Nrf2及天然产物导向的Nrf2激活剂研究进展
2021-02-01程茂军谢赛赛
程茂军,郭 杰,刘 婧,谢赛赛,3*
1江西中医药大学 中药固体制剂制造技术国家工程研究中心;2江西中医药大学药学院;3江西本草天工科技有限责任公司,南昌 330004
氧化应激(oxidative stress)是指机体在受到刺激时,活性氧(reactive oxygen species, ROS)或活性氮(reactive nitrogen species,RNS)自由基的含量超过其自身可清除的范围,导致机体内氧化还原平衡失调,系统和组织损伤,最终引起一系列疾病的产生。为应对氧化应激,机体自我表达产生了一系列内源性抗氧化因子,包括抗氧化分子和解毒酶,其中,Nrf2-Keap1-ARE信号通路在介导机体产生内源性抗氧化因子过程中起到了至关重要的作用[1]。研究表明,在包括肝,肺,胃肠道,脑,皮肤,以及膀胱在内的多种组织器官中,Nrf2蛋白的缺失会导致细胞保护基因呈现出较低的基础与诱导表达水平。此外,Nrf2蛋白在癌症、神经退行性疾病、自身免疫疾病、糖尿病等疾病中发挥重要作用[2],因此,寻找高效的Nrf2激活剂已成为治疗氧化应激参与类疾病的重要策略。
天然产物对于药物发现具有重要意义,许多药物都直接或间接来源于天然产物[3]。目前,已知的绝大多数Nrf2激活剂是天然产物或天然产物的衍生物。本文阐述了Nrf2-Keap1-ARE信号通路的结构及其作用机制,并综述了近期国内外文献中报道的含有Nrf2激活活性的天然产物及其衍生物,以期为更深入的研究提供参考。
1 Keap1-Nrf2-ARE信号通路
细胞内存在多种抵御氧化应激的抗氧化因子,包括维持细胞稳态的调节性和结构性蛋白以及Ⅱ相解毒酶,如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPX)、还原型谷胱甘肽(GSH)、NAD(P)H醌氧化还原酶(NQO1)、血红素加氧酶-1(HO-1)等[4-9]。当机体的ROS或RNS水平过高时,细胞会诱导这些抗氧化因子的表达,抵御氧化应激,编码这些抗氧化因子的基因含有共同启动子元件,称为抗氧化反应元件(antioxidant response element,ARE),亦称亲电体反应元件[10](electrophile response element,EpRE)。
ARE是由Daniel实验室在研究编码谷胱甘肽巯基转移酶Ya启动子时发现的一种含有特异性DNA序列的顺式调控元件,位于编码解毒酶及细胞保护蛋白基因的上游[11]。通常,ARE诱导下游抗氧化因子的表达是通过激活转录因子E2相关因子2(nuclear factor erythroid 2 related factor 2,Nrf2)来调控的。Nrf2由Moi等[12]发现并命名,属于Cap‘n’Collar碱性亮氨酸拉链家族的一员,大量存在于中枢和外周组织器官中。调控Nrf2活性的主要因子为Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)[13]。Keap1通过Nrf2上的两个结合位点(DLG和ETCE)与Nrf2产生相互作用,再通过与Cullin3(Cul3)依赖的E3泛素连接酶复合物衔接,促进Nrf2发生泛素化介导的蛋白酶体降解,进而抑制Nrf2活性。
研究发现,一分子Nrf2能够同两分子Keap1结合,DLG和ETCE是连接Nrf2与Keap1的关键序列。正常情况下,Keap1蛋白与ETGE和DLG都产生相互作用,通过与Cullin3(Cul3)依赖的E3泛素连接酶复合物的底物衔接,导致Nrf2泛素化并被蛋白酶体降解,抑制Nrf2的活性。在氧化应激条件下,ROS、RNS、亲电试剂等能够共价修饰Keap1的半胱氨酸残基(如Cys151、Cys257、Cys273、Cys288和Cys297),导致Keap1构象发生变化,Nrf2从具有较低亲和力的DLG上释放下来,而Nrf2本身仍通过高亲和力的ETGE与Keap1相连(Nrf2与Keap1结合的此类结构模型被称为“门闩-铰链模型[14]”),从而阻止了Nrf2的泛素化,致使Nrf2与Keap1结合处于饱和状态,新合成的Nrf2易位至细胞核,与小分子肌腱纤维瘤蛋白(maf)结合成异二聚体并识别结合ARE,启动下游抗氧化因子的表达,提高细胞的抗氧化能力(图1)[15]。
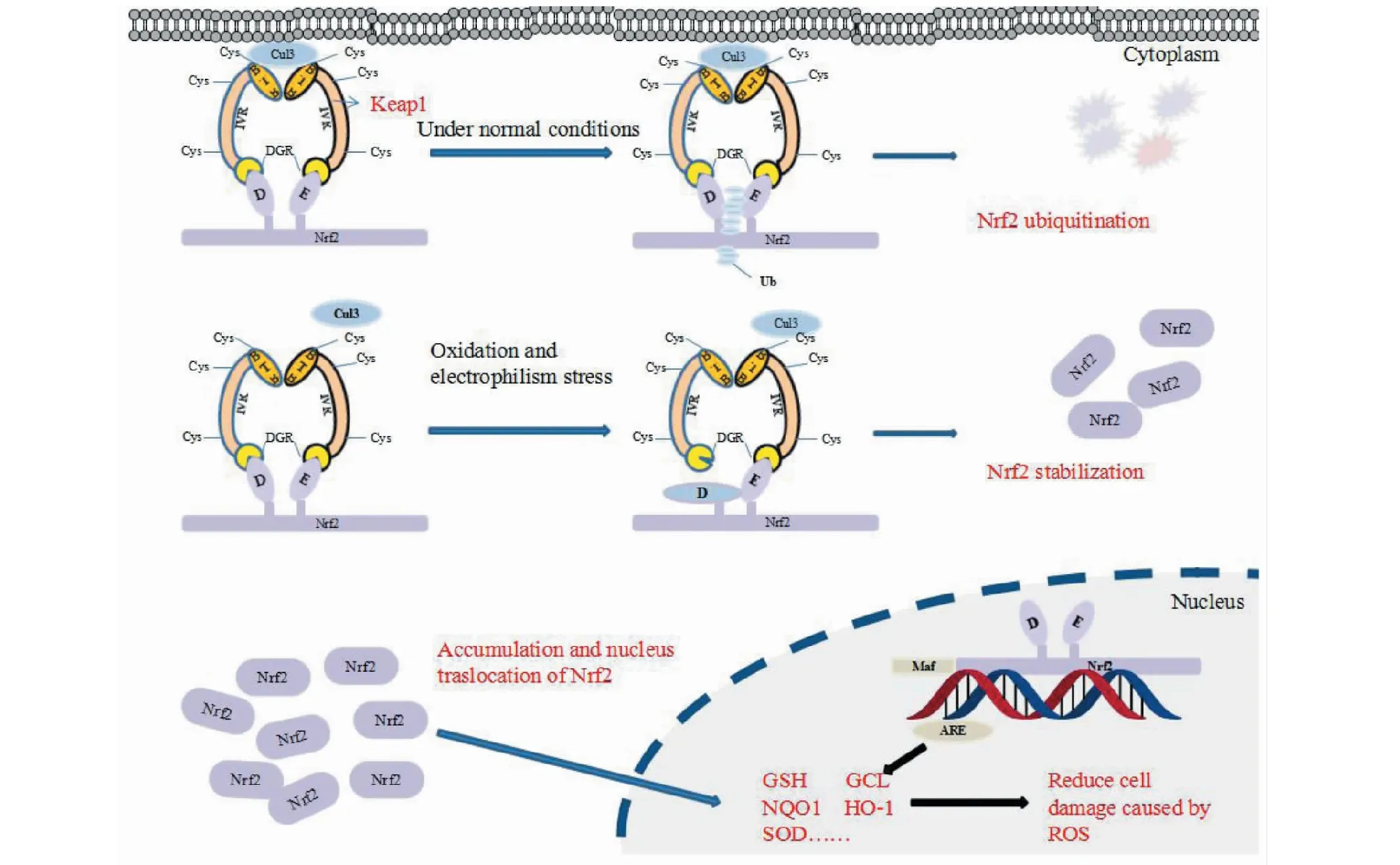
图1 Keap1-Nrf2-ARE 信号通路Fig.1 Keap1-Nrf2-ARE signaling pathway注:D为DLG基序,E为ETGE基序。正常情况下,Keap1与胞浆内的Nrf2相互作用,致使Nrf2泛素化,使Nrf2维持在一个相对较低的水平;在氧化应激条件下,Keap1的半胱氨酸残基被共价修饰,致使Keap1构象变化导致Nrf2释放并易位至细胞核以调节抗氧化酶的表达。Note:D is DLG motif,E is ETGE motif.Under normal physiological conditions,Nrf2 interacts with Keap1,followed by Nrf2 ubiquitination,maintaining Nrf2 at a relatively low level.Under oxidative stress,cysteine residues of Keap1 will be covalently modified.Subsequent conformational changes of Keap1 result in Nrf2 relieving and translocating into nucleus to regulate antioxidant enzymes.
除了上述的Keap1依赖的Nrf2-ARE信号通路调控模式外,Nrf2也能够通过非Keap1依赖的方式受到调控,如对于Nrf2的核转位和降解有着重要的贡献的磷酸化作用、伴侣蛋白结合、Nrf2半胱氨酸修饰等[16]。
2 基于天然产物及其衍生物的Nrf2激活剂
目前,绝大部分天然产物的Nrf2激活剂是Keap1-Nrf2相互作用间接抑制剂,它们通过同Keap1上的巯基官能团反应,修饰半胱氨酸残基,导致Keap1结构发生变化,Keap1-Nrf2复合物相互作用被破坏,Nrf2被激活。基于激活剂的结构以及同半胱氨酸反应的类型,Keap1-Nrf2相互作用间接抑制剂被分成迈克尔加成受体、可氧化双酚类、异硫氰酸酯类、二硫杂环戊烯硫酮及二烯丙基硫类、多烯化合物等类型。
2.1 迈克尔加成受体
含有α,β-不饱和羰基的迈克尔加成受体是最为常见的Nrf2激活剂,它们被认为是温和的路易斯酸,能够同Keap1中的半胱氨酸残基(温和的路易斯碱)发生迈克尔加成反应(图2),致使Keap1构象发生变化,阻止了Nrf2的泛素化,进而诱导Ⅱ相解毒酶及抗氧化蛋白的表达。常见的迈克尔加成受体有:姜黄素、黄酮类化合物、香豆素、肉桂酸衍生物、萜类化合物、生物碱类化合物等。
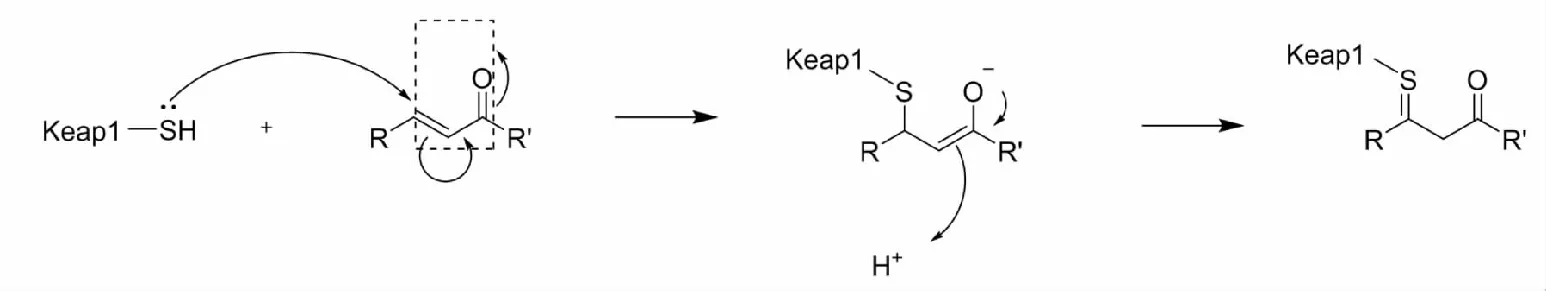
图2 迈克尔加成受体的作用机制Fig.2 Mechanism of action of Michael addition receptor
2.1.1 姜黄素及其衍生物
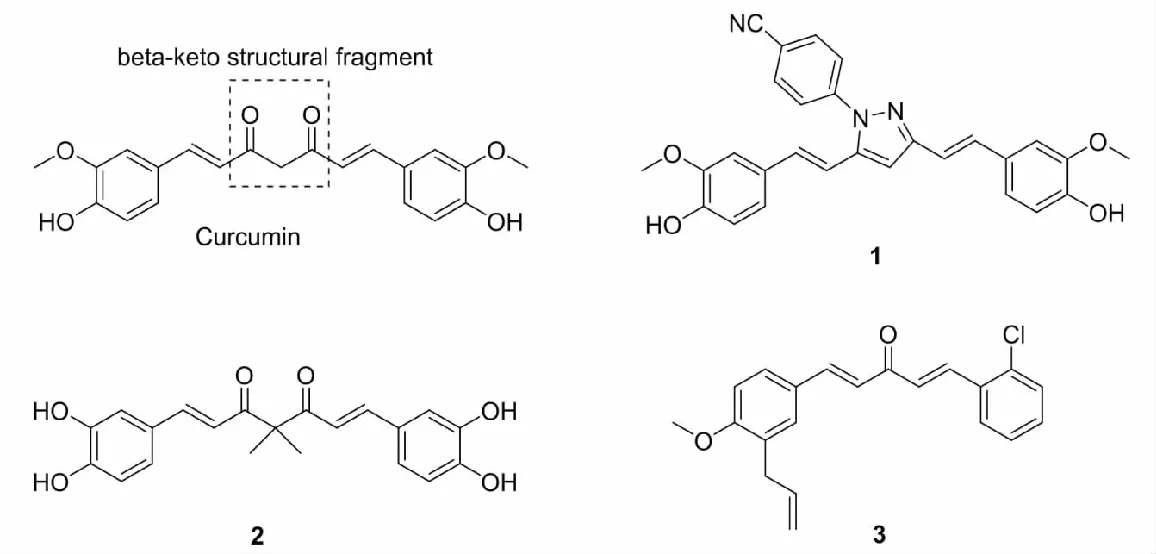
姜黄素(curcumin)是从姜黄根茎中提取的黄色膳食素,被广泛用于食品香料以及着色剂中,是一类具有代表性的抗氧化剂,近年来又被运用于神经退行性疾病中。研究发现,姜黄素含有α,β-不饱和羰基,属于典型的迈克尔加成受体,能够与半胱氨酸残基反应,使Keap1结构发生变化,进而激活Keap1-Nrf2-ARE信号通路,诱导一系列具有抗氧化功能的Ⅱ相解毒酶的表达,发挥细胞保护作用[17]。但是,姜黄素分子结构中存在活性亚甲基,致使其在生理条件下稳定性较差,生物利用度极低,严重制约了其在临床上的应用[18]。为克服这一局限性,出现了许多致力于修饰姜黄素分子结构的研究。
Liao等[19]通过对β-酮结构片段的修饰,设计合成了六种姜黄素吡唑衍生物,通过测试细胞活力和乳酸脱氢酶(lactate dehydrogenase,LDH)释放,评估了这六种化合物对硝普钠(SNP)诱导的PC12细胞损伤的神经保护作用,筛选出最佳化合物1,并验证得出1的神经保护作用是通过激活Keap1-Nrf2-ARE信号通路实现的。
Tu等[20]通过在姜黄素活性亚甲基上引入双甲基,改变姜黄素苯环上的取代基,设计合成了一系列姜黄素衍生物,通过评估这一系列化合物的稳定性以及对叔丁基过氧化氢(t-BHP)诱导的HepG2细胞氧化损伤的修复能力,筛选出最佳化合物2,并发现2能够与机体内的Fe2+和Cu2+反应,产生相应的醌类结构,从而激活Keap1-Nrf2-ARE信号通路,增加Ⅱ相解毒酶的表达。
Li等[21]通过修饰姜黄素β-二酮部分合成了一系列的单羰基姜黄素类似物,通过测定细胞活力以及丙二醛(malondialdehyde,MDA)、H2O2和SOD的表达水平,评估了这些化合物在H2O2或t-BHP诱导的H9c2细胞氧化损伤的修复能力,筛选出最佳化合物3,并发现3能够促进HO-1和SOD的表达,在一定程度上阻止小鼠心肌细胞的损伤。
2.1.2 黄酮类化合物
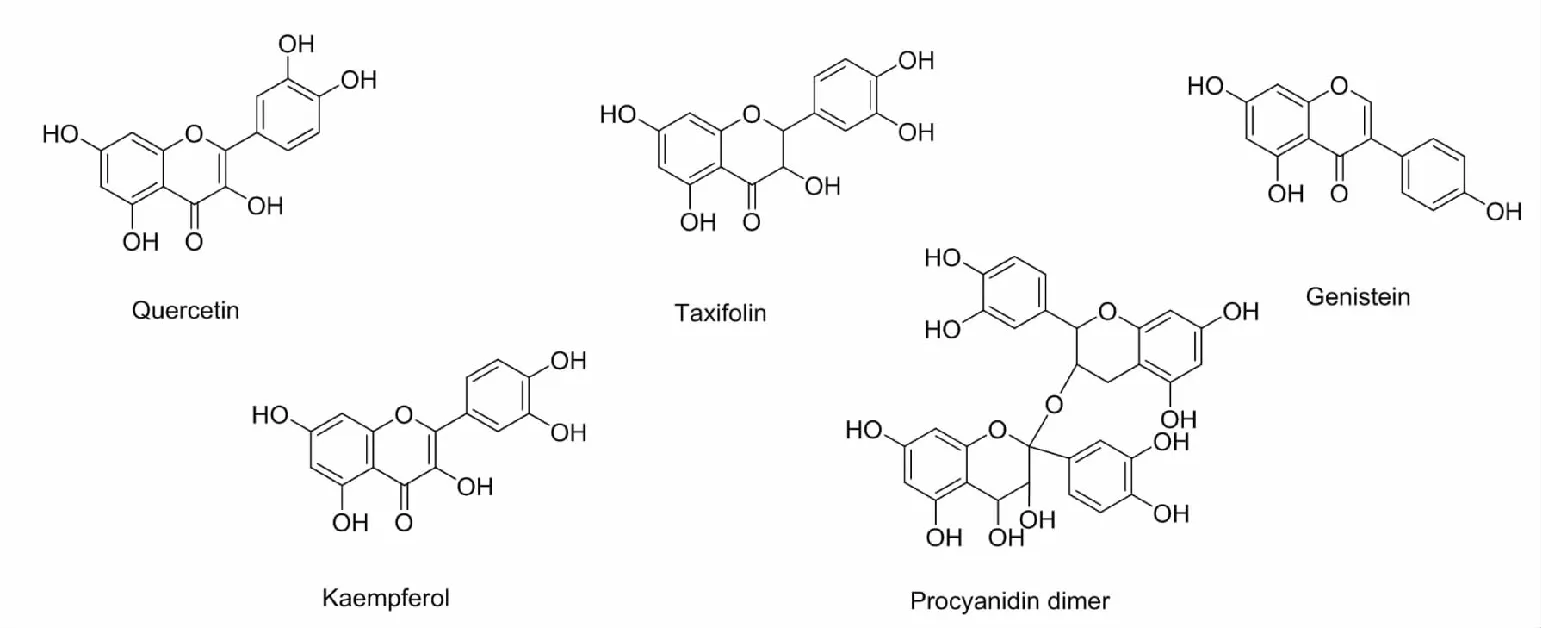
黄酮(flavonoid)类化合物广泛存在于某些植物和浆果中,其母核为2-苯基色原酮,包括黄酮、二氢黄酮、异黄酮、二氢异黄酮、查尔酮等,是一类强力的抗氧化剂,能够有效的清除体内的自由基,阻碍细胞的退化和衰老。随着研究的深入,人们发现槲皮素(quercetin)、二氢槲皮素(taxifolin)、山奈酚(kaempferol)、金雀异黄素(genistein)、原花青素(procyanidins)等天然黄酮类化合物能够通过激活Nrf2-Keap-ARE信号通路来抵御机体内氧化应激损伤。
Li等[22]为寻找新型黄酮类Nrf2激活剂,对黄酮类化合物进行了系统研究,他们从植物中提取出了74种黄酮类化合物,通过NAD(P)H:醌还原酶(QR)分析法检测此类化合物在Hepa1c1c7小鼠肝癌细胞中的Nrf2激活活性,筛选出活性最佳化合物4(25 μM下NAD(P)H:QR = 2.08),并发现4通过丝裂原活化蛋白激酶(MAPK)、蛋白激酶C(PKC)和蛋白激酶R样内质网激酶(PERK)引起Nrf2磷酸化,从而激活Nrf2,增加细胞的抗氧化活性。
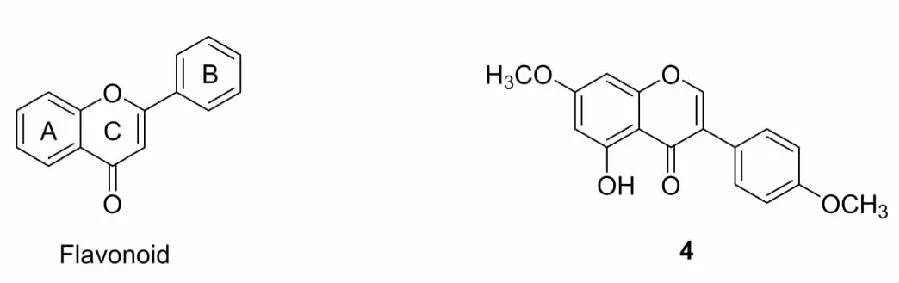
查尔酮(chalcone)是植物体内合成黄酮的前体,是一类具有的抗虫、抗炎、抗肿瘤、抗氧化、抗真菌等药理活性的黄酮类化合物。研究发现,查尔酮的Nrf2激活活性比其他黄酮类化合物更显著。因此,出现了许多研究致力于修饰查尔酮结构来设计更出色的Nrf2激活剂。
首先,Kumar和Lee等[23,24]发现在查尔酮B环上引入三氟甲基(尤其邻位取代)或用磺酰基代替查尔酮的α,β-不饱和羰基中的羰基能显著增加查尔酮的Nrf2激活活性,据此,他们合成了查尔酮衍生物5(诱导GCLM1的表达量为萝卜硫烷的4.5倍)和6(IC50= 1.52 μM)。随即,三氟甲基和磺酰基取代的查尔酮衍生物引起了研究者的广泛关注,Lounsbury等[25]在Kumar等研究的基础上,将B环的苯环改变为嘧啶环,设计了化合物7,发现经7处理的BEAS2-2B(人正常肺上皮细胞)具有更高含量的HO-1(表达量为5的3倍)。Kim等[26]研究阿尔兹海默病时,在A环邻位引入三氟甲基,在B环的4-引入1-丙酰基哌嗪,合成了生物利用度大、细胞膜穿透性好的查尔酮衍生物8,8能够在mRNA和蛋白质水平上激活Nrf2并诱导HO-1和GCLC的表达,还可以抑制BV-2小胶质细胞中一氧化氮的产生并下调了炎症介质,此外,他们还发现8能够恢复东莨菪碱诱导的小鼠模型的学习和记忆功能。Choi等[27]研究帕金森病过程中,在6的基础上引入了改善药物性质的氮杂环,合成了化合物9,不仅克服了6存在的溶解度低、代谢稳定性差、抑制细胞色素P(CYP)和对hERG(KCNH2基因)产生安全性隐患等缺陷,还提高了化合物的Nrf2激活活性(6:EC50= 530 nM;9:EC50= 346 nM),他们还发现9能够减轻1-甲基-4-苯基-1,2,3,6-四氢吡啶(MPTP)诱发的帕金森病小鼠模型中内多巴胺神经元的损伤,减缓帕金森病相关的运动障碍。除上述的三氟甲基和磺酰基外,研究还发现甲氧基也能够增强查尔酮的Nrf2激活活性,如Cui等[28]通过在查尔酮上引入甲氧基合成了化合物10,发现10能够保护视网膜色素上皮细胞免受氧化应激诱导的细胞死亡和线粒体功能障碍。
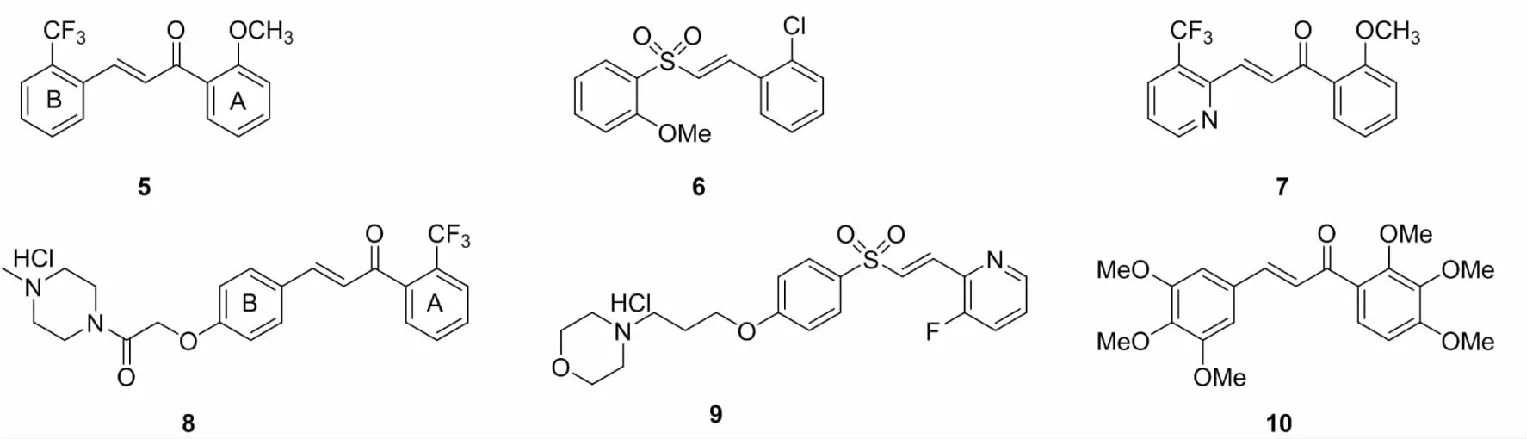
黄腐酚(xanthohumol)是由啤酒花的蛇麻腺分泌产生的异戊二烯基查尔酮类物质,具有预防癌症、糖尿病,抗菌抗病毒等生理活性。Li等[29]在研究黄腐酚对顺铂诱导肾毒性的保护作用和机制时,发现黄腐酚能够抑制肾脏组织中肿瘤坏死因子α(TNF-α),白介素-1β(IL-1β)和白介素-6(IL-6)的水平,抑制顺铂引起的GSH和SOD的活性下降以及NF-κB的活化,剂量依赖性上调Nrf2和HO-1的表达,从而减轻顺铂的肾毒性。Bai等[30]设计并合成了一批黄腐酚类似物,通过测试化合物对H2O2和6-羟基多巴胺(6-OHDA)诱导的PC12细胞氧化损伤的修复能力,筛选出最佳化合物11和12,分子机制研究表明,化合物11和12通过促进Nrf2的核积累从而增强细胞抗氧化防御系统,是有效的Nrf2激活剂。
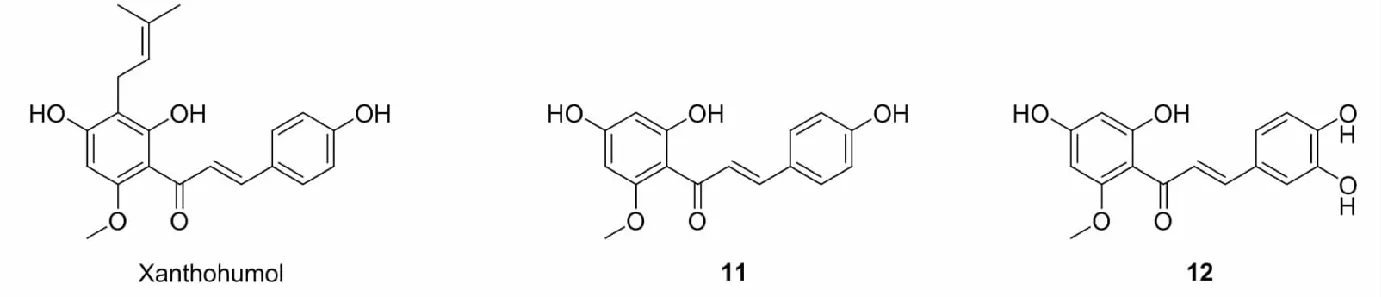
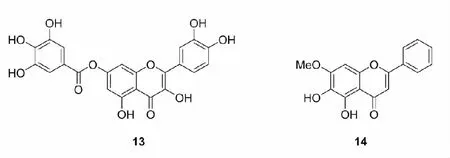
黄酮醇类(flavonols)是数量最多、分布最广泛的一类黄酮类化合物,具有抗过敏、抗炎、抗癌等药理活性。Roubalová等[31]合成了良好Nrf2激活活性的黄酮醇类化合物13,13能够激活AREc32细胞中的ARE,增加RAW264.7和Hepa1c1c7细胞中Nrf2的积累,诱导RAW264.7细胞的HO-1的表达以及增加Hepa1c1c7细胞中NQO1、HO-1和GCLM水平。Jing等[32]在研究了黄芩素-7-甲醚(14)对高原缺氧小鼠脑组织的保护作用及机制时,发现经14预处理后能够显著降低高原缺氧小鼠脑组织中H2O2、MDA和NO的含量以及LDH的活性,提高抗氧化酶的活性,同时进一步研究表明14能够增加Nrf2和HO-1的表达,进而缓解高原缺氧导致脑组织氧化应激损伤。
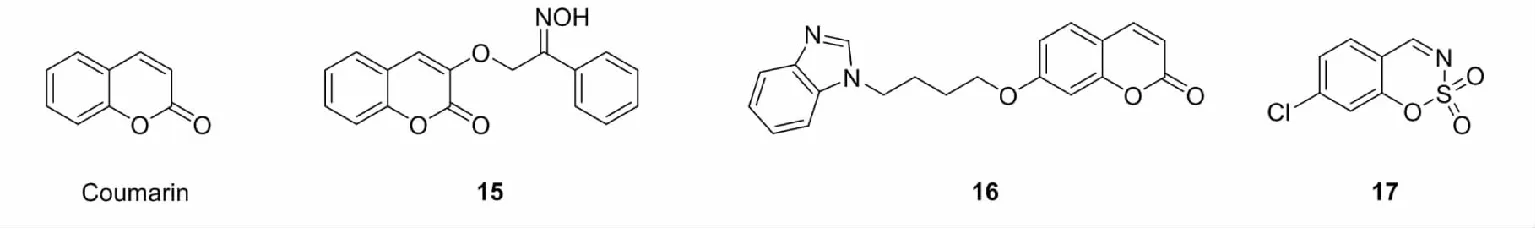
2.1.3 香豆素类
香豆素(coumarin)是存在于黑香豆、香蛇鞭菊等植物的一类化合物,具有抗肿瘤、抗菌、抗真菌、抗凝血作用等活性。香豆素母核为苯并呋喃酮,含有α,β-不饱和羰基,能够与半胱氨酸残基反应,导致Keap1结构发生变化,进而激活Nrf2。
Chang等[33]通过在香豆素呋喃酮部分引入取代基,设计合成了一系列含肟或酰胺的香豆素衍生物,筛选出Nrf2激活活性最好的化合物15(EC50= 3.45 ± 0.25 μM)。Liu等[34]通过在香豆素苯环上引入取代基,设计合成了一类羟基香豆素16,16通过提高PKC磷酸化水平,破坏Nrf2-Keap1相互作用,进而激活Nrf2,诱导HO-1的表达。Wei等[35]受到Lee和Choi等的启发,将香豆素的呋喃酮部分结构优化,设计了一系列香豆素衍生的亚氨基磺酸类化合物,通过测试化合物对H2O2诱导的H9c2细胞氧化损伤的修复能力,筛选出最佳化合物17,17能够诱导Nrf2的核转位,上调内源性抗氧化蛋白HO-1和NQO1的表达,保护细胞免受氧化损伤。
2.1.4 肉桂酸类
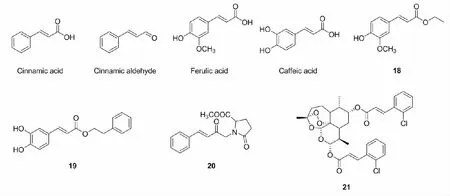
肉桂酸(cinnamic acid)是肉桂皮或安息香分离出的有机酸,结构为苯并丙烯酸,与之结构类似还有肉桂醛(cinnamic aldehyde)、阿魏酸(ferulic acid)、咖啡酸(caffeic acid)等。其中,阿魏酸乙酯(18)和咖啡酸苯乙酯(19)均被证实具有抗氧化作用[36,37]。此外,研究人员发现或合成了许多含有Nrf2激活活性的苯并丙烯酸类化合物。
2008年,Wondrak等[38]在研究皮肤光老化时设计合成了一系列肉桂酰基化合物,通过荧光素酶报告基因检测发现肉桂醛和1-肉桂酰基-5-氧代-2-吡咯烷甲酸甲酯(20)具有Nrf2激活活性,能够诱导抗氧化蛋白HO-1和NAD(P)H的表达,抑制细胞内氧化应激,阻碍皮肤光老化。Nicholas等[39]在研究动脉粥样硬化时,也发现肉桂醛能够激活Nrf2,此外还发现肉桂醛能够抑制肥胖Ⅱ型(ZDF)大鼠中球囊损伤后的新内膜增生。Li等[40]近期在研究骨关节炎时,通过在二氢青蒿素上引入苯并丙烯酸类结构设计了一类具有药理活性的青蒿素衍生物21,并指出其作用机制是通过调节Nrf2和NF-κB信号通路,减弱骨关节炎滑膜的炎症反应。
2.1.5 萜类
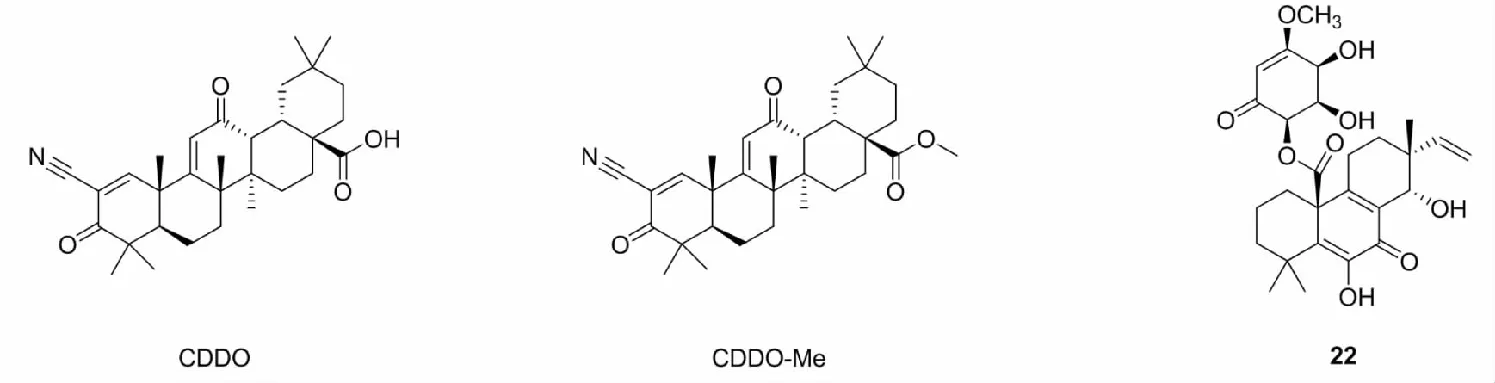
萜类是分子式为异戊二烯的整数倍的烯烃类化合物,部分含有α,β不饱和羰基或者共轭多烯长链的结构具有Nrf2激活作用。2005年,Couch等[41]通过齐墩果酸合成了一种抗炎效果的三萜类化合物CDDO,实验还发现,CDDO能够与Cys151残基发生共价反应,是一类潜在的Nrf2激活剂。类似的化合物还有CDDO-Me(也被称为RTA402),临床前研究表明,CDDO-Me能够激活Nrf2和抑制NF-κB,但是,在一项关于CDDO-Me治疗慢性肾脏病的Ⅲ期临床试验中,研究人员发现其在心脏方面存在安全性隐患,致使试验宣告失败[42]。此外,Wang等[43]于2017年在真菌Botrysphaeria中提取了一种具有明显的QR活性二萜类化合物22,还证实了22通过活化激酶PERK、PI3K、PKC和MEK1/2,激活Nrf2,增加NQO1和γ-GCS的表达,从而保护细胞免受As(Ⅲ)的诱导损伤[44]。
2.1.6 生物碱类
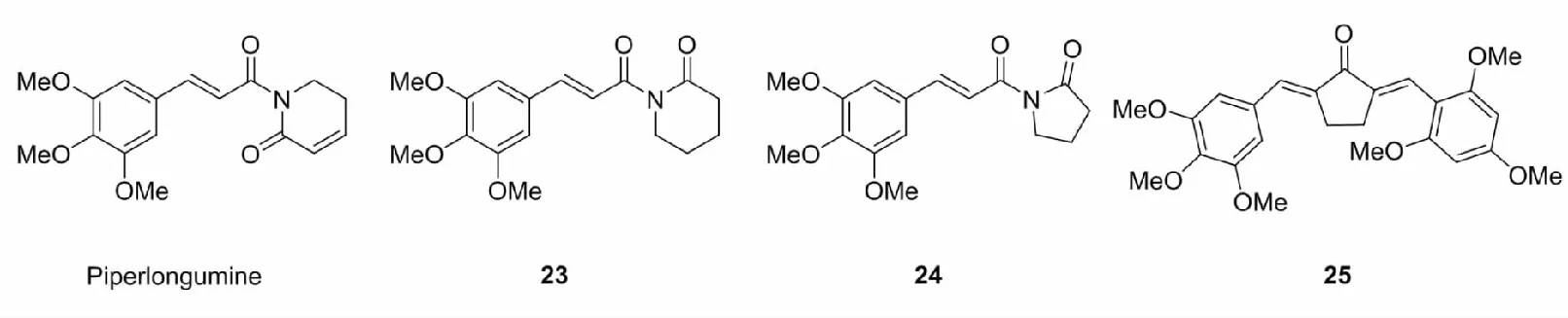
荜拔酰胺(piperlongumine)是天然植物荜拔提取出的一类生物碱,具有抗动脉粥样硬化、镇痛、抗炎、抗真菌、抗肿瘤等活性。荜拔酰胺是一种查尔酮类似物,含有α,β-不饱和羰基,具有潜在的Nrf2激活活性。Peng等[45]合成了一系列荜拔酰胺类似物,筛选出细胞毒性最小的化合物23和24,研究发现,23和24能够促进Nrf2核易位,上调一系列细胞抗氧化因子的表达,从而保护PC12细胞免受氧化损伤。Li等[46]在研究脑缺血再灌注损伤时,将筛选出的具有抗氧化作用、低毒性的二酮骨架与3,4,5-三甲氧基苯基杂化,设计了一系列新颖的荜拔酰胺类似物,其中活性最佳的化合物25是有效的Nrf2激活剂,具有更低的细胞毒性,能够激活Nrf2,减轻体内脑损伤。
胡椒碱(piperine)是胡椒中提取出来的一种桂皮酞胺类生物碱,具有抗氧化、抗肿瘤、抗寄生虫、免疫调节等生物活性。此外,Choi等[47]发现胡椒碱,能够剂量依赖性的增加HO-1的表达,激活Keap1-Nrf2和MAPK信号通路,从而保护细胞免受顺铂诱导的细胞凋亡。但是由于其生物利用度低,作用机制不明确,严重制约了其在临床上的运用。为了提高胡椒碱的生物利用度,研究人员开发了大量的胡椒碱类似物。例如,Wang等[48]在研究帕金森综合症时合成了一系列胡椒碱类似物,通过评估化合物对H2O2诱导的神经样PC12细胞的氧化损伤修复能力,筛选出活性最佳化合物26,化合物26的ROS清除和细胞保护可能与Nrf2的活化,HO-1、NQO1等Ⅱ相解毒酶表达的上调有关,进一步研究还发现,化合物26能够减轻MPTP诱发的帕金森病小鼠模型的行为障碍,保护酪氨酸羟化酶免疫阳性的多巴胺能神经元。Yang等[49]在研究阿尔兹海默病时,合成了一类新型胡椒碱衍生物27,27在体外具有很强的激活Keap1-Nrf2-ARE信号通路的能力,该团队还发现27通过直接抑制了Keap1和Nrf2之间的相互作用激活Nrf2,从而抑制了氧化应激和硫氧还蛋白互作蛋白(TXNIP)介导的Nod样受体蛋白3(NLRP3)炎性小体的激活。
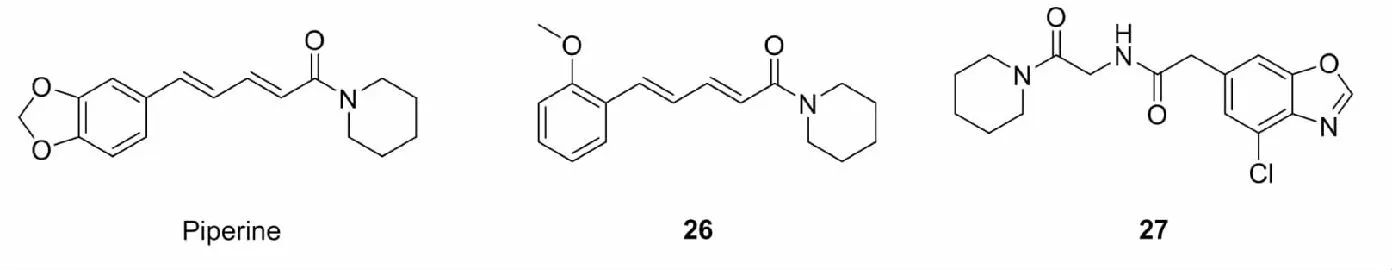
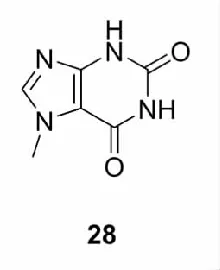
此外,研究人员还发现了其他生物碱类Nrf2激活剂。如Endesfelder等[50]研究发现咖啡因(28)能激活大脑内Nrf2,增加Ⅱ相解毒酶的表达,从而减少因高氧血症引起的新生儿脑神经损伤;Shu等[51]在探究荷叶总生物碱(TAL)对醋氨酚(APAP)所致小鼠急性肝损伤的保护作用机制时,发现在TAL的作用下,肝内AMPK磷酸化水平提高,导致Nrf2蛋白入核,HO-1和GCLC基因表达上调,从而降低血清中的丙氨酸氨基转移酶(ALT)和天门冬氨酸氨基转移酶(AST)活性,下调肝组织中TNF-α、IL-1β、IL-6和MDA含量,上调肝组织中SOD、CAT、谷胱甘肽过氧化物酶(GSH-Px)和GSH的水平,最终保护APAP诱导的急性肝损伤。
2.2 可氧化双酚类
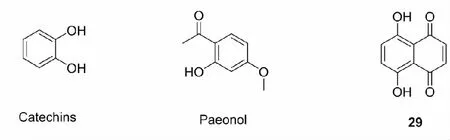
多羟基酚类化合物广泛存在于自然界中,部分多羟基酚类化合物能够在体内氧化成具有亲电活性的醌类化合物(图3),与半胱氨酸的巯基反应,使Keap1构象发生变化,导致Nrf2易位至细胞核中,与ARE结合并诱导下游Ⅱ相解毒酶的表达,保护细胞抵御氧化应激[52]。常见的可氧化双酚Nrf2激活剂有儿茶酚(catechins)[53]、丹皮酚(paeonol)[54]、白藜芦醇(resveratrol)等。此外,Son等[55]还于2013年发现一种能够诱导内源性抗氧化反应原件在神经元和星形胶质细胞中表达的天然化合物5,7-二羟基-1,4-萘醌(29)。

图3 多羟基酚类的作用机制
白藜芦醇是一类具有抗氧化,强化血管壁、促进肠胃消化、降低血脂肪、增加身体抵抗力、抗辐射的多羟基酚类化合物,最初是由日本学者在毛藜芦的根茎中发现,研究发现,白藜芦醇能与Keap1半胱氨酸的硫醇基团反应,增加Nrf2在细胞核的释放,增加机体内的Ⅱ相解毒酶的表达[56],但是白藜芦醇溶解性差,生物半衰期短,造成白藜芦醇在此方面的研究受限,为克服这一局限性,出现了许多致力于白藜芦醇结构修饰的研究。
2015年,Lin等[2]将白藜芦醇的3,5,4' -三羟基易位,设计了六种白藜芦醇羟基化类似物,以t-BHP诱导的HepG2细胞作为氧化损伤模型,检测这六种化合物的细胞保护活性,筛选出了最佳化合物30,此外,还发现30利用细胞内的铜离子构建一个有效的促氧化体系,产生对应的邻醌,导致蛋白激酶B(Akt)的磷酸化,抑制Nrf2的泛素化,从而激活Nrf2。同年,Li等[57]使用化学或酶促方法合成了三种均具有Nrf2激活活性白藜芦醇二聚体31~33,但是它们活性均比白藜芦醇单体低。
2.3 异硫氰酸酯类
天然的异硫氰酸酯类化合物主要以硫苷形式存在于十字花科植物当中,具有抗肿瘤、抗氧化等活性。异硫氰酸酯类化合物含有典型的官能团结构(-N=C=S),该官能团中碳原子受到双键两侧的N和S原子的吸电子诱导效应,致使C原子具有很强的亲电性,导致异硫氰酸酯能够与半胱氨酸残基的巯基反应生成二硫代氨基甲酸酯(图4),造成Keap1构型变化,从而激活Nrf2[58]。
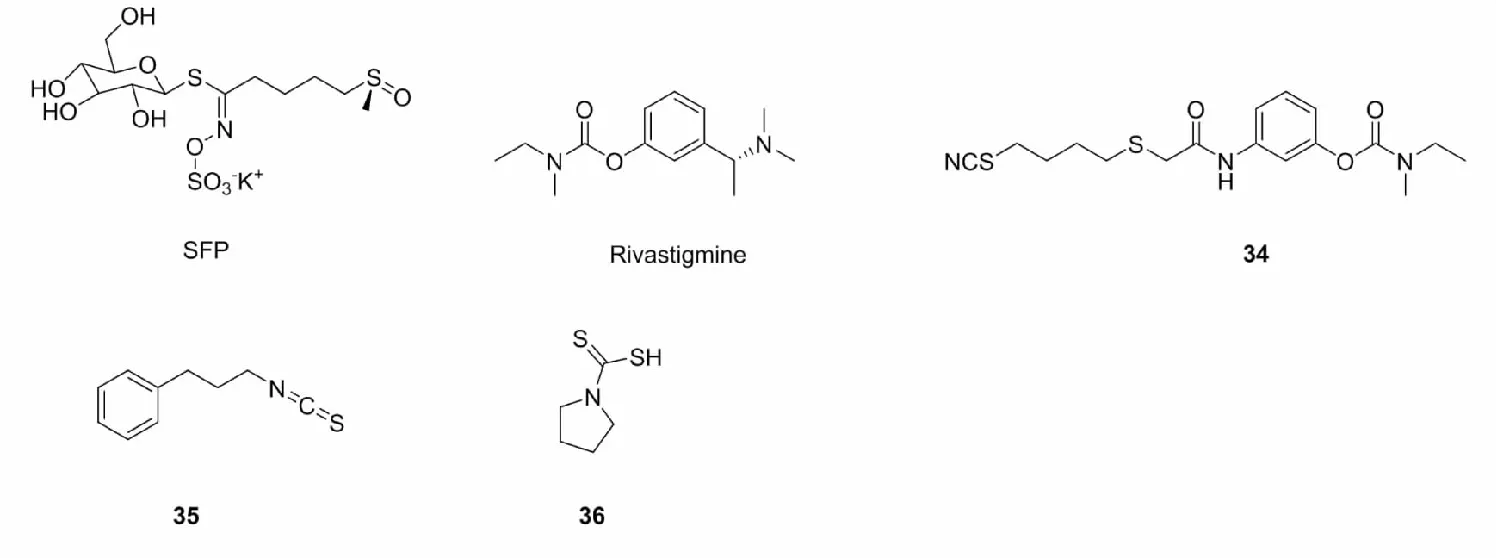
萝卜硫烷(SFP)又称“莱菔硫烷”,典型的异硫氰酸盐。在动物的肾脏组织中,SFP能够上调Nrf2和HO-1基因的表达,下调IL-6和caspase-3基因的表达[59]。Sestito等[60]在研究H2S供体治疗阿尔兹海默病的过程中,将萝卜硫烷和芥氨酸与利伐斯的明(rivastigmine)杂合,合成了六种具有抗炎抗氧化活性的化合物,其中34在5μM下具有Nrf2激活活性。
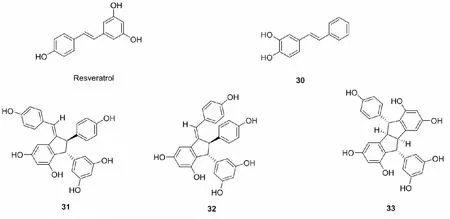
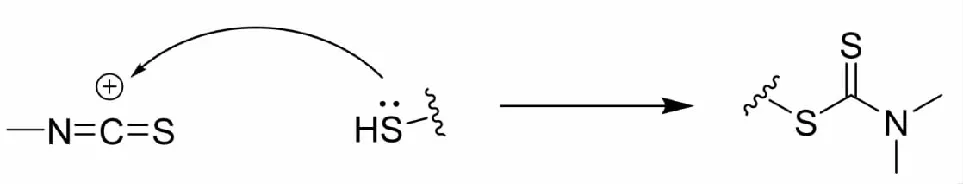
图4 异硫氰酸酯类的作用机制Fig.4 Mechanism of action of isothiocyanates
苯乙基异硫氰酸酯(35)也是自然界中存在的常见异硫氰酸酯类化合物,2013年,Krajka-Kuzniak等[61]评估比较了异硫氰酸苯乙酯、吲哚-3-甲醇、黄腐酚和白藜芦醇及其两两组合对人胰腺癌细胞中的NF-κB和Nrf2表达的影响,筛选出Nrf2激活活性最大的苯乙基异硫氰酸酯与黄腐酚组合,发现苯乙基异硫氰酸酯与黄腐酚组合能够增强谷胱甘肽转移酶(GSTP)、NQO1和SOD的表达,保护细胞免受氧化应激损伤。
此外,Liddell等[62]发现一类具有Nrf2激活活性的二硫代氨基甲酸酯类化合物36,36能诱导Ⅱ相解毒酶的表达,激活Nrf2并抑制NF-кB。
2.4 二硫杂环戊烯硫酮及二烯丙基硫类
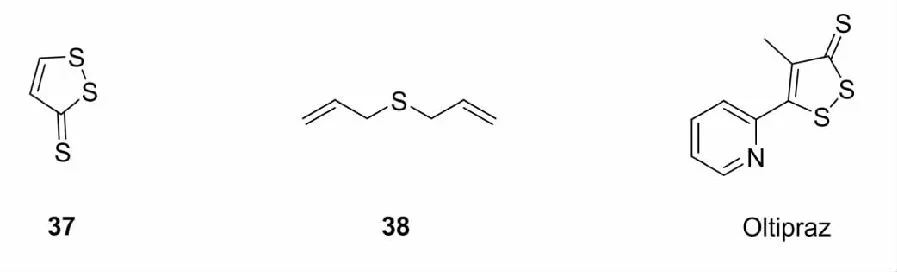
二硫杂环戊烯硫酮是对抗癌物质具有细胞保护作用的含硫五元环状化合物,烯丙基硫醚类化合物是一类能够有效抑制多种癌细胞的有机硫化合物,研究发现,二硫杂环戊烯硫酮和二烯丙基硫类化合物中的二硫键能够与半胱氨酸的巯基发生巯基-二硫键交换反应[63],修饰Keap1,抑制Nrf2的泛素化,从而上调Ⅱ相解毒酶的表达。常见的此类Nrf2激活剂有3H-1,2-二硫杂环戊烯-3-硫酮(37)和二烯丙基硫醚(38)[64]。此外,研究还发现FDA批准的最早用于治疗抗血吸虫病的药物oltipraz也能够增加GSH等Ⅱ相解毒酶的表达,清除机体内的ROS[65]。
2.5 多烯类化合物
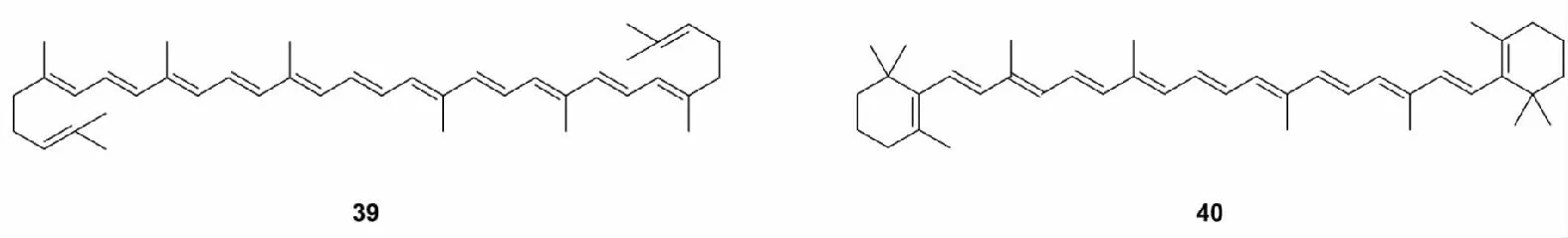
多烯类化合物是具有多个单双键交替的高度不饱和有机物,此类化合物在机体内较易转化成亲电代谢产物,能够同半胱氨酸残基的巯基发生反应,激活Nrf2。比较典型的有番茄红素(39)和β-胡萝卜素(40)[66]。
2.6 其他天然产物
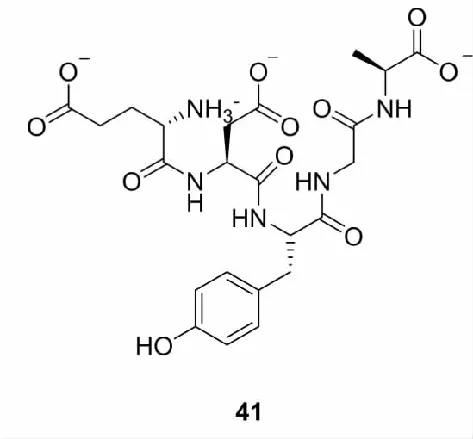
除了上述的天然产物外,研究还发现其他结构类型的天然Nrf2激活剂。如多肽类化合物,Wang等[67]从甲鱼蛋白中提取出了7个抗氧化多肽,通过ARE-荧光素酶报告基因检测筛选出Nrf2激活活性最佳的多肽41。
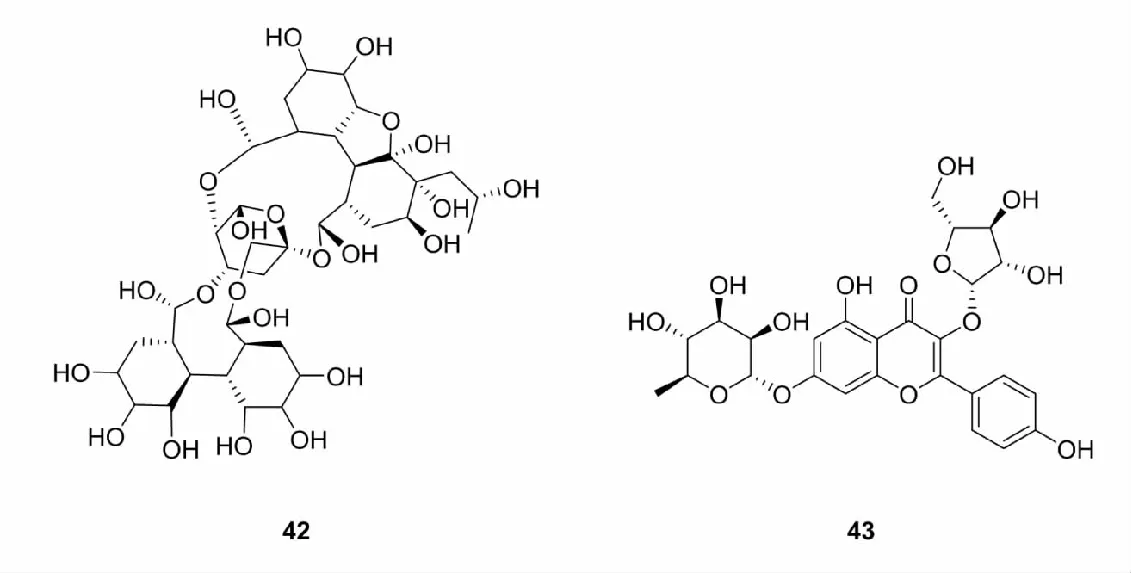
除上述比较常见的天然Nrf2激活剂外,自然界中还存在硒代化合物、过氧化物、含有离去基团的亲电试剂的天然Nrf2激活剂,甚至发现三价砷化合物、重金属离子及金属复合物等无机化合物也可以激活Keap-Nrf2-ARE信号通路。此外,Nrf2-Keap1蛋白-蛋白结合抑制剂(protein-protein interaction inhibitors,PPI)越来越受到研究人员的广泛关注[68,69],PPI是Keap1-Nrf2相互作用直接抑制剂,能够直接干扰Nrf2-Keap1蛋白相互作用,致使二者解离,从而激活Nrf2发挥抗氧化防御作用[70],目前,已有部分基于天然产物结构的PPI被报道,如Zhang等[71]从芸香科吴茱萸中提取出的芸香碱(rutaecarpine),Martiniano等[72]提取的乙酰基香叶素(42)和山奈酚糖苷类化合物(43)。
3 讨论和展望
Nrf2-Keap1-ARE信号通路是细胞抵御氧化应激的重要机制,能够诱导机体自身产生的一系列Ⅱ相解毒酶抵御氧化应激。与一般抗氧化剂中和自由基不同,提高Ⅱ相解毒酶的活性可以形成一个持续的抗氧化过程,作用时间长且效果佳,据此,该信号通路已成为许多疾病的靶标[73]。天然产物是生物与大自然长时间“斗争”下的“杰作”,具有结构多样、活性多样等特点,是药物设计者的灵感源泉[3]。此外,许多天然产物具有亲电性,可以修饰蛋白分子的亲核部分(尤其是活性半胱氨酸残基部位),因此许多天然产物可以使Keap1结构发生变化。在此基础上,“基于天然产物的结构修饰开发出更佳的Nrf2激活剂”已成为热门研究策略。但是,随着研究的进展,人们发现Keap1结构修饰剂与半胱氨酸残基发生反应常常是不可逆的,长期作用使得Nrf2异常持续活跃有致癌变可能,此外,Nrf2过度激活还会促进癌细胞的增殖[74],因此,在今后天然产物结构优化中,找到更合适的Nrf2激活剂尤为重要。
本文介绍了Keap1-Nrf2信号通路结构及其作用机制,总结了近年来常见的天然产物导向的Nrf2激活剂,相信随着提取技术和现代生物合成技术的不断进步,越来越多的高活性的Nrf2激活剂将会被合成或者被发现,为今后治疗某些疾病提供一种新的策略。
