美洲黑杨与小叶杨杂交F1代扦插无性系苗生长性状动态分析
2021-02-02陈玉华吴海楠陶申童吴吉妍杨文国童春发
陈玉华,姚 丹,吴海楠,陶申童,吴吉妍,杨文国,童春发*
(1. 南京林业大学林学院,林木遗传与生物技术教育部重点实验室,南方现代林业协同创新中心,江苏 南京 210037; 2.金陵科技学院动物科学与技术学院,江苏 南京 210038)
美洲黑杨(Populusdeltoides)属于杨柳科(Salicaceae)杨属(Populus)黑杨派,原产北美洲大陆中部和东部,是我国杨树自然分布区中唯一缺乏的种,于20世纪70年代引入我国[1-2]。其树干高大通直,树皮光滑或纵裂,常灰白色;生长迅速,抗叶斑病能力强,但抗寒抗旱以及生根能力较差;材质优良,是我国短轮伐期工业用材经营重要树种,在商业化集约栽培人工林中占主体地位[3-4]。美洲黑杨种质资源丰富,生长、形态等表型性状在种源间、种源内家系间以及家系内无性系间等多个水平上存在广泛的遗传变异,成为杨树选育改良的重要材料[5-6],并基于此培育了很多生产中广泛应用的优良品种,如南林系统、中林系统等的优良无性系[3]。小叶杨(P.simonii)属于杨柳科杨属青杨派,是我国北方地区主要乡土树种和栽培树种。幼树皮灰绿色,老树皮暗灰色、纵裂;芽细长,先端长尖;叶菱状卵形、菱状椭圆形或菱状倒卵形,中部以上较宽,先端骤尖或渐尖,基部楔形、宽楔形或窄圆,具细锯齿,无毛;叶柄圆,有毛或无毛。小叶杨有较强的抗旱、抗盐碱、抗逆境能力,并且生根能力很强,种内形态差异多样,在林木各项抗性育种工作中成为世界性重要基因资源[7-8]。由于两种杨树无论在形态还是在生物学特性上都有很大的差异,因此两者杂交所形成的F1代是研究杨树重要数量性状的很好材料。
杨树分子育种中一项重要的基础研究是构建遗传连锁图谱,并在此基础上识别控制重要数量性状基因在基因组上的位置及其遗传效应的大小。为此,近二三十年以来有很多有关杨树遗传图谱构建和数量性状基因位点(QTL)定位研究的报道[9-19]。在这些研究中,首先要建立一个作图群体,也就是选择两个亲本进行杂交获得一个含有大量子代的杂交群体。然后,获取群体中适当数量个体的分子标记和表型性状数据,从而可以开展遗传图谱构建和QTL定位的研究工作。杨树作图群体中子代一般都是实生苗,单株树可能受到虫害、风灾等因素的影响而折断甚至死亡,因此群体中的个体数会逐年减少,一些即使存活的单株其表型性状测量值也不准确,这会直接影响到QTL定位的准确性,也可能是过去一些杨树生长性状QTL定位研究中存在结果不一致现象的原因之一[9, 15, 17-18, 20]。为了解决这个问题,可以利用F1代的实生苗建立像杨树无性系测定那样的随机区组试验林[21-24],不但可以达到以多个无性系单株来长期保存单个F1代实生苗基因型的目的,而且可以准确地测定实生苗后期表型性状值,从而可以极大地提高杨树QTL研究的精确性。
2009—2011年,本课题组建立了美洲黑杨×小叶杨杂交F1代作图群体,于2017年利用已存活的子代个体建立了无性系随机区组试验林。本研究以美洲黑杨×小叶杨F1代的无性系试验林,对其1年和2年生的扦插苗在不同生长阶段的苗高和地径进行了测定,采用多变量方差分析法分析不同无性系、不同区组生长性状的差异性,使用随机效应模型估算不同时间点无性系重复力,利用聚类分析方法对不同无性系的苗高进行分类,以期发现F1代无性系生长性状的变异规律,为后续利用该随机区组试验林进行QTL定位研究提供理论参考。
1 材料与方法
1.1 试验地概况
试验地位于江苏省句容市下蜀镇南京林业大学实习林场(119°14′E,31°59′N),气候属北亚热带季风气候,年平均气温为15.2 ℃,极端最高气温为39.6 ℃,极端最低气温为16.7 ℃,日均气温高于10 ℃的植物生长期为226 d,总积温4 859.6 ℃,年均日照2 157 h,平均无霜期233 d,年平均降雨量1 055.6 mm,年均空气相对湿度79%。土壤为黄棕壤,土层深厚,质地黏重,水养分条件中等。
1.2 试验设计
试验材料来自美洲黑杨×小叶杨杂交F1代群体。母本为美洲黑杨(南方型) ‘I-69’,父本为小叶杨。2009—2011 年连续3年对同样的亲本进行人工授粉杂交试验,共获得F1代苗木400 余株。2012 年春季移植到南京林业大学下蜀林场。2017年春选取生长良好的234株美洲黑杨×小叶杨F1代个体,采集朝向一致、年龄相同(2年以下)、粗度1~2 cm、长度18~20 cm的枝条,进行扦插。扦插按完全随机区组试验设计,3次重复,每小区1个无性系,每无性系6株2行,株行距50 cm×60 cm,共计扦插4 212个插条。2017年6月8日开始对苗高进行观测,其后每隔约15 d 观测1次,直至2017年10月14日所有苗木驻芽为止,对苗高共进行了8次观测。2018年5月24日开始每隔约20 d观测1次,对苗高共进行了7次观测。2017和2018年底,对每株树的地径各测量1次。
1.3 数据处理
美洲黑杨×小叶杨F1代无性系苗高和地径以单株测定数据进行统计分析。在统计分析前,对原始数据进行异常值查找、剔除,运用R语言对每年多次观测的苗高数据绘制散点连线图,删除由于死亡或风折等原因造成苗高没有随时间增加而增长的单株数据。应用SPSS 22.0统计软件对两个生长性状数据进行多变量方差分析,利用HalfsibSS 1.0软件[25]计算无性系重复力,使用R语言包Hclust根据无性系在多个时间点的效应对无性系进行聚类分析。
多变量方差分析和重复力计算使用如下线性模型:
yijkt=μt+Bit+Fjt+BFijt+eijkt。
(1)
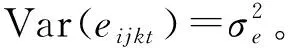
在进行无性系聚类分析时,首先将模型(1)视作固定效应模型,并考虑如下约束条件:
(2)
计算出各个无性系效应的最佳线性无偏估计[26]。然后根据无性系在多个时间点的效应值,采用欧式距离计算无性系间的距离,类间距离采用系统聚类法中的Ward法(离差平方和法),通过R语言软件Hclust函数实现聚类分析,同时调用Plot函数绘制扇形聚类图。根据肘部法则(Elbow method)确定参试无性系分类的个数[27]。
2 结果与分析
2.1 美洲黑杨小叶杨F1代无性系苗高、地径的方差分析

图1 美洲黑杨×小叶杨F1代无性系随机区组试验林2年生苗高和地径直方图Fig.1 Histograms for the tree height and ground diameter in the clonal randomized block testforest derived from the F1 progeny of Populus deltoides×P. simonii
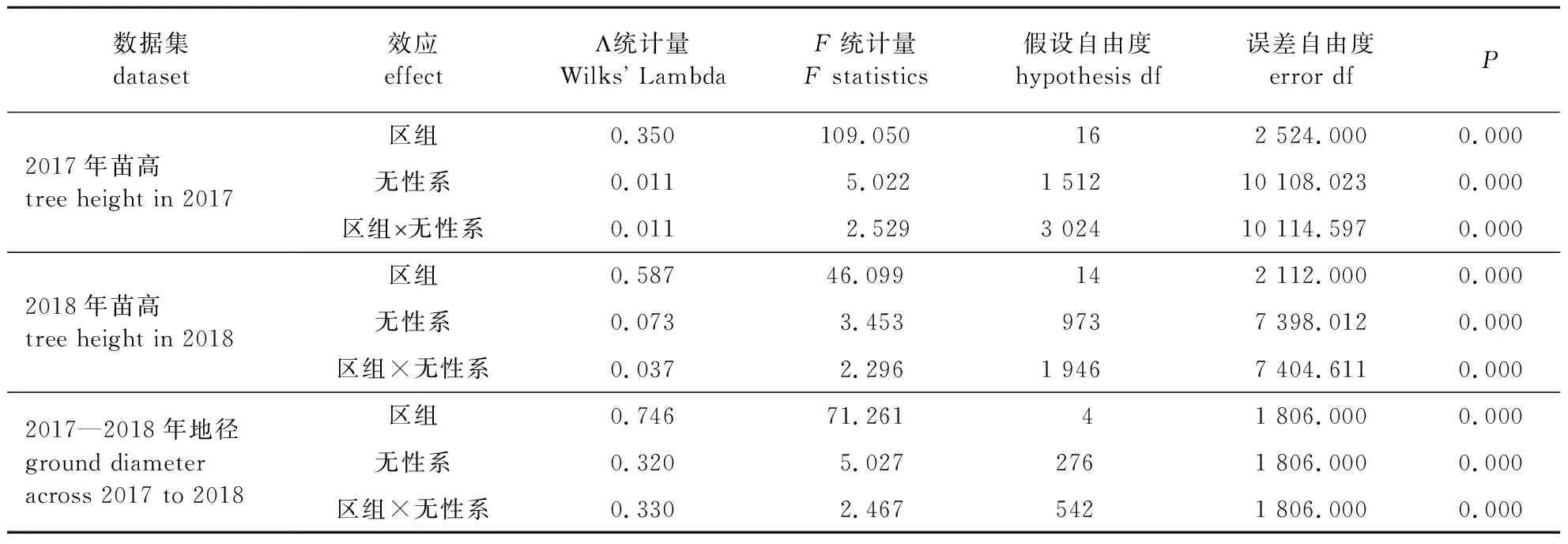
表1 美洲黑杨×小叶杨F1代无性系随机区组试验林苗高和地径多变量方差分析
2.2 美洲黑杨×小叶杨F1代无性系试验林苗高和地径变异参数
美洲黑杨×小叶杨F1代无性系在扦插后2年中若干个生长时间点苗高和地径变异参数如表2所示。可以看出,在2年连续观测中,苗高总体平均值从34.67 cm逐渐增长至317.44 cm,标准差从11.08 cm逐渐增长至60.97 cm,变异范围从极差76.7 cm逐渐增长至425.0 cm,变异系数从31.96%逐渐降低至19.21%。表明随着树木月龄的增长,美洲黑杨×小叶杨F1代无性系苗高变异绝对值在增大,但相对值在减小。地径2年的总体平均数分别为1.50 cm和2.02 cm,变化范围极差分别为2.90 cm和3.58 cm,标准差分别为0.40 cm和0.53 cm,变异系数分别为26.66%和26.20%,说明地径也是随着树龄的增长,变异绝对值在增大,但相对值在减小,与苗高表型变异趋势基本上一致。
从表2还可以看出,随着树龄的增长,苗高重复力先逐渐减小,并在2018年度第1次测量时达最低值0.398,而后又随着树龄的增长而逐渐增大。总的来说,苗高重复力变动范围为0.398~0.689,说明美洲黑杨×小叶杨F1代无性系苗高受遗传因素的影响在不同生长时期不是固定不变的,有一定变动幅度,基本上受到中等强度以上的遗传因素控制。地径的重复力值2017年较2018年低,分别为0.376和0.594,表明随着时间的变化,地径同苗高一样受到中等程度以上遗传因素的影响。

表2 美洲黑杨×小叶杨F1代无性系随机区组试验林苗高和地径生长性状变异
2.3 美洲黑杨×小叶杨F1代无性系苗高聚类分析
为了揭示无性系间生长性状的变异,分别对2017和2018年无性系效应最佳线性无偏估计值进行了聚类分析。根据肘部法则(Elbow method),2017年观测的190个无性系和2018年观测的139个无性系均分为8个类较适宜(表3)。图2A和图2B分别显示了2017和2018年观测的无性系的扇形聚类图,可以清楚地看出每一个无性系具体属于何种聚类。虽然2018年观测的139个无性系包含在2017年的190个无性系之中,但是2018年的无性系每个聚类不是完全包含在2017的某个聚类之中。表3给出了2017和2018年无性系两两聚类间共有的无性系数目以及2018年每个类中无性系的数目。例如,2018年的第一个类(CL1)中的28个无性系分别来自2017年的CL1、CL2、CL3、CL4和CL6类,其共有的无性系数目分别为6、1、9、8和4;其他类的无性系至少来自2017年的两个聚类,只有第8类来自2017年的1个聚类,因为该类只有1个无性系。
图2C显示了2017年在8个时间点观测的无性系每个聚类均值效应散点连线图。发现第2、7和8类的无性系均值效应随时间延长明显地增加,其中第8类(CL8)在各个时间点的效应都是最大的。与其相反,第1、3和5类的无性系均值效应随时间明显地下降,其中第5类下降最多。此外,发现第6类的均值效应在前6个观测时间点基本上平缓略微为负,但是在后2 次观测时间点则逐渐增加变为正效应;第4类均值效应的变化趋势则第6类恰恰相反,在前6个观测时间点基本上平缓略微为正,而在后2次时间点逐渐下降变为负效应。
图2D显示了2018年在7个时间点观测的无性系每个聚类均值效应散点连线图。虽然有4个类的均值效应在各个时间点均为负值,而另外4个类的效应均为正值,但是同2017年的类均值效应变化趋势明显不同,有7个类的均值效应随着时间变化比较平缓,只有第8类比较特殊只含有一个无性系,其效应前期随时间而增加而后期也比较平缓。
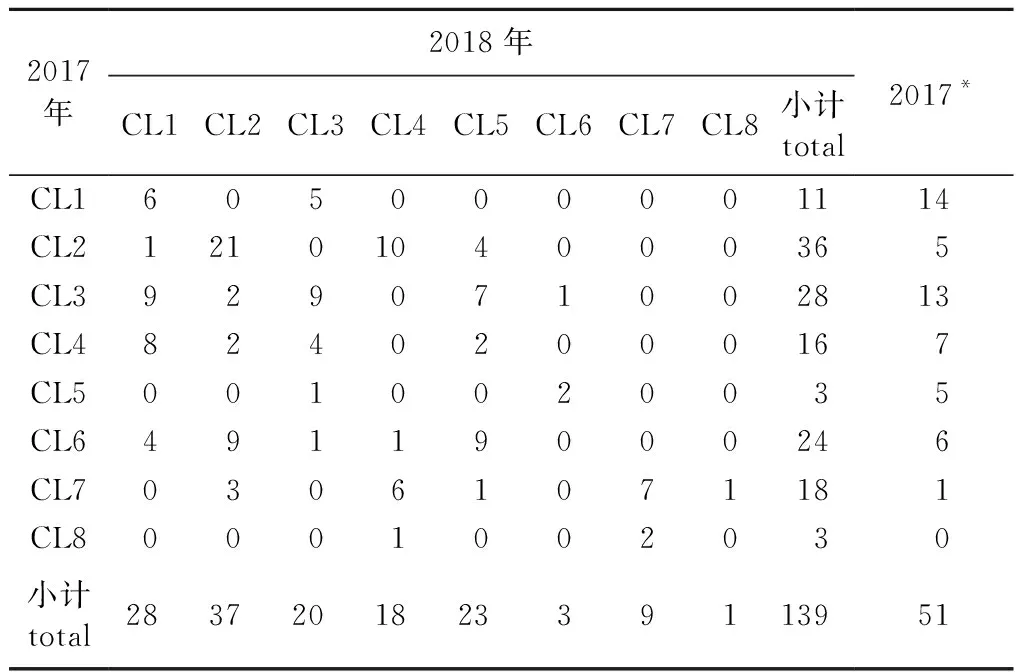
表3 2017和2018年无性系分类间共有无性系数目

图2 美洲黑杨×小叶杨F1代无性系苗高聚类分析Fig.2 The clustering analysis of clones from the F1 progeny of P. deltoides × P. simonii
3 讨 论
本研究所建立的杨树无性系随机区组试验林为杨树数量性状基因定位研究提供了一个很好的遗传作图材料。由于每一个无性系在区组内和小区内都有重复,使得1个无性系在群体中有超过10个单株,而每个无性系代表1个实生苗的基因组,因此这样的作图群体可以最大限度地保存每个子代的基因组信息,免受因自然灾害造成作图群体中子代基因组类型的减少。另一方面,由于无性系有多个重复的单株,使得它所代表的实生苗表型性状值可以有多个重复的测量值,因此使用这些无性系来进行QTL定位会更加准确。但是,在动植物育种研究中,传统的遗传作图群体为自交系中的回交(BC)和F2代群体,或是异交群体中的杂交F1代群体,一般使用类似于林木杂交F1代中实生苗的个体以及区间作图(interval mapping)[33]或复合区间作图(composite interval mapping)[34]等统计分析方法[18, 35-36]来进行QTL定位研究,这些统计方法不能直接用于本研究所述的杨树无性系随机区组试验林作图群体。因此,针对本次研究所建立的杨树无性系试验群体,依据对该群体所获得遗传分析的结果,后续要建立相应的统计方法用于QTL定位或进行全基因组关联分析[37-39],这将有助于杨树数量性状基因定位的精确性,从而加速杨树分子育种的进程。
