银翘散及其拆方对流感病毒感染自然杀伤细胞活性的影响及转录组的比较分析
2021-02-03陈能斌周德润王湛钧
陈 蓓,马 荣,陈能斌,周德润,王湛钧
1.新疆医科大学第一附属医院 药学部,新疆 乌鲁木齐 830011
2.上海中医药大学附属岳阳中西医结合医院,中西医结合临床研究所,上海 200437
3.上海市中医药研究院,中西医结合临床研究所,上海 200437
4.华东理工大学生物工程学院,上海 200237
银翘散出自清代吴塘所著的《温病条辨》,为辛凉解表的代表方剂,其主要功用为辛凉透表、清热解毒,是临床上治疗流行性感冒的常用中医药方剂。银翘散原方由连翘(30 g)、金银花(30 g)、桔梗(18 g)、薄荷(18 g)、淡竹叶(12 g)、荆芥穗(12 g)、淡豆豉(15 g)、牛蒡子(18 g)、生甘草(15 g)为散,以鲜芦苇根煎汤服用。现代药理学研究证明,银翘散能够降低流感病毒感染小鼠死亡率及肺部病毒载量、延长小鼠生存周期[1-2]。在胸腺缺陷小鼠 [T 细胞缺陷,自然杀伤(natural killer,NK)细胞正常] 模型中,NK 细胞活性在流感病毒感染后第1 天显著升高,第3、5、7 天明显降低;银翘散各剂量组NK 细胞活性在流感病毒感染后第3、5、7 天较模型组有明显提高[3]。本课题前期研究表明,在SCID 小鼠(T 和B 淋巴细胞缺陷,NK细胞活性较正常小鼠高)模型中,NK 细胞活性在流感病毒感染第3 天开始降低;经银翘散治疗后,NK 细胞活性有所提高,并且各剂量组间体现出一定的量效关系[4],同时银翘散还能够增强抗病毒免疫应答相关细胞及因子水平[5-6]。
对于不同的免疫状态,流感病毒均能使NK 细胞活性呈现降低趋势,而银翘散能够翻转这种免疫状态,增强NK 细胞的免疫功能。为此,本实验进一步将流感病毒感染的NK 细胞作为研究模型,通过君、臣、佐、使的拆方研究,明确银翘散发挥抗病毒及免疫增强作用的主要药物,为银翘散在临床中的精简应用提供实验依据,为中药抗流感病毒方剂的现代化研究提供研究思路。
1 材料
1.1 药品与试剂
金银花(批号 1410001W)、连翘(批号1409002W)、薄荷(批号1408001S)、牛蒡子(批号1411002W)、淡豆豉(批号1410001W)、荆芥穗(批号1408001S)、桔梗(批号1410002S)、芦苇根(批号1407001W)、淡竹叶(批号14109001S)和生甘草(批号1411003S),均为配方颗粒,购自华润三九医药股份有限公司。本实验中使用的药物剂量均参照方剂中的临床剂量,将生药剂量折算为颗粒剂的用量,并将临床剂量按照体表面积等效剂量转换法,换算为小鼠剂量,并按照临床配伍比例配伍使用。故本研究中各药物的小鼠剂量为金银花0.44 g/kg、连翘0.22 g/kg、薄荷0.22 g/kg、牛蒡子0.132 g/kg、淡豆豉0.11 g/kg、荆芥穗0.088 g/kg、桔梗0.264 g/kg、芦苇根0.11 g/kg、淡竹叶0.088 g/kg、生甘草0.367 g/kg。各拆方组中药味的剂量与其在全方中的剂量相同。银翘散和各拆方分别按照方剂配伍比例将各药物配方颗粒混合后,均以高纯水溶解,用于小鼠ig 给药。将银翘散和臣药中使用的配方颗粒分别按照方剂配伍比例混合后,溶于高纯水,0.22 μm 微孔滤膜滤过后作为细胞实验用储备液。利巴韦林颗粒(批号140920),购自四川百利药业有限责任公司。FITC-Ly-49D、PE-Ly-49H、FITC-CD3e、PE-CD49b 和Alexa Fluor 647-NKp46抗体,均购自BD Pharmingen 公司。红细胞裂解液和4%多聚甲醛溶液,购自北京索莱宝生物科技有限公司。
1.2 实验动物
4 周龄SPF 级ICR 和C57BL/6J 小鼠,雌性,体质量13~15 g,购自北京华阜康生物科技股份有限公司,实验动物许可证号SCXK(京)2014-0004。动物实验按照新疆医科大学第一附属医院动物伦理委员会相关规定进行,动物实验伦理批准号为IACUC-20140214009。小鼠饲养于生物安全实验室(ABSL-2)动物室,室内温度26 ℃,湿度(60±5)%,且12 h 明暗交替,正常饮食、饮水。小鼠给药体积为0.2 mL/10 g。
1.3 病毒
小鼠流感病毒A/Puerto Rico/8/1934(PR8,H1N1 型),由中国中医科学院中药研究所生物安全实验室提供,经鸡胚传代使用。将鸡胚中获得的病毒原液用生理盐水稀释600 倍,作为工作液,用于小鼠感染实验。
人流感病毒A/Hong Kong/8/68(HK8,H3N2型)购自美国ATCC(VR-1679™),经犬上皮细胞MDCK 传代后保存为病毒原液。
1.4 细胞株
人自然杀伤细胞NK-92MI,购自美国ATCC(CRL-2408TM)。细胞以α-MEM(无核糖核苷和脱氧核糖核苷)培养基培养,培养基中添加0.2 mmol/L肌醇、0.1 mmol/L β-巯基乙醇和0.02 mmol/L 叶酸,含12.5%马血清和12.5%胎牛血清。细胞置于37 ℃二氧化碳孵箱中培养。银翘散和臣药对该细胞的剂量分别为银翘散50 μg/mL,臣药100 μg/mL。剂量经细胞活性实验(CCK-8 法)检测获得,为药物在NK-92MI 细胞中的最高无毒剂量。
1.5 仪器
FACS Calibur 流式细胞仪,美国Beckman-Coulter 公司。EG1160 包埋机,德国Leica 公司;H93/RM2126 切片机,德国Leica 公司;ECLIPSE LV100POL/50IPOL 光学显微镜,日本尼康公司。
2 方法
2.1 实验分组、小鼠肺炎模型的制备及给药
将ICR 和C57BL/6J 小鼠分别随机分为对照组、模型(PR8 病毒感染)组、银翘散组(3.09 g/kg)、君药(金银花、连翘)组(1.00 g/kg)、臣药(薄荷、牛蒡子、淡豆豉、荆芥穗)组(0.83 g/kg)、佐药(桔梗、芦苇根、淡竹叶)组(0.70 g/kg)和使药(生甘草)组(0.56 g/kg)、阳性药物(利巴韦林)组(82.5 mg/kg)。ICR 小鼠每组10 只,C57BL/6J 小鼠每组3 只。将小鼠乙醚浅麻醉后,用PR8 病毒工作液滴鼻感染小鼠,30 μL/只,对照组小鼠滴鼻感染等体积的生理盐水。PR8 病毒感染后1 h 按照组别分别ig 给药。
2.2 小鼠肺质量抑制率的检测
各组ICR 小鼠给药后第5 天,称体质量,ip 氯胺酮(100 mg/kg)麻醉后45 min,脱颈椎处死,摘取肺脏并称质量。计算肺指数(肺质量/体质量)和肺质量抑制率。
肺质量抑制率=(对照组小鼠肺质量-各药物组小鼠肺质量)/(对照组小鼠肺质量-模型组小鼠肺质量)
2.3 肺脏病理检测(HE 染色)
将各组ICR 小鼠肺脏固定于4%多聚甲醛溶液中24 h。组织浸于液体石蜡,然后进行包埋、切片和贴片。切片置于60 ℃环境中烘烤1 h,经脱蜡处理后,依次进行苏木素染色和伊红染色。切片于光学显微镜下观察并拍照。
2.4 肺脏NK 细胞Ly-49D 和Ly-49H 受体含量的检测
各组C57BL/6J 小鼠给药后第5 天,ip 氯胺酮(100 mg/kg)麻醉后45 min,脱颈椎处死,摘取肺脏,制备单细胞悬液,裂解红细胞后,每只小鼠取1×107个细胞,以FITC-Ly-49D 抗体(1.5 μL/500 μL体系)和PE-Ly-49H 抗体(2.5 μL/500 μL 体系)4 ℃共同孵育30 min,400×g离心10 min。取沉淀,用2 mL PBS 洗一遍,400×g离心10 min。将细胞沉淀用400 μL 2%多聚甲醛固定。采用流式细胞仪检测NK 细胞表面激活性受体Ly-49D、Ly-49H 的含量。同时设置Ly-49D、Ly-49H 抗体的同型对照组。
2.5 肺脏NK 细胞NKp46 受体的检测
各组C57BL/6J 小鼠给药后第5 天,ip 氯胺酮(100 mg/kg)麻醉后45 min,脱颈椎处死,摘取肺脏,制备单细胞悬液,裂解红细胞后,每只小鼠取1×107个细胞,以FITC-CD3e(1.5 μL/500 μL 体系)、PE-CD49b(2.0 μL/500 μL 体系)和Alexa Fluor 647-NKp46(2.0 μL/500 μL 体系)抗体4 ℃共同孵育30 min,400×g离心10 min。取沉淀,用2 mL PBS洗一遍,400×g离心10 min。将细胞沉淀用400 μL 2%多聚甲醛固定。采用流式细胞仪检测NK 细胞毒性受体NKp46 的含量。同时设置NKp46 抗体的同型对照组。
2.6 H3N2 流感病毒感染NK-92MI 细胞模型的制备及药物干预
将NK-92MI 细胞接种于6 孔板。实验设对照组、模型(H3N2 感染)组、银翘散组和臣药组,每组3 个复孔。将H3N2 病毒原液以无血清培养基稀释82.5 倍,作为病毒工作液。以病毒工作液培养NK-92MI 细胞6 h,1.5 mL/孔;对照组以等体积不含病毒的培养基培养。补加完全培养基1.5 mL/孔,继续培养16 h。银翘散组和臣药组分别加入银翘散和臣药储备液,使质量浓度分别为50、100 μg/mL;对照组加入等体积的高纯水。药物作用10 h。
2.7 NK-92MI 细胞转录组分析
收集“2.6”项中的细胞,提取细胞总RNA,分离并纯化mRNA,构建cDNA 文库,然后采用Illumina 测序平台(Illumina HiSeqTM2500)对文库进行测序(上海欧易生物医学科技有限公司)。对差异基因进行基因本体论(gene ontology,GO)和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)富集分析。
2.8 统计方法
数据以±s表示,采用SPSS 13.0 软件中单因素方差分析,P<0.05 表示差异具有统计学意义。
3 结果
3.1 对流感病毒感染小鼠肺炎的抑制作用
流感病毒PR8 的感染使得小鼠体质量下降、肺脏质量增加,进而肺脏指数显著高于正常小鼠(P<0.01)。与模型组相比,阳性药物利巴韦林、银翘散、君药(金银花、连翘)、臣药(薄荷、牛蒡子、淡豆豉、荆芥穗)、佐药(桔梗、芦苇根、淡竹叶)和使药(甘草)均能够降低由流感病毒引起的肺指数增加;其中利巴韦林、银翘散和臣药能够显著抑制肺指数的增加(P<0.05、0.01);且臣药组对肺炎的抑制作用略优于银翘散组。结果见表1。
表1 银翘散及其拆方对流感病毒感染小鼠肺指数的影响(±s,n=10)Table 1 Effect of YQS and its recipes on lung index of mice infected with PR8 virus (±s,n=10)
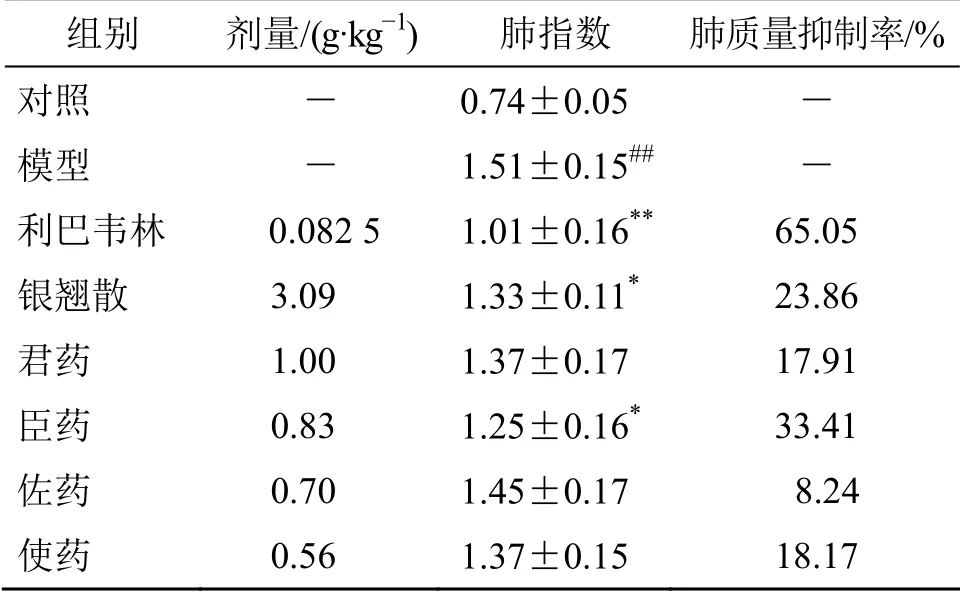
表1 银翘散及其拆方对流感病毒感染小鼠肺指数的影响(±s,n=10)Table 1 Effect of YQS and its recipes on lung index of mice infected with PR8 virus (±s,n=10)
与对照组比较:##P<0.01;与模型组比较:*P<0.05 **P<0.01##P < 0.01 vs control group; *P < 0.05 **P < 0.01 vs model group
组别 剂量/(g·kg-1) 肺指数 肺质量抑制率/%对照 - 0.74±0.05 -模型 - 1.51±0.15## -利巴韦林 0.082 5 1.01±0.16** 65.05银翘散 3.09 1.33±0.11* 23.86君药 1.00 1.37±0.17 17.91臣药 0.83 1.25±0.16* 33.41佐药 0.70 1.45±0.17 8.24使药 0.56 1.37±0.15 18.17
通过以上结果发现,利巴韦林和银翘散臣药的药效优于银翘散全方,故针对对照组、模型组、利巴韦林组和臣药组小鼠的肺脏进行了病理分析。肺脏病理检测(HE 染色)显示,对照组小鼠肺脏可见清晰肺泡;肺泡、支气管内无任何细胞浸润,支气管壁绒毛排列整齐。模型组小鼠(PR8 流感病毒感染)肺泡腔缩小,甚至塌陷,支气管中可见白细胞聚集,支气管壁绒毛细胞排列杂乱。经利巴韦林和臣药治疗的小鼠肺脏仍可见大量完好的肺泡,支气管壁绒毛排列整齐,白细胞浸润减少;利巴韦林组肺泡内仍可见因红细胞从血管漏出引起的水肿。见图1。
3.2 对流感病毒感染小鼠肺脏NK 细胞激活性受体Ly-49D、Ly-49H 含量的影响
与正常C57BL/6J 小鼠相比,PR8 流感病毒感染小鼠后使NK 细胞激活性受体Ly-49D、Ly-49H在肺脏的含量显著降低(P<0.01)。经利巴韦林、银翘散、君药、臣药、佐药和使药治疗后,各组小鼠肺脏2 种受体含量均明显增加,除佐药组外,与模型组相比,其余各组差异显著(P<0.05、0.01),见表2。结果显示,各药物诱导2 种受体表达的作用由强到弱依次为臣药>使药>银翘散>君药>佐药。各组小鼠肺脏NK 细胞激活性受体Ly-49D和Ly-49H 流式图见图2。

图1 臣药对流感病毒PR8 感染小鼠肺脏病理的影响 (HE 染色,×200)Fig.1 Effect of Ministerial drug on pulmonary pathology in mice infected with influenza virus PR8 (HE staining,× 200)
表2 银翘散及其拆方对流感病毒感染小鼠肺脏NK 细胞激活性受体Ly-49D、Ly-49H 含量的影响 (±s,n=3)Table 2 Effects of YQS and its recipes on content of Ly-49D and Ly-49H in lungs of mice infected with PR8 virus(±s,n=3)
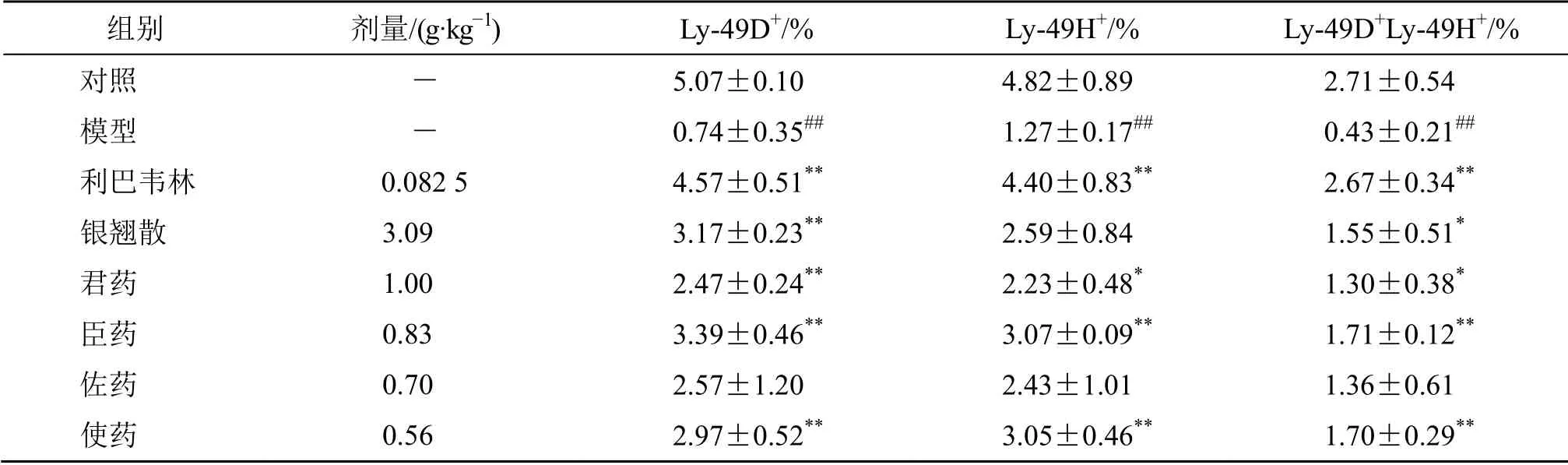
表2 银翘散及其拆方对流感病毒感染小鼠肺脏NK 细胞激活性受体Ly-49D、Ly-49H 含量的影响 (±s,n=3)Table 2 Effects of YQS and its recipes on content of Ly-49D and Ly-49H in lungs of mice infected with PR8 virus(±s,n=3)
与对照组比较:##P<0.01;与模型组比较:*P<0.05 **P<0.01##P < 0.01 vs control group; *P < 0.05 **P < 0.01 vs model group
组别 剂量/(g·kg-1) Ly-49D+/% Ly-49H+/% Ly-49D+Ly-49H+/%对照 - 5.07±0.10 4.82±0.89 2.71±0.54模型 - 0.74±0.35## 1.27±0.17## 0.43±0.21##利巴韦林 0.082 5 4.57±0.51** 4.40±0.83** 2.67±0.34**银翘散 3.09 3.17±0.23** 2.59±0.84 1.55±0.51*君药 1.00 2.47±0.24** 2.23±0.48* 1.30±0.38*臣药 0.83 3.39±0.46** 3.07±0.09** 1.71±0.12**佐药 0.70 2.57±1.20 2.43±1.01 1.36±0.61使药 0.56 2.97±0.52** 3.05±0.46** 1.70±0.29**

图2 各组小鼠肺脏NK 细胞激活性受体Ly-49D 和Ly-49H 流式图Fig.2 Flow diagram of NK cell activating receptors Ly-49D and Ly-49H in mouse lung of each group
3.3 对流感病毒感染小鼠肺脏NK 细胞毒性受体NKp46 含量的影响
实验中首先分选出CD3e 阴性(CD3e-)细胞,即排除T 淋巴细胞。在CD3e-细胞群中,CD49b阳性(CD49b+)的细胞即为NK 细胞。NKp46 是NK 细胞表面的1 个激活性受体,主要介导细胞毒性反应。正常C57BL/6J 小鼠肺脏表达一定量的NKp46 受体。当PR8 流感病毒感染C57BL/6J 小鼠后,小鼠肺脏NK 细胞表面NKp46 受体的含量出现减少的现象。通过小鼠肺脏NK 细胞激活性受体L49D/H 的检测对各药物活性进行初筛,发现臣药的药效最好,因此选择臣药和银翘散进一步进行小鼠肺脏NK 细胞毒性受体NKp46 含量的检测。经银翘散和臣药治疗的PR8 病毒感染小鼠,NKp46 受体含量均高于模型组,且臣药组效果优于银翘散组。抗病毒阳性药物利巴韦林不能改善因病毒感染引起的肺脏NK 细胞NKp46 受体含量减少的现象(图3)。
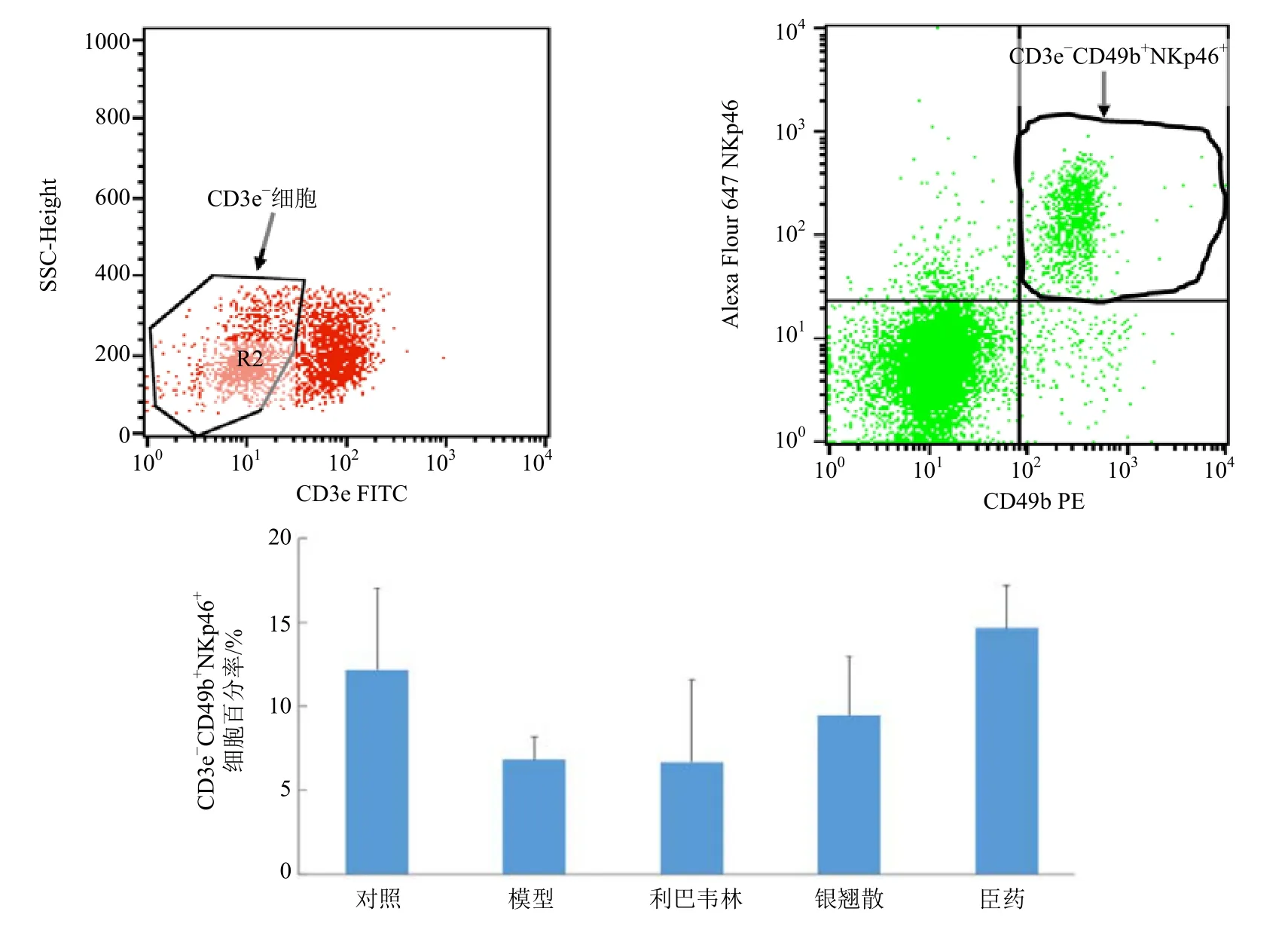
图3 银翘散及其拆方对流感病毒PR8 感染小鼠肺脏NK 细胞细胞毒性受体NKp46 含量的影响 (±s,n=3)Fig.3 Effect of YQS and its recipes on expression of cytotoxic receptor NKp46 in pulmonary NK cells of mice infected with influenza virus PR8 (±s,n=3)
3.4 对流感病毒感染自然杀伤细胞转录组的影响
3.4.1 主成分分析和聚类分析 主成分分析能有效展示基因表达差异对样本的影响;基因表达相似的样本距离近,说明样本相似。聚类分析通过计算样本之间的距离,考察样本之间的相似性。如图4 所示,对照组、模型组、臣药组和银翘散组,同一组别的样本距离相近,不同组之间在距离上能够区分开来;不同处理方法引起NK-92MI 细胞mRNA 表达不同。
3.4.2 差异基因GO 富集分析 在得到差异基因后,从分子和细胞水平,根据基因的功能,将功能相似的基因富集为一类。目前GO 分析有3 个亚类,包括生物过程(如信号转导)、分子功能(如酶活性)和细胞成分(如核糖体)。每个亚类根据功能不同划分出不同条目。在GO 富集分析中,在差异最显著的前30 条上调基因条目中,银翘散组和臣药组有16个条目相同;相同条目中,臣药组上调基因不仅涵盖了银翘散组中表达上调的基因,而且上调基因种类比银翘散组更广泛。具体条目和基因名称见表3和图5。在相同上调基因中,DDIT3、JUN、FOS、EGR1、EGR2、ATF、AREG、TNFAIP3、PPP1R15A、ZFP36L110 个基因出现频次较高,这些基因主要参与细胞生长和凋亡过程。不同上调基因条目见表4。

图4 H3N2 流感病毒感染NK-92MI 细胞转录组数据主成分分析 (A) 及聚类分析 (B)Fig.4 Principal component analysis (A) and cluster analysis (B) of cell transcriptome data of NK-92MI infected with H3N2 influenza virus

表3 GO 富集分析中臣药和银翘散组相同上调的基因Table 3 Same up-regulated genes in Ministerial drug and YQS group based by GO enrichment analysis

图5 银翘散和臣药组相同上调基因条目中基因维恩图Fig.5 Venn diagram of genes in same up-regulated genes items of Ministerial drug and YQS groups

表4 GO 富集分析中银翘散和臣药组不同上调基因条目Table 4 Different up-regulated gene entries in Ministerial drug and YQS groups based on GO enrichment analysis
在GO 富集分析的下调基因条目中,臣药组富集到30 个条目,银翘散组仅富集到10 个条目,具体基因条目见表5。其中仅nucleus 条目与臣药组相同,臣药下调该条目中的51 个基因(FOXD4L1、EID3、SSH3、CCNE1、CENPBD1、SYCP3、PER3、CDKN2C、HSPA1B、PHF13、HEXIM1、FIGNL2、TREX2、ZNF296、ZBED1、UBC、H2AFX、CBX2、NFKBIE、RBM14、TUBB2A、HSPA5、TUBB4B、FEN1、FBXO43、FAM71F2、CABYR、EID2B、TEF、GADD45B、ATF5、BCL9L、CEMIP、MYLK2、ZCCHC5、PHF7、SERTAD3、FOXD2、FOXD4、TUBA1C、IMP3、TIGD1、TIGD3、TIGD4、TBR1、FOXJ1、DYRK3、ZFC3H1、UTP3、RARG、HIST1H2AG),银翘散仅下调该条目中的5 个基因(FOXD4L1、EID3、SSH3、TRIM22、AKAP17A)。
3.4.3 差异基因KEGG 富集分析 利用KEGG 数据库对差异基因进行信号通路(pathway)分析,可以找到富集差异基因的pathway 条目,寻找不同样本的差异基因可能与哪些通路的改变有关。在富集的差异最显著的前20(Top 20)条信号通路中,银翘散组和臣药组有13 条通路重合,见表6。相同信号通路所涉及到的上调基因,臣药组基本囊括了银翘散组的上调基因,且臣药组上调的基因数目比银翘散组多(图6)。臣药和银翘散组富集到的不同信号通路见表7。
在KEGG 富集到的Top 20 下调信号通路中,银翘散组仅富集到17 条信号通路;其中,仅necroptosis信号通路同时被银翘散和臣药调节,见表8。

表5 GO 富集分析中银翘散和臣药组下调基因条目Table 5 Down-regulated gene entry in Ministerial drug and YQS groups based on GO enrichment analysis
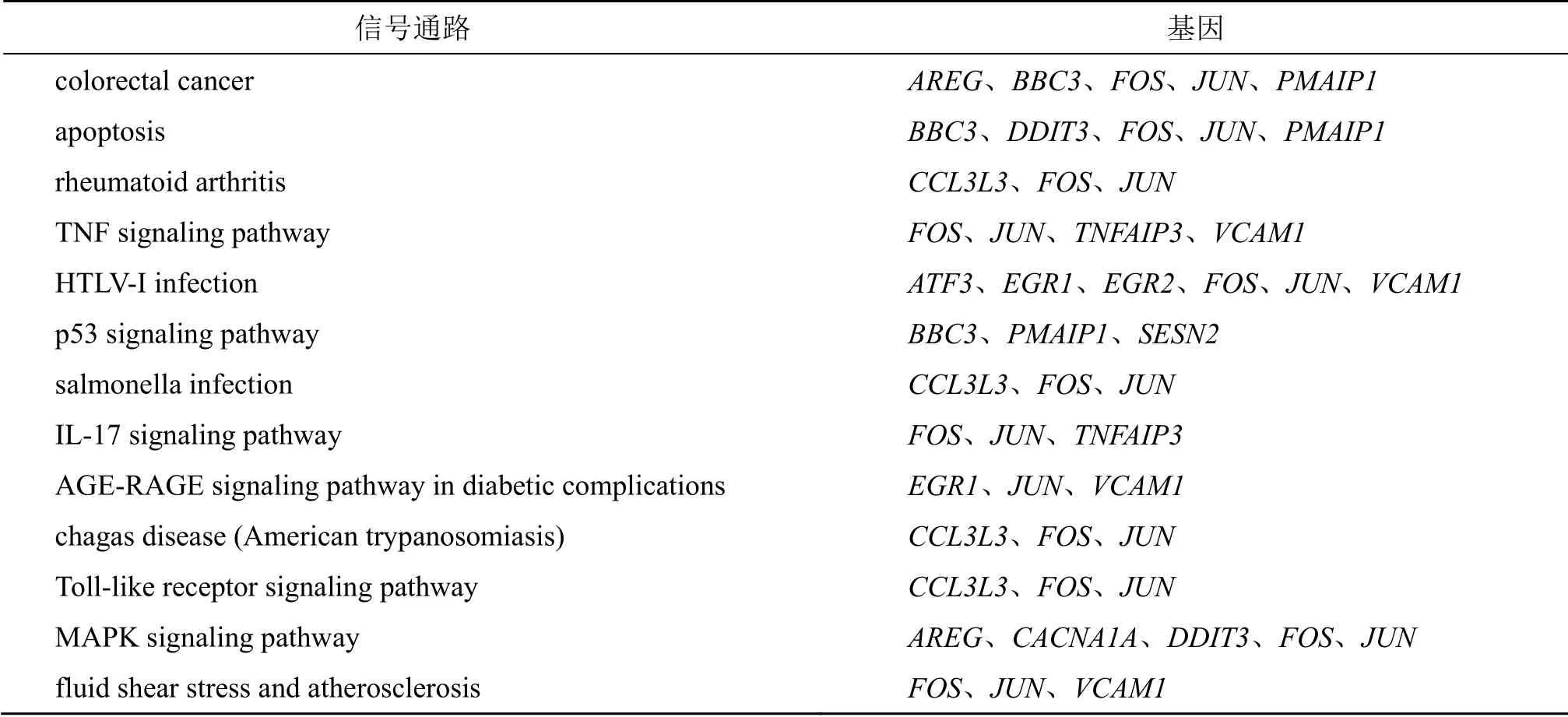
表6 KEGG 富集分析中银翘散和臣药组相同上调信号通路及基因Table 6 KEGG enrichment analysis of same up-regulated signal pathways and genes in Ministerial drug and YQS groups
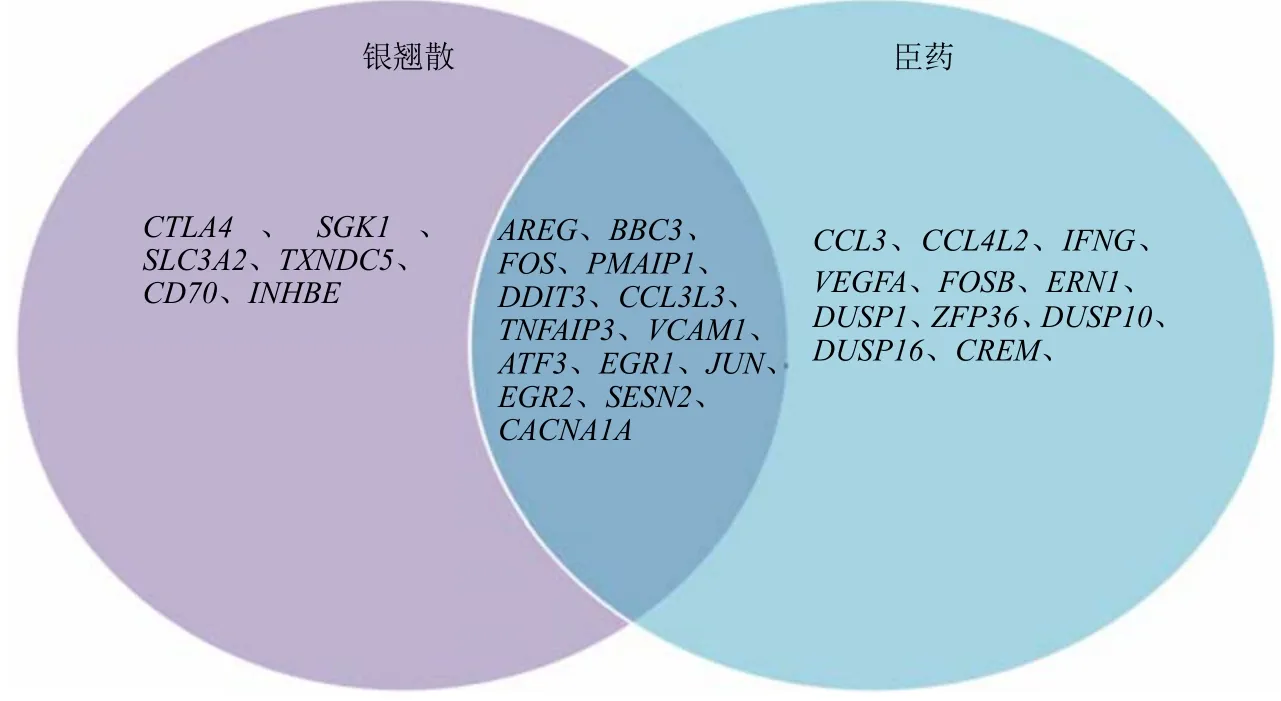
图6 银翘散和臣药组相同上调信号通路中基因的维恩图Fig.6 Venn diagram of genes in same up-regulated signal pathway of Ministerial drug and YQS groups

表7 KEGG 富集分析中银翘散和臣药组不同上调信号通路Table 7 Different up-regulated signal pathways in Ministerial drug and YQS groups based on KEGG enrichment analysis

表8 KEGG 富集分析中银翘散和臣药组下调信号通路(前20)Table 8 Down-regulated signal pathways (Top 20) in Ministerial drug and YQS groups based on KEGG enrichment analysis
4 讨论
本研究显示,银翘散及其拆方治疗病毒性肺炎药效强度为臣药>银翘散>使药>君药>佐药,药效强度并非与君、臣、佐、使的中药配伍理论完全一致。金银花和连翘为君药,疏散风热、清热解毒、避秽化浊;臣药薄荷和牛蒡子疏散风热、清利头目、解毒利咽,荆芥穗和淡豆豉发散解表;佐药淡竹叶、芦苇根清热生津,桔梗宣肺止咳;生甘草调和诸药,又合桔梗利咽止咳,为佐使之用[7]。从银翘散的方解可以看出,金银花和连翘的功效相当于现代药理学中抗病毒、抗菌的作用,而解热作用略微。薄荷、牛蒡子、荆芥穗和淡豆豉构成的臣药组,其功效涵盖了现代药理学的解热、抗病毒、抗菌的作用。淡竹叶、芦苇根和桔梗都有解热作用,桔梗还具有镇咳的作用[8]。现代药理学研究表明生甘草的作用较为广泛,包括免疫增强、抗病毒、抗菌、抗炎、镇咳、祛痰等作用[9-10]。采用现代药理学方法对中药复方的配伍进行探究,能够明确各配伍组分对复方整体药效的贡献程度。
NK 细胞在清除流感病毒中发挥着关键作用。人类NK 细胞表面存在2 类受体:激活性受体(killer activating receptor,KAR)和抑制性受体(killer inhibiting receptor,KIR),它们分别介导NK 细胞的激活和抑制。与人类NK 细胞相似,小鼠NK 细胞表面也存在这2 类受体。研究证实[11],小鼠感染流感病毒后,Ly-49 抑制性受体(Ly-49I 和Ly-49Q)缺陷的小鼠比野生型小鼠存活时间更长;而当回补表达Ly-49I 和Ly-49Q 受体后,小鼠又恢复了对流感病毒的敏感性。提示NK 细胞参与流感病毒的疾病过程,Ly-49 受体直接参与了NK 细胞清除流感病毒的免疫应答效应。
Ly-49 基因家族受体主要表达在小鼠NK 细胞,是C 型凝集素超家族成员之一。Ly-49D 是Ly-49 家族中第一个被证实传递激活性信号的蛋白分子[12-13],属于KAR 类受体,Ly-49D 受体介导的生物学效应是促进NK 细胞活化,进而促使NK 细胞发挥杀伤效应。PR8 流感病毒感染显著下调Ly-49D 受体介导的促炎细胞因子和趋化因子的产生[14]。
Ly-49H是Ly-49 家族的另一个激活性受体,其激活效应由NK 细胞表面的跨膜接头蛋白DAP12(DAP12 的胞质域含有免疫受体酪氨酸激活基序)介导。Ly-49H 的抗病毒效应最早发现于小鼠巨细胞病毒(murine cytomegalovirus,MCMV)。表达Ly-49H 受体的转基因小鼠更不易感染MCMV[15]。当 Ly-49H-DAP12 受体复合物突变的小鼠感染MCMV 时,脾脏病毒滴度增加30~40 倍,肝脏病毒滴度增加2~5 倍,且肝脏NK 细胞产生γ 干扰素(interferon-γ,IFN-γ)的比例降低了(56.00±0.22)%,说明Ly-49H的正常活化对抑制MCMV具有重要作用[16]。有研究还发现,Ly-49H 受体表达呈阳性的NK 细胞会在小鼠淋巴组织和非淋巴组织存在数月;在MCMV 感染小鼠的恢复期,当再次感染MCMV 时,Ly-49H 受体表达呈阳性的NK 细胞会被重新激活,并迅速发生脱颗粒、释放细胞因子[17]。这打破了人们对NK 细胞在传统意义上的认识:NK细胞不仅参与抗病毒反应的固有免疫应答,而且同样参与了获得性免疫应答。关于Ly-49H 受体在流感病毒中的作用,目前还未见相关文献报道。本研究中,PR8 流感病毒的感染使Ly-49H 受体在肺脏的含量降低,经抗病毒药物利巴韦林和银翘散及其拆方(臣药、使药、君药和佐药)治疗后,Ly-49H受体含量均升高;而且,这些药物对流感病毒PR8感染引起的小鼠肺炎有抑制作用。上述药物对肺炎的抑制作用和对肺脏中Ly-49D、Ly-49H 含量的促进作用基本一致,其作用强度均为臣药>银翘散/使药>君药>佐药。说明上述药物对病毒性肺炎的抑制作用越强,NK 细胞激活性受体(L-49D和Ly-49H)在肺脏中的含量也越高。
NK 细胞的细胞毒效应是NK 细胞杀伤靶细胞的重要途径,该效应主要由细胞毒性受体介导。人类主要有3 种细胞毒性受体:NKp30、NKp44 和NKp46。小鼠NK 细胞的细胞毒性反应由NKp46 介导;小鼠的NKp46 又称作NCR1(natural cytotoxicity receptor 1)。NKp46 识别流感病毒血凝素[18],使NK细胞能够识别病毒感染的细胞,进而损伤靶细胞,防止流感病毒感染宿主细胞[19]。有研究者采用绿色荧光蛋白(green fluorescent proteins,GFP)报告基因标记的NKp46 受体示踪NK 细胞,发现在小鼠感染流感病毒PR8 后的5 d 内,肺脏、血液、脾脏和骨髓均观察到GFP,且肺脏中GFP 数量最多,感染第5 天的数量高于前4 d;并证明NKp46 对于体内流感病毒的清除发挥了关键作用[20]。本研究中,小鼠肺脏中NKp46 受体因感染PR8 流感病毒含量降低,银翘散和臣药均能增加NKp46 受体的含量,这可能是银翘散和臣药抑制小鼠感染PR8病毒引起肺炎的作用途径之一。
本研究分别从药效学(肺质量抑制率)和作用机制(NK 细胞激活性受体和细胞毒性受体)2 方面证实:银翘散中臣药(薄荷、牛蒡子、淡豆豉、荆芥穗)的药效与银翘散全方相似,且略优于银翘散。这为银翘散的精简提供了现代药理学依据,臣药可以作为一个独立的复方用于流感病毒引起的感冒的治疗。为了进一步探究臣药优于银翘散的原因,本实验建立了人流感病毒HK8 感染人NK 细胞株NK-92MI 的细胞模型,分别以银翘散和臣药进行干预,分析药物对NK 细胞转录组的影响。
银翘散显著上调的基因在臣药组也同样显著上调;且很多基因在臣药治疗后上调,而银翘散不能显著上调这些基因(图5)。如对细胞表面成分的影响,银翘散上调6 个基因(VCAM1、SLC3A2、HRG、AREG、KCNE2、SLC7A11),而臣药同时还上调VEGFA、HBEGF、TIGIT3 个基因。T 细胞免疫球蛋白和免疫受体酪氨酸基抑制基序结构域( T-cell immunoglobulin and immunoreceptor tyrosine-based inhibitory motif domain,TIGIT)是一种新型的抑制性受体,在T 细胞和NK 细胞上均有表达。TIGIT 编码的蛋白有助于辅助性T 细胞和树突状细胞之间的相互作用,以调节T 细胞依赖的B细胞的反应。TIGIT 虽被定义为抑制性受体,但研究表明,NK 细胞上TIGIT 受体表达阳性的野生型小鼠对各种刺激的反应增强,而TIGIT 受体缺陷会损害 NK 细胞的“missing-self”识别功能。TIGIT-CD155 信号通路能够使NK 细胞更好地作为效应细胞参与到免疫应答反应中,而这一通路是不依赖于主要组织相容性复合体 I ( major histocompatibility complex I,MHC I)类分子的新的NK 细胞激活的途径[21]。
再如有关分子功能的“RNA polymerase II regulatory region sequence-specific DNA binding”条目,银翘散显著上调4 个基因(EGR1、ATF3、NR4A2、NR4A3),而臣药除上调这4 个基因外,同时还上调GLIS1、CREBRF、BCL6、CREM、FOSL2、ARID5B6 个基因。如 cAMP 反应元件调节子(cAMP responsive element modulator,CREM)编码的蛋白是一种转录因子,能够结合病毒中的环磷酸腺苷(cyclic adenosine monophosphate,cAMP)响应元件,而且它是细胞因子生成重要的调节因子[22]。FOS 相关抗原2(Fos-related antigen 2,FOSL2)编码的蛋白是亮氨酸拉链蛋白,它可以与JUN 家族的蛋白形成二聚体,从而形成转录因子复合物AP-1,主要参与细胞增殖、分化和转化的调节。研究报道,FOSL2是一个新颖的参与NK 细胞成熟和功能发挥的转录因子[23]。富含AT 序列的相互作用结构域蛋白5B(AT-rich interactive domain-containing protein 5B,ARID5B)的编码产物是DNA 结合蛋白,它是一个富含AT 的结构域。研究显示,降低NK-92MI 细胞中ARID5B的表达,会引起细胞中线粒体氧化代谢损伤和电子传递链相关基因表达下调,进一步表现出细胞生存受损和IFN-γ 生成减少;相反,过表达ARID5B后,NK-92MI 细胞氧化代谢和IFN-γ 的生成均增加[24]。对于GO 分析中富集到的下调基因,臣药富集到30 个条目,银翘散仅富集到10 个条目(表5);说明臣药对NK 细胞的作用位点远多于银翘散。
在KEGG 富集分析的TOP20 上调信号通路中,臣药和银翘散有13 条信号通路相同,且臣药上调的基因不仅包含银翘散上调的基因,还富集到其他基因(图6)。另外,对于相同信号通路,臣药和银翘散也出现不同的调节方式。如雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路,银翘散对该通路表现为显著上调(表7),而臣药对该通路表现为显著下调(表8);且富集到的基因也不同,银翘散上调SESN2、SGK1、SLC3A23 个基因,臣药下调FZD2、TNF、WNT10B3 个基因。
由此推测臣药和银翘散在转录水平对NK 细胞中基因的调节作用相似,但臣药的作用更为广泛。臣药的调节作用不仅涵盖了银翘散的全部作用,涉及到的作用点还比银翘散更为广泛,这或许是臣药药效与银翘散相似的一个重要原因。
综上所述,本研究首先从药效学和效应相关机制证明,对于流感病毒感染小鼠引起的肺炎,臣药的治疗作用与银翘散相似,且略优于银翘散;并证明该药效与NK 细胞活性增强相关。进一步采用转录组学分析得知,臣药对NK 细胞中基因的调节作用比银翘散更为广泛。本研究为银翘散的拆方研究提供研究思路,亦为其组分臣药作为一个独立的复方、替代银翘散用于临床提供实验依据。
利益冲突所有作者均声明不存在利益冲突
