围栏封育对高寒草甸生物量和群落结构的影响
2021-02-23桑丁松毛
桑丁松毛
(青海省玉树市林草综合服务中心,青海 玉树 815099)
草地施肥、围栏封育是恢复退化高寒草甸的主要措施[1]。草地施肥能在短时间内快速改善土壤表层肥力以及草地植物群落结构,有助于草地生产力的恢复[2]。但也有研究指出,草地施肥会减少群落物种数量,降低物种多样性[3]。相比于草地施肥,围栏封育在增加地上生物量的同时,还能显著改善土壤理化性质和生物活性,增加土壤种子库多样性[4]。本文以青海玉树州长期固定高寒草甸监测样地为研究对象,分析了多年围栏封育对其生物量和群落结构的影响,以期为退化的高寒草甸研究提供数据参考。
1 材料与方法
1.1 试验地概况
试验地位于玉树州囊谦县(30.79°N、97.20°E),选取10 m×10 m 的固定监测样地,自2009 年开始对监测样地进行围栏封育。试验地为冬季牧场,土壤类型为高寒草甸土,植被类型主要是以莎草科高山嵩草、禾本科细柄茅和蓼科圆穗蓼等为主。主要伴生植物为火绒草、委陵菜、垂穗披碱草及高山唐松草等,并伴有其他杂草。
1.2 试验方法
在固定样地随机选取10 个1 m×1 m 的小样方,2009—2019年连续11年对采取的样方进行观测,并对1 m2样方的植被盖度、群落平均株高、物种数及毒杂草物种数等形态指标进行测定。对1 m2高寒草甸样方的生物量指标进行测定,牧草鲜重为剪去植物地上部后的称重;可食牧草鲜重为莎草科牧草等优势牧草的重量;将地上生物量分别称取鲜重后,在65 ℃恒温箱内烘干至恒重,分别计算牧草干重和可食牧草干重;可食牧草比例为可食牧草干重与牧草干重之比。
1.3 数据处理
采用Excel 2010 对数据进行处理,利用SPSS 17.0进行方差分析和相关性分析。
2 结果与分析
2.1 围栏封育对高寒草甸形态指标的影响
由表1 可见,围栏封育下,高寒草甸形态指标呈现不同的变化。其中,植被盖度在11年内的变化范围为72.17%~89.44%,在2015 年达到最大值,为89.44%,而在2011 年最小,为72.17%;群落平均株高和物种数均在2013 年达到最大值,分别为15.89 cm 和17.44 种,这2 个指标较2009 年围栏封育开始时均明显提高(P<0.05),分别增加了117.67%和66.10%,2 个指标分别在2012、2011 年达到最小值,分别为6.50 cm 和8.33 种;毒杂草物种数自2009 年围栏封育开始后呈不断减少的趋势,2019 年达到最小值,为1.40 种。

表1 围栏封育后高寒草甸形态指标变化情况
2.2 围栏封育对高寒草甸生物量的影响
由表2 可见,在2009—2019 年围栏封育期间,高寒草甸样方各个生物量均表现出先增后降的趋势。在2013年,牧草鲜重、可食牧草鲜重、牧草干重和可食牧草干重均达到最大值,分别为708.33、655.56、360.96 和309.79 g,与2009 年相比均显著增加(P<0.05),分别增加了2.07 倍、3.53 倍、1.41 倍和2.49 倍;之后,随着围栏封育年限的增加,这4 个生物量均逐渐减少。随着围栏封育年限增加,可食牧草比例大幅增加,在2017年达到最大值,为89.74%,与2009年相比增加了51.00%。
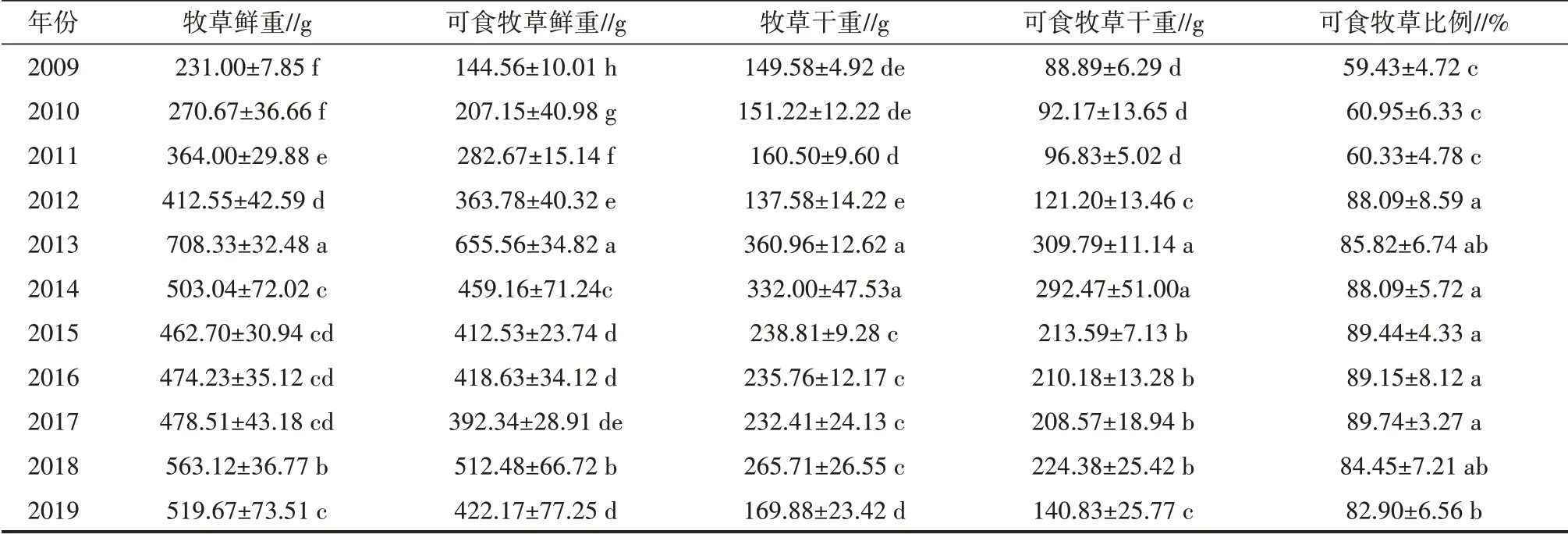
表2 围栏封育后高寒草甸生物量变化情况
2.3 围栏封育下高寒草甸各指标相关性分析
由表3 可知,围栏封育年限与物种数呈极显著正相关(P<0.01),与牧草鲜重显著负相关(P<0.05)。物种数与牧草鲜重呈显著正相关(P<0.05),与可食牧草鲜重、牧草干重及可食牧草干重均呈极显著正相关(P<0.01)。
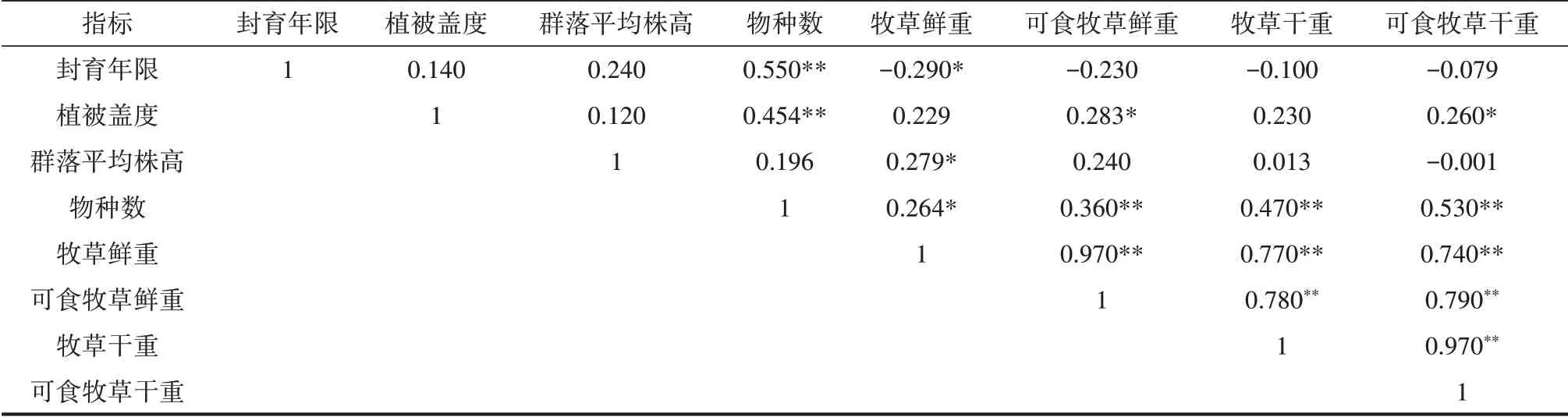
表3 高寒草甸各指标之间的相关性
3 小结与讨论
围栏封育是利于退化高寒草甸恢复的有效措施,其可以有效减少动物的啃食踩踏,促进优势牧草生长[5]。研究表明,围栏封育能显著增加高寒草甸生物量和物种组成[6,7]。本研究也同样发现,随着围栏封育年限增加,高寒草甸样地上生物量、可食牧草鲜重及物种数都明显增加;此外,相关性分析也表明封育年限与物种数呈极显著正相关。
封育年限对高寒草甸生物量及物种数的影响很大。王兰英等[8]研究发现,围封降低了高寒草甸物种数。陈智勇等[9]在西藏退化高寒草甸围栏封育研究中发现,退化草地生物多样性及生物量均随着围封年限的增加呈现先增后减的现象,这与本研究结果一致。
目前,关于高寒草甸围封年限拐点并无一致结论。本研究的拐点出现在封育第5 年,而陈智勇等[9]研究发现第7 年时出现拐点,也有研究指出,在封育第4年为高寒草甸生物量变化拐点[10]。但不论是短期还是长期围栏封育,其对高寒草甸土壤微生物和土壤结构等多方面确实存在重要影响[11],能够促进退化高寒草甸恢复。要进一步探究围封对高寒草甸的影响,还需要通过长期动态监测围栏封育高寒草甸的生物量及土壤理化性状的变化。
