湘云金鳙染色体组型分析
2021-01-30杨春英刘良国刘飞周工健李游
杨春英, 刘良国, 刘飞, 周工健, 李游
湘云金鳙染色体组型分析
杨春英1, 刘良国2, 刘飞3, 周工健4, 李游3
(1. 湖南文理学院 师范学院, 湖南 常德, 415000; 2. 湖南文理学院 教务处, 湖南 常德, 415000; 3. 动物学湖南省高校重点实验室, 湖南 常德, 415000; 4. 湖南湘云生物科技有限公司, 湖南 常德, 415000)
采用PHA和秋水仙素活体注射法, 对洞庭湖区人工养殖湘云金鳙的染色体组型进行分析。结果表明: 湘云金鳙的染色体二倍体数为2N= 48, 核型公式为: 24 m+18 sm+6 st, NF = 90。未发现端部着丝粒染色体, 也未发现带有特殊标志性特征如随体和次缢痕等的染色体。同时, 本文将洞庭湖区与其他水域鲢亚科鱼的核型进行比较, 探讨了鲢亚科鱼类的染色体遗传多样性。
洞庭湖区; 湘云金鳙; 染色体组型
鳙()隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲢亚科(Hypophthal- michthyinae), 又称花鲢、胖头鱼[1]。鳙具有生长快, 个体大, 肉质鲜美, 营养价值高, 抗病力强等优点, 是中国特有淡水鱼类资源, 也是我国大水面水体中的主要养殖对象, 在我国淡水养殖总产量中位居第四[2]。国内对鳙的研究始于19世纪50年代, 主要集中在鳙的人工繁养殖[3]、病害防治[4]、年龄生长和食性[5]、生理[6]、遗传多样性[7]、深加工及综合利用[8]等方面。20世纪70–80年代, 周暾[9]、刘凌云[10]、昝瑞光[11]、余先觉等[12]对鳙鱼的染色体组型进行过研究; 2006年, 孔庆亮等[13]开展了对江苏省铜山地区的鳙鱼的染色体DAPI核型分析。近年来, 由于人工繁殖缺乏科学监管, 鳙的种质资源出现退化[5], 生态环境的相对恶化又导致鳙鱼的天然资源在急剧的减少[15], 以上种种原因均导致鳙的遗传多样性水平下降。
湘云金鳙(red var.)是湖南湘云生物科技有限公司从天然发生基因突变的鳙鱼鱼种中, 经多年人工培育和系统选育, 获得的具有稳定遗传性状和优良养殖性状的新品系[16]。该品系由于生长速度较快、抗逆能力较强、味道鲜美、鱼体为桔红色, 深受消费者以及渔业养殖户的欢迎[17]。目前有关湘云金鳙的研究相对较少, 仅见严斌等[18]对湘云金鳙和普通鳙鱼进行了RAPD及微卫星分析; 孙远东[17]对湘云金鳙群体的生物学和遗传特性进行了研究; 刘飞等[19]对金鳙和黑鳙两种鱼的肌肉营养成分进行分析和营养价值进行评定; 杨福忠等[16, 20]对湘云金鳙的生物学特征及养殖特性进行了报道。关于湘云金鳙的染色体组型研究尚未见报道。本实验采用PHA和秋水仙素活体注射法, 对洞庭湖区人工养殖的湘云金鳙进行染色体组型分析, 以丰富鳙的遗传学基础数据, 为鳙的遗传选育提供理论参考, 同时也为鳙在鲤科鲢亚科鱼类中的系统演化研究提供数据参考。
1 材料与方法
1.1 实验材料
实验鱼湘云金鳙取自湖南湘云公司养殖基地, 从池塘中随机选取8尾(3雌5雄)用于染色体组型分析。实验鱼体长424~449 mm, 体重1 355~1 512 g。湘云金鳙运回后立刻放入室内水族箱(控温21~24 ℃)暂养1周后开始实验(图1)。
1.2 实验方法
1.2.1 肾细胞染色体标本的制备
制作湘云金鳙肾细胞染色体标本的方法参照刘良国等[21]的染色体标本制备方法并加以改进, 操作步骤如下: 实验前1 d对腹腔注射植物血球凝集素(PHA), 12 h后再次对腹腔注射PHA, 两次注射PHA溶液的剂量均为按鱼体重10 μg/g, 第2次注射PHA4~5 h后, 按鱼体重2 μg/g注射秋水仙素, 1.5~2.0 h后, 剪断鱼鳃或鱼尾失血10 min, 取出肾脏(头肾), 放入培养皿中, 用生理盐水(0.8% NaCl)清洗干净, 剔除附着的血块和脂肪组织; 用眼科剪剪碎肾脏, 吸入10 mL离心管中, 用吸管充分吹打(200次以上); 再吸入少量生理盐水, 混合均匀后静置5 min稍待沉淀, 吸取上层细胞悬液至另一离心管; 离心后的细胞沉淀低渗处理, 低渗液为0.075 mol/L KCl液, 卡诺氏液固定, 换液3次, 制成细胞悬液; 用滴管在距离干净冷冻玻片1 m处滴片, 酒精灯火焰干燥方法制片, Giemsa染液染色2 h后, 对玻片反面用微流水冲洗, 室温下晾干后, 获得湘云金鳙的肾细胞染色体玻片标本。

图1 实验用洞庭湖区湘云金鳙
1.2.2 染色体数目统计及组型分析
在光学显微镜下观察湘云金鳙的肾细胞染色体玻片标本, 选择100个形态比较清晰、分散较好的染色体中期分裂相, 油镜下用显微数码成像系统逐一进行拍照保存, 再利用Photoshop软件统计拍摄的每个分裂相的染色体数目, 统计湘云金鳙的染色体众数分布数据。确定其染色体数目后, 选取5个数目完整、分散良好、图像最清晰、染色体长度适中的中期分裂相, 用Image J软件测量其全长、短臂长和长臂长, 用Microsoft excel软件计算其相对长度和臂比, 得到湘云金鳙的核型数据, 依照Levan等[22]的染色体分组标准对湘云金鳙进行核型分析。

表1 洞庭湖区湘云金鳙染色体数目分布
2 实验结果
2.1 洞庭湖区湘云金鳙的染色体数目
统计8条湘云金鳙共100个数目完整、图像清晰的中期分裂相的结果(表1): 染色体数目在45及以下、47和49及以上的均为1个细胞, 染色体数为45的3个细胞, 为46的4个细胞, 为48的40个细胞。染色体数目为48的细胞占计数细胞总数的80%, 众数百分率高于75%的常规标准, 由此确定洞庭湖区人工养殖湘云金鳙的标准染色体数2N = 48(图2)。
2.2 洞庭湖区湘云金鳙的染色体组型
根据洞庭湖区人工养殖湘云金鳙染色体相对长度和臂比值的测量结果(表2), 将其染色体配成24对, 按Levan命名法, 湘云金鳙的核型由12对中部着丝粒染色体、9对近中部着丝粒染色体、3对近端部着丝粒染色体组成(图3), 没有端部着丝粒染色体, 核型公式为2N = 24 m+18 sm+6 st, NF = 90, 在湘云金鳙的雌雄个体之间未发现与性别有关的异型性染色体, 也未发现带有特殊标志性特征如随体和次缢痕等的染色体。

图2 洞庭湖区湘云金鳙的染色体中期分裂相(CN = 48) (×1 000)

表2 洞庭湖区湘云金鳙的染色体核型分析

图3 洞庭湖区湘云金鳙的染色体中期分裂相及组型(标尺=5 μm)
根据已报道的鳙的染色体组型(表3)可知, 湖北、云南、北京、江苏、湖南等地的鳙的染色体数目是一致的, 但在染色体组型上存在一定差异; 且湖南洞庭湖区的鳙与湖北武汉的鳙染色体组型差异较小, 而与北京和江苏的鳙染色体组型差异较大。

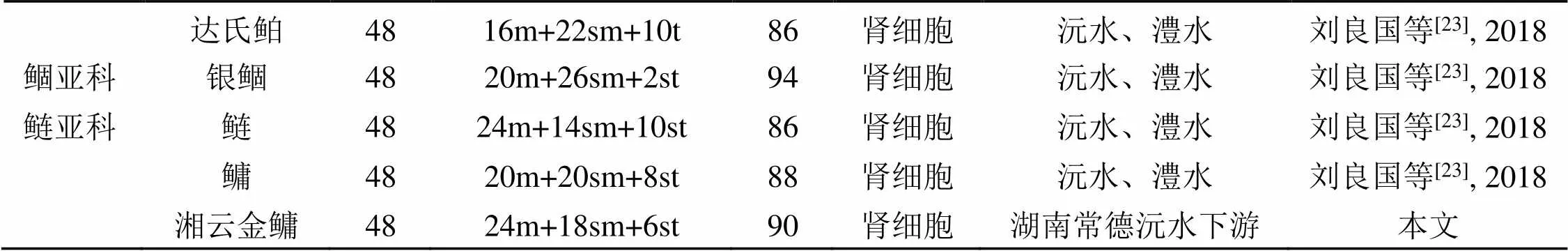
表3 不同水域鳙、鲢鱼的核型比较
注: 部分文献臂数算法跟本文采用的不一致, 现已全部换成余先觉所用的方法得出。
3 分析与讨论
染色体是遗传物质的主要载体, 鱼类染色体核型分析对研究鱼类的分类、系统演化、遗传变异、性别决定、杂交育种等具有重要意义。根据前人报道(表3)可知: 不同区域鳙的染色体二倍体数目相同, 而在染色体组型上存在差异, 表现出一定的多态性。究其原因, 有可能是由于鳙鱼群体所处水域不同, 环境差别大, 由于长期人工繁殖, 各个地区的鳙鱼无法与其他地区的鳙鱼进行基因的交流, 致使它们为适应不同的环境, 在染色体组型方面产生差异。有研究表明, 鳙、鲢不同种群间形态特征差异的大小与其地理距离成正相关[26]。据表3中不同区域鳙的染色体组型来分析, 北京和云南鳙的核型差异较大, NF值差别最大。湖南与湖北鳙的核型差异较小, NF值差别最小。该结果表明不同区域鳙染色体组型差异的大小与其地理距离成正相关, 这与它们在形态特征上的差异是一致的。
周暾[9]的研究显示湖北武汉东湖鳙的2N = 48, 核型公式为: 6 m+36 sm+6 st, NF = 90; 余先觉等[12]的研究认为湖北武汉鳙的2N = 48, 核型公式为: 18 m+22 sm+8 st, NF = 88; 周暾和余先觉两位学者在相近时间取相同地区的鳙鱼得出的核型也不完全相同; 两者相比, 余先觉所得鳙鱼的m染色体明显比周暾的多, sm染色体少, st染色体相差不大, 其原因有可能是取材不同, 细胞分裂时期不同, 导致观察到染色体的效果也不相同, 也有可能是人工误差, 在肉眼判断着丝粒位置或手工测量臂长时出现一些误差。有关湘云金鳙细胞遗传学背景的研究表明[18], 湘云金鳙群体遗传多样性明显强于普通鳙鱼群体; 本文正是基于湘云金鳙与普通鳙鱼存在DNA分子水平上的差异, 才开展了对湘云金鳙的染色体组型分析。当然, 分子水平的差异不一定反映到染色体水平差异上来, 但本次得出洞庭湖区人工养殖湘云金鳙的2N = 48, 核型公式为: 24 m+18 sm+6 st, NF = 90, 与刘良国等[23]的研究相比, 总体特征基本相同, 说明鳙在遗传上具有保守性; 具体数据上又存在着些许差异, 表现出一定的多态性。这种差异如果不是测量误差所致, 可能正好反映出湘云金鳙与普通鳙鱼在分子水平上的差异。据此, 初步推测湘云金鳙染色体组型形成的原因, 可能是由于普通鳙鱼染色体发生易位、臂间倒位或异染色质增减等较大的染色体重排等变化造成。
鲢亚科的基本染色体二倍数为48, 余先觉等[12]在中国淡水鱼类染色体一书中认为鲢亚科鱼是通过罗伯逊易位由2N = 50的原始类群进化而来, 并指出在2N数相同的分类群体间, 染色体臂数变化从进化上低位类群到高位类群表现逐渐升高的趋势。本文报道的金鳙核型与昝瑞光[11]、刘凌云[24]、苏泽古[25]和余先觉[12]报道的鲢的核型相对比, 发现鲢的中部着丝粒染色体较少, 近端部着丝粒染色体较多, NF值相对较低, 故在鲢亚科中, 鲢应处于低位类群, 而鳙属于较高位类群。
鲤科隶属于鲤形目, 是世界现生鱼类中最大的一个科, 拥有约220属2 420种[27], 也是我国淡水鱼类中最大的一个科, 占全国淡水鱼类总数的一半以上。关于鲤科科以下类群的划分及其系统关系, 陈宜瑜[28]将鲤科鱼类划分为鲃系(分化为野鲮亚科、鲤亚科和鲃亚科)和雅罗鱼系(分化为雅罗鱼亚科、鲢亚科、鲴亚科、鲌亚科、鮈亚科、鳑鲏亚科、鱼丹亚科)。何舜平等[29]的研究表明鲢亚科与雅罗鱼亚科的亲缘关系最近, 而朱蓝菲等[30]利用同工酶表型对20种鲤科鱼类进行分析时发现鲢亚科与鲌亚科的关系较近, 凌去非等[31]用线粒体COII基因序列分析25种鲤科鱼类时也验证了朱蓝菲的观点。
表4中列出了雅罗鱼系中雅罗鱼亚科、鲌亚科和鲴亚科中几种鱼的染色体组型, 这几种鱼与鲢亚科鱼类染色体二倍体数相同、核型特征相似, 均取自洞庭湖水域, 且均由同一学者采用同一方法获得, 排除了由于实验方法或实验条件的不同而带来的可能的误差, 具有较高的参考价值。

表4 洞庭湖区雅罗鱼亚科、鲌亚科、鲴亚科和鲢亚科鱼类的核型比较
续表4
通过表4中4个亚科鱼类的染色体组型比较可知, 鲢亚科与鲌亚科的NF值和染色体组型最接近, 故本文支持朱蓝菲和凌去非的观点, 鲢亚科与鲌亚科的关系更近。鲴亚科中的银鲴与雅罗鱼亚科中的赤眼鳟端部和近端部着丝粒染色体较少, 中部与近中部着丝粒染色体较多, NF值相对较高。根据余先觉等[12]的研究, 在洞庭湖水域的这4个亚科中, 似乎鲴亚科与雅罗鱼亚科应处于较高位类群, 而鲢亚科与鲌亚科属于低位类群。当然, 表4中鲴亚科和雅罗鱼亚科每科均只列出了一种鱼的核型, 还需进一步对洞庭湖水域更多的鱼类进行染色体组型分析, 为确定雅罗鱼系各亚科的亲缘关系和系统演化提供依据。
[1] 湖北省水生生物研究所鱼类研究室. 长江鱼类[M]. 北京: 科学出版社, 1976.
[2] 农业农村部渔业渔政管理局. 2019年中国渔业统计年鉴[M]. 北京: 中国农业出版社, 2019.
[3] 伍献文, 钟麟. 鲩、青、鲢、鳙的人工繁殖在我国的进展和成就[J]. 科学通报, 1964, 9: 900–907.
[4] 陈启鎏. 青、鲩、鳙、鲢等家鱼寄生原生动物的研究—Ⅲ.寄生鳙和鲢的原生动物[J]. 水生生物学报, 1956(2): 279–298.
[5] 李思发, 杨和荃, 陆伟民. 鲢、鳙、草鱼摄食节律和日摄食率的初步研究[J]. 水产学报, 1980, 4(3): 275–283.
[6] 胡成钰. 鳙鱼(Aristichthysnobilis)外周血细胞的细微结构[J]. 江西科学, 1990, 8(3): 20–28.
[7] 吴力钊, 王祖熊. 长江下游鳙鱼天然种群的生化遗传变异[J]. 水生生物学报, 1991, 15(1): 94–96.
[8] 周桂. 鲢鳙鱼在膨化食品中的应用[J]. 广西民族学院学报(自然科学版), 1997, 3(2): 146–149.
[9] 周暾. 鳙鱼染色体组型的研究[J]. 淡水渔业, 1980(4): 5–9.
[10] 刘凌云. 鳙鱼的白血细胞培养及其染色体组型分析[J]. 北京师范大学学报(自然科学版), 1981(3): 79–83.
[11] 昝瑞光, 宋峥. 鲤、鲫、鲢、鳙染色体组型的分析比较[J]. 遗传学报, 1980, 7(1): 72–77.
[12] 余先觉, 周暾, 李渝成. 中国淡水鱼类染色体[M]. 北京: 科学出版社, 1989.
[13] 孔庆亮, 李宗芸, 傅美丽, 等. 鳙鱼染色体的DAPI核型分析[J]. 四川动物, 2006, (01): 64–67.
[14] 楼允东. 我国鱼类育种研究五年回顾[J]. 淡水渔业, 1999(9): 1–3.
[15] 沈俊宝. 长江“四大家鱼”资源急需保护和增殖[J]. 中国水产, 2002(12): 16–19.
[16] 杨福忠, 向升亮, 张兵, 等. 湘云金鳙的生物学特征及养殖特性[J]. 当代水产, 2019(4): 95–96.
[17] 孙远东, 袁志栋, 谭树华, 等. 人工诱导湘云金鳙雌核发育的研究[J]. 水产学报, 2015, 39(1): 8–15.
[18] 严斌, 彭亮跃, 谢伟民, 等. 新型湘云金鳙遗传多样性的RAPD及微卫星分析[J]. 激光生物学报, 2011, 20(1): 54–60.
[19] 刘飞, 孟昱林, 韩志琦, 等. 金鳙和黑鳙的肌肉营养成分分析及评价[J]. 淡水渔业, 2017, 47(2): 101–106.
[20] 杨福忠, 向升亮, 张兵, 等. 湘云金鳙的生物学特征及养殖特性[J]. 当代水产, 2018(12): 87–88.
[21] 刘良国, 赵俊, 崔淼. 尖鳍鲤的染色体组型分析[J]. 华南师范大学学报(自然科学版), 2004, 36(1): 108–111.
[22] Levan A, Tred K, Sandberg A. Nomencture for centromeric position on chromosomes [J]. Heredita, 1964, 52(2): 201–220.
[23] 刘良国, 杨春英, 杨品红. 洞庭湖水系鱼类资源与染色体研究[M]. 北京: 科学出版社, 2018.
[24] 刘凌云. 鲢鱼染色体组型的分析[J]. 遗传学报, 1981(3): 251–255, 287–288.
[25] 苏泽古, 许克圣, 陈尚萍, 等. 白鲢三倍体及其核型的研究[J]. 动物学研究, 1984(S2): 15–20, 83–84.
[26] 李思发, 周碧云, 倪重匡, 等. 长江、珠江、黑龙江鲢、鳙和草鱼原种种群形态差异[J]. 动物学报, 1989, 35(4): 390–398.
[27] Nelson J S. Fish of the world [M]. 4th edition. New York: Wiley, 2006.
[28] 陈宜瑜. 中国动物志, 硬骨鱼纲, 鲤形目(中)[M]. 北京: 科学出版社, 1998.
[29] 何舜平, 刘焕章, 陈宜瑜, 等. 基于细胞色素b基因序列的鲤科鱼类系统发育研究(鱼纲:鲤纲:鲤形目)[J]. 中国科学, 2004, 34(1): 96–104.
[30] 朱蓝菲, 陈湘粦, 王祖熊. 20种鲤科鱼类同工酶的表型分析及有关进化问题的探讨[J]. 水产学报, 1983, 7(2): 145–152.
[31] 凌去非, 李思发. 鲤科25种鱼类线粒体COⅡ基因序列差异及其系统进化关系[J]. 水产学报, 2006, 30(6): 747–752.
Karyotype analysis ofred var.
Yang Chunying1, Liu Liangguo2, Liu Fei3, Zhou Gongjian4, Li You3
(1. Normal College, Hunan University of Arts and Science, Changde 415000, China; 2. Teaching Affairs Department, Hunan University of Arts and Science, Changde 415000, China; 3. Key Laboratory of Zoology in Hunan Higher Education, Changde 415000, China; 4. Hunan Xiangyun Biology Science and Technology Limited Company, Changde 415000, China)
The chromosomes number and karyotype of artificial feedingred var. in Dongting lake region are analyzed by using phytohemagglutinin (PHA) and colchicine injection method in vivo to make renal cell chromosome specimens. The results show that the diploid number of metaphase chromosomes of the feedingred var. in Dongting lake region is 2N = 48, and the karyotype formula is 24 m+18 sm+6 st, NF = 90. The telocentric chromosome, the satellite chromosome or secondary constrictions are not found in the artificial feedingred var. in Dongting lake region. At the same time, the genetic diversity are explored by comparing with the karyotypes of the Hypophthalmichthyinae.
Dongting lake region;red var.; karyotype
10.3969/j.issn.1672–6146.2021.01.010
Q959.481
A
1672–6146(2021)01–0046–05
杨春英, 1124292768@qq.com; 刘良国, liuliangguo@ foxmail.com。
2020–03–18
湖南省教育厅基金项目(19C1270); 国家自然科学基金项目(31572619)。
(责任编校: 郭冬生)
