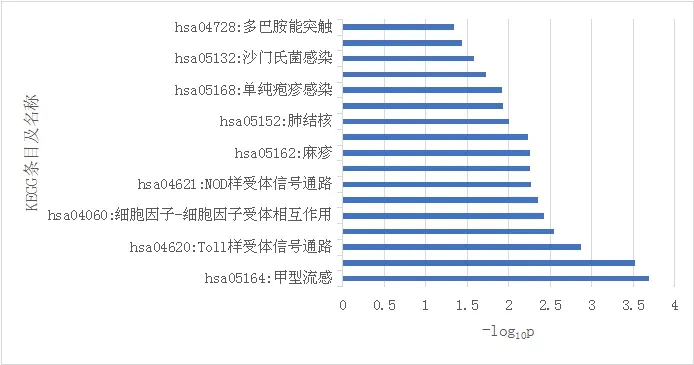
图2 差异表达基因KEGG通路富集分析
3 讨论
精神障碍研究难点之一在于难以获得活体脑组织,因此涉及大脑组织的研究常常为尸检研究。本研究数据来源于精神分裂症患者与正常人尸检脑组织DLPFC的内皮细胞。DLPFC 与精神分裂症的核心症状之一——认知功能下降有关,而不断有研究证明血管内皮紊乱相关的炎症反应与精神分裂症有关[8-9],因此认为DLPFC区域的内皮细胞为研究精神分裂症的炎症机制的较好来源之一。
本研究差异表达基因GO富集分析结果表明这些差异表达基因与炎症反应过程有关,例如与趋化因子、对病毒的反应以及干扰素-γ等有关;与既往研究一致[8]。趋化因子能诱导附近的细胞定向趋化,在炎症反应中起重要作用。干扰素-γ是一种促炎性细胞因子,其表达增高代表炎症水平的升高。本研究还发现内皮细胞的差异表达基因在涉及炎症反应KEGG通路中有所富集,包括趋化因子信号通路、细胞因子-细胞因子受体信号通路、肿瘤坏死因子信号通路、Toll样受体信号通路等。既往研究也表明趋化因子在精神分裂症中的作用,如Hong等[10]发现精神分裂症患者涉及神经免疫的趋化因子水平明显高于正常人。Martinuzzi等[11]发现具有更高水平趋化因子的精神分裂症患者症状更为严重,预后也更差。另外,荟萃分析发现细胞因子的改变与精神分裂症的认知功能缺损有关[12],但仍需进一步研究明确不同细胞因子在精神分裂症中的作用。此外,肿瘤坏死因子、Toll样受体与精神分裂症的关系在研究中均有报道[13],这些研究均提示精神分裂症生物学过程涉及炎症机制。有学者总结神经炎症是精神分裂症发生发展的重要机制之一,抗炎治疗能帮助缓解精神分裂症的症状[14],本研究结果亦证实了这一理论。
综上,结合已有精神分裂症的炎症机制相关研究,本研究进一步证实了精神分裂症与炎症之间的关系;提示炎症相关的指标或许可以成为辅助精神分裂症的诊断、分类以及预后判断的生物学标志。然而本研究也具有一定的局限性,首先,该样本来源为尸检,对精神分裂症患者生前用药情况以及症状严重程度不详,因此研究结果可能无法排除药物的干扰,精神分裂症组组内异质性可能较大。其次,样本量较小,相对而言统计学效能不够,还需进一步扩大样本量进行相关研究分析。
