湿地松引种家系的个体育种值评价*
2021-02-11谭志强邓乐平李义良陈秀云李福明郭文冰赵奋成
谭志强 邓乐平 李义良 陈秀云李福明 郭文冰 赵奋成 王 哲
(1. 台山市红岭种子园,广东 江门 529223;2. 广东省森林培育与保护利用重点实验室/广东省林业科学研究院 ,广东 广州510520)
湿地松Pinus elliottii原产美国东南部,因其生长迅速、适应性强,是人工造林的理想树种。我国最早引种湿地松,可追溯到上世纪30 年代初,由旅美华侨肖德钦先生带回一批湿地松种子[1]。经过几十年的引种试验和示范,证明湿地松是很有推广价值的速生用材、松脂树种[2-4]。从1973 年开始,陆续大量进口湿地松种子,在我国广东、广西、湖南、湖北、江西、安徽等13 省(区)广泛种植,成为我国亚热带地区一个主要的造林树种[5-7]。为了获得品质更好的湿地松种子,广东、湖南、浙江、福建等省建立了湿地松种子园[8-10]。广东省台山市红岭种子园是国家级林木良种繁育中心、全国特色种苗基地,经过几代人努力,建起国内的第一个湿地松初级种子园、一代种子园、改良种子园、精选种子园和二代种子园[11-12]。然而,湿地松作为外引树种,建园材料大部分来自三四十年代引种的湿地松林分,遗传背景窄,解决这一问题的办法是从原产地继续引进新的基因资源。种子园于1995 年从美国引进湿地松全同胞家系、半同胞家系种子,开展试验与测定工作,现对其进行个体育种值评价,筛选优良个体拓宽现有种子园遗传基础。
1 材料与方法
1.1 试验材料概况
试验材料取自广东省台山市红岭种子园试验区内的湿地松种质资源林,地理位置为22°11′N,112°49′E;属南亚热带海洋性气候,年均温21.8 ℃,年积温7 654 ℃,无霜期360 天,年降雨量1 940 mm;平均海拔30 m,土壤为花岗岩发育的酸性砖红壤,pH 值5.0~5.5,土层深厚。
该湿地松种质资源林于1997 年6 月种植,种子来源为美国,共126 个全同胞家系、32 个自由授粉家系。试验设计为随机完全区组设计,1~4株单行小区,15 个区组,每个家系的设计总株数40 株。试验林地势平缓,前茬为湿地松人工林。造林前除杂、炼山、挖穴,穴规格为40 cm×40 cm×30 cm ,株行距3 m×3 m。试验林四周种植2 行湿地松作为保护行。同期还种植了改良代种子园混合种子、湿地松一代种子园10 个湿地松混合家系的林分作为生长量对比。
1.2 数据获取与统计分析
2014 年11 月,对种质资源林进行每木检尺,树高H测量采用超声波测高测距仪(Haglöf Vertex IV,瑞典),1.3 m 处胸径DBH 采用测围尺,利用形数公式换算单株材积Vol。
利用统计软件Asreml-R 3.0 的个体模型对亲本、个体的育种值作分析[13],半同胞家系与全同胞家系个体模型采用的线性模型分别为:

2 结果与分析
2.1 湿地松种质资源总体表现
对早期引种的158 个湿地松家系、改良的混合种子、湿地松一代种子园10 个湿地松混合家系生长量进行了比较,结果显示广东改良的家系在树高、胸径、材积均值上高于引种家系,引种家系与一代种子园种子的均值相似。引种家系的最大值高于改良家系与一代混合种子,最小值小于改良家系与一代混合种子,说明引种的家系中变异丰富,并且存在优良的种质,可通过选优,作为新一轮育种材料的增补。
使用Asreml-R 软件的个体模型对引种湿地松半同胞家系和全同胞家系进行遗传参数估算时,全同胞家系3 个性状的家系效应均不显著,半同胞家系中区组效应不显著,最终模型的各项效应估算值如表2 所示。

表1 3 个湿地松子代群体的生长表现Table 1Growth performance of three progeny populations of Pinus elliottii
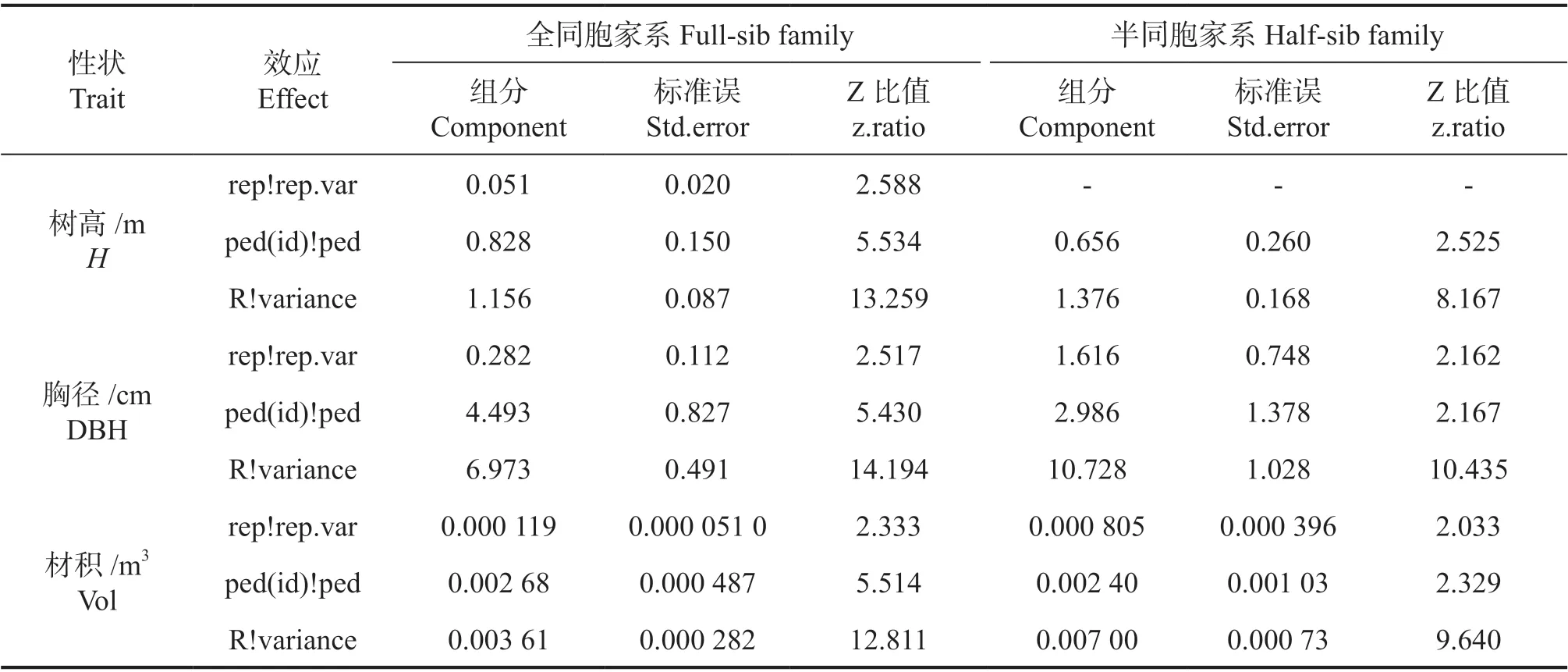
表2 个体模型中各项效应的估算值Table 2 Estimated value of each effect in the individual model
根据模型各项效应的估算值,计算半同胞家系树高、胸径、材积的单株狭义遗传力分别为0.323、0.218、0.255;全同胞家系树高、胸径、材积的单株狭义遗传力分别为0.417、0.392、0.426(表3)。在此基础上进一步计算遗传变异系数,成林树高趋于稳定,树高的变异较小,这也体现在两种家系的树高性状遗传变异系数(6%,表3),材积的遗传变异较丰富,因此将材积性状作为后续育种值排名的依据更可靠。对育种值计算的精确度估算结果显示,半同胞家系个体育种值精确度范围较大,为0.4~0.9,主要集中在0.5~0.6,而全同胞家系个体育种值精确度范围较集中,均在0.8~0.9(图1)。

图1 湿地松引种家系个体育种值计算精确度分布Figure 1 Distribution map of calculation accuracy of individual breeding value of Pinus elliottii introduced families
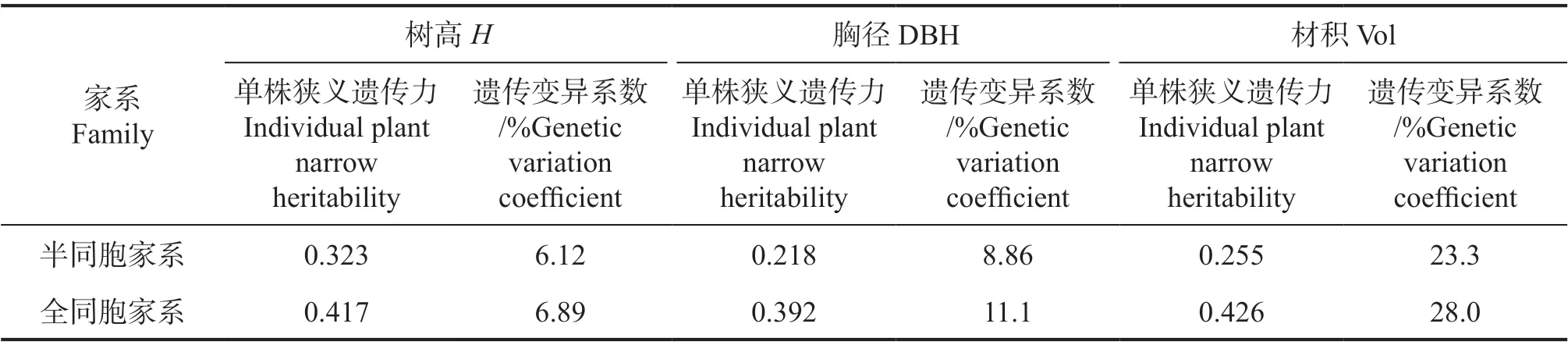
表3 湿地松引种家系遗传参数估算Table 3 Estimation of genetic parameters of Pinus elliottii Introduced families
个体模型可同时评价亲本与子代的育种值,126 个全同胞家系中,有7 个家系材积性状的个体育种值排名在前30 位,个体育种值高的个体集中在家系28、52、63 中(表4)。前30 名优良个体来源的家系中,28、52、63、96、107 这5 个家系的材积性状在家系育种值排名中也在前30 个内(表5),但家系13 与48 的家系育种值表现一般,这说明家系整体表现一般的,也存在可作为优良亲本的个体。考虑到遗传多样性,从前30 名的个体中选择了7 个个体进入新一轮育种群体。

表4 湿地松全同胞家系材积性状育种值排名前30 的个体Table 4 Top 30 individuals with breeding value for volume trait of full-sib Pinus elliottii families

表5 湿地松全同胞家系材积性状育种值排名前30 的家系Table 5 The top 30 Pinus elliottii full-sib families with the breeding value of volume trait
半同胞家系的表现与全同胞相似,排名前15名的个体集中在127、136、140、141 和144 上(表6),从每个家系选择1~2 个个体,共7 个优良个体,入选新一轮育种群体。对其它半同胞、全同胞家系个体,采用生长量育种值排名靠前、综合性状表现优良的原则进行优树选择。

表6 湿地松半同胞家系材积生长量育种值排名前15 的个体Table 6 Top 15 individuals with breeding value for volume trait of half-sib Pinus elliottii families
3 结论与讨论
作为引种树种,遗传背景越来越窄是必须面对的难题,需要通过不断从原产地引进新的种质资源拓宽遗传背景。从生长数据上来看,一代种子园混合种子的生长量与引种家系相当,改良代家系的生长量明显高于引种家系和一代种子园,邓乐平等[12]通过比较改良种子园与一代种子园子代4、13、18 年生时的单株材积,显示增益达到15.08%~25.66%。引种家系优良个体的生长量大于选育的家系,且最差的个体生长量也低于改良代家系和一代种子园,表明引种家系包含更多的遗传变异,优良种质可以在一定程度拓宽现有种子园的遗传基础。
林木中大多生长量性状是数量性状,因此大部分林木生长量性状的狭义遗传力为0.1~0.3[14]。Dorman 等[15]总结湿地松树高、胸径、材积狭义遗传力范围分别为0.03~0.37、0.22~0.58、0.16~0.35。赵奋成等[16]对种子园内60 个湿地松自由授粉家系进行了多年持续观察,其树高、胸径、材积单株狭义遗传力范围分别为0.27~0.46、0.21~0.38、0.25~0.4,本研究结果显示半同胞家系和全同胞家系的三个性状的单株遗传力均在此范围内,其中全同胞家系材积的单株遗传力相对最高(0.426),这可能与试验地只有一个地点有关。总体上来说,湿地松生长量性状受中等程度的遗传控制,经过选择可获得一定的遗传增益。
在Asreml 里,使用个体模型计算时,输入了子代对应的谱系,通过MME 方程的求解,可同时获得子代个体和母本的育种值。通过对子代个体育种值的排名情况,可以直接筛选优良单株[13]。项目引进湿地松的材料是种子,没有其亲本的材料,所以并没有计算亲本的育种值,只计算子代的育种值并排名。由于全同胞家系拥有更详细的谱系信息,计算出来的育种值精确度要高于半同胞家系。本研究以材积育种值为排名的标准,对应个体的胸径性状育种值排名也靠前,树高性状部分个体的育种值排名却相对不高,这可能与两个原因有关,一是树高对材积的影响只占小部分;二是树高测量的误差相对更大。随着分子标记技术的广泛应用,本研究下一步将利用分子标记确定入选个体的谱系关系、遗传距离,为拓宽湿地松种子园遗传基础提供科学依据。引进的湿地松种质宝贵的基因资源,目前只利用其生长性状的优良个体,在将来会对其他例如产脂量、抗性等有价值的性状进行评价,其丰富变异等待进一步挖掘利用。
