土壤细菌群落结构和多样性对不同林龄闽楠人工林的响应
2021-01-29
(中南林业科技大学 a.林学院;b.水土保持与荒漠化重点实验室,湖南 长沙 410004)
闽楠Phoebe bournei(Hemsl.) Yang 是中国传统珍贵用材树种和优良园林绿化树种,已被列入了国家重点发展的南方集林区珍贵用材树种和国家战略储备林树种之一[1]。土壤微生物群落结构和多样性直接影响土壤理化性质及土壤养分的组成与转化[2]。土壤细菌对土壤环境质量的变化高度敏感,常被用来评价农林复合生态系统的环境变化[3],土壤细菌群落结构和多样性的变化特征在生态系统的养分循环中起着关键作用,影响着植被的发育和演替[4]。土壤微生物作为热点问题,已成为近年来国内外土壤学研究的热点之一。人工林年龄的变化对土壤微生物多样性[5]和微生物群落结构[6]有很强的影响。王超群等[7]对不同林龄杉木人工林土壤微生物群落代谢功能差异进行研究,结果表明0~20 cm 土层土壤微生物群落的代谢活性、Shannon-Wiener 多样性指数、Pielou 均匀度指数、Simpson 优势度指数、McIntosh 多样性指数和McIntosh 均匀度指数均随林龄的增加而增加。牛小云等[8]通过对辽宁不同林龄日本落叶松人工林土壤微生物、养分及酶活性进行研究,结果表明土壤微生物、养分及酶活性各项指标基本呈现11 或47 a 林龄较高,20 或34 a 林龄较低,且随着林龄增加土壤微生物群落结构及酶活性变化呈减小的趋势。此外,土壤中细菌群落结构受土壤理化性质的驱动,有机质与土壤微生物群落结构之间存在很强的关联性[9],土壤中的氮磷钾元素是影响细菌群落组成的主要因素[10]。目前,随着人工林年龄增长,土壤微生物方面的研究在桉树[11]、松树[12]和杉木[13]等速生树种中研究较多,而关于闽楠人工林的研究,大多集中在对闽楠人工林的生物量[14]和碳密度[15],优良家系的培育、育苗技术[16-17]和遗传改良[18]及其生长特性等方面[19-20],对于不同林龄闽楠人工林微生物群落结构和多样性,及其与土壤化学性质的研究却很少被关注。本研究选择5年生、10年生、15年生和40年生闽楠人工林作为研究对象,通过Illumina HiSeq 高通量技术和冗余分析的方法探究不同林龄闽楠人工林土壤细菌群落结构和多样性的演变特征及其与化学性质的关系,旨在揭示细菌群落结构和多样性对不同林龄闽楠人工林的响应机制,找出影响土壤细菌群落结构和多样性的最敏感的环境因子,以期为促进闽楠人工林土壤质量的改善,以及闽楠人工林高效可持续经营提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于湖南永州市金洞林场(110°53′25″—112°13′17″E,26°2′33″—26°21′25″N)。属于中亚热带东南季风湿润气候区,年均降水量为1 600~1 900 mm,年平均气温16.3~17.7℃,年均日照时间1 300~1 740 h,无霜期293 d。土壤类型为第四纪红壤和黄壤,成土母岩以砂岩、页岩、碳质板岩为主。研究区内森林覆盖率达86%,林木蓄积达2.98×106m3,林场境内森林茂盛,物种多样,据调查有高等植物210 科,1 557 种,属国家保护植物银杏、红豆杉、钟萼木、伞花木、吉赤皮青冈、古圆槠、古楠木等56 种。
1.2 样地设置与样品的采集
分别选择坡度、坡向、海拔、土壤和地形等立地因子相同或相似的不同林龄闽楠人工林试验样地,分别为:5年生闽楠幼龄林(Y5)、10年生闽楠中龄林(Y10)、15年生闽楠近熟龄林(Y15)和40年生闽楠成熟林(Y40)。在每个林龄分上坡、中坡和下坡设置3 块20 m×20 m 的标准地,共12块标准地,并对样地内林木进每木检尺,测定胸径、树高,调查样地的地形地貌、土壤类型、群落结构及林下植被进行调查。样地基本概况见表1。
土壤样品采集于2019年3月中旬,在每块标准样地内采用“S”布点采取土壤样品,取18 个土芯(直径5 cm),取去除土壤表层的腐殖质及杂质后的0~20 cm 的土样混合成一个复合样品。然后利用四分法将土壤分为4 份取其中一份,每一个林龄总共有3 个复合样品。从每一个取出的复合样品中,取大约10 g 去除根和石子的新鲜土过2 mm 筛,放入灭菌后的离心管中并立即放入液氮中用于16 Sr DNA 分析,到达实验室后样品保存在零下80 ℃的冰箱中,其余的新鲜土(约500 g)自然风干后过2 mm 的筛用于化学性质的测定。
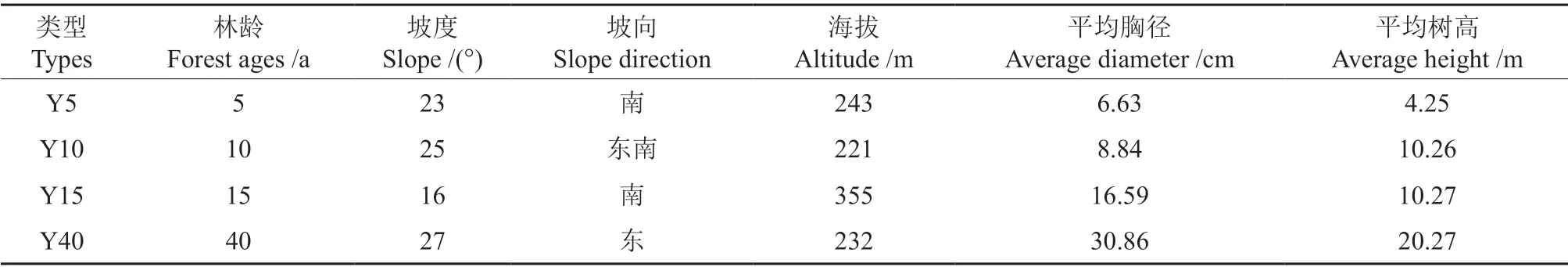
表1 不同林龄阶段闽楠人工林样地基本概况Table 1 Basic overview of the plantation of Phoebe bournei in different stages of forest ages
1.3 土壤化学性质的测定
土壤pH 值采用水土比2.5:1 的酸度计法;有机碳(SOC)采用高铬酸钾高温外加热法;全氮(TN)采用凯氏定氮法;全磷(TP)和全钾(TK)采用氢氧化钠熔融法;碱解氮(AN)采用碱解扩散法;土壤有效P(AP)、速效K(AK)采用Mehlich3浸提液-原子吸收分光光度计法。
1.4 DNA 提取及细菌16S rDNA PCR 扩增及测序技术
土壤总DNA 采用CTAB 方法对样本的基因组DNA 进行提取之后利用琼脂糖凝胶电泳检测DNA 的纯度和浓度,取适量的样品于离心管中,使用无菌水稀释样品至1 ng/μL。以稀释后的基因组DNA 为模板,16srdnav3-v4 区引物为341F (CCTAYGGGRBGCASCAG)-806R(GGACTACNNGGGTATCTAAT),进行PCR 扩增。酶和缓冲液使用New England Biolabs 公司的Phusion®High-Fidelity PCR Master Mix with GC Buffer。使用高效和高保真的酶进行PCR,确保扩增效率和准确性。PCR 反应体系和程序(30 μL):Phusion Master Mix(2×)15 μL;Primer(2 μmol)3 μl(6 μmol);gDNA(1 ng/μL)10 μL(5~10 ng)H2O2μL 反应程序:98℃预变性1 min;30 个循环(98℃,10 s;50℃,30 s;72℃,30 s);72℃,5 min。采用Bio-radT100 梯度PCR 仪进行分析,PCR 产物使用2%浓度的琼脂糖凝胶进行电泳检测。根据PCR 产物浓度进行等浓度混样,充分混匀后使用1×TAE 浓度2%的琼脂糖胶电泳纯化PCR 产物,选择主带大小在400~450 bp 之间的序列,割胶回收目标条带。产物纯化试剂盒使用的是Thermo Scientific公司GeneJET胶回收试剂盒。
1.5 细菌多样性分析
使用Qiime 软件(Version 1.7.0)计算Shannon均匀度指数、Simpson 均匀度指数、Ace 丰富度指数和Chao1 丰富度指数。
1.6 数据统计方法
统计方法采用Microsoft Excel 2016、SPSS20.0和Canoco 5.0 数据处理软件,并进行ANOVA 单因素方差分析,采用LSD 差异显著性检验比较不同林龄闽楠人工林土壤化学性质和微生物多样性的差异,利用冗余分析(RDA)揭示土壤化学性质与微生物群落结构和多样性的相互关联机制。
2 结果与分析
2.1 不同林龄闽楠人工林土壤化学性质的差异
土壤pH 值、有机碳、全氮、全磷、全钾、碱解氮和速效钾在不同林龄闽楠人工林之间均存在显著差异(P<0.05),而有效磷在不同林龄闽楠人工林之间差异性不显著(P=0.185)(表2)。其中,土壤pH 值、有机碳、速效钾含量随着林龄增加先增高后下降,在15年生时最高,分别为4.09、23.55 g/kg 和150.50 mg/kg。全氮的含量随着林龄增加先增高后下降再增高的趋势,在10年生时最高为3.16 g/kg。全磷的含量总体上随着林龄增加而增高,在40年生时最高为0.98 g/kg。全钾的含量随着林龄增加先下降后增高,在5年生时最高为7.45 g/kg。碱解氮的含量随着林龄增加呈现先下降后增高再下降的趋势,在15年生时最高为158.23 mg/kg。闽楠人工林土壤类型主要属于酸性红壤(pH 值<4.5)。
2.2 不同林龄闽楠人工林土壤细菌测序数据分析
16S rDNA 检测表明,在97%的相似水平下,随着林龄增加OTUs 平均个数先增高后下降,在10年生时最高为3 179 个,5年生时最小为2 755个,12 个样品共34 998 个OTUs 被测出,每个样品范围为1 939~2 420,平均OTU 个数2 142 个。样品共得到616 646 条序列数,各个样品的有效序列长度一般在411 bp 左右,文库覆盖率范围为98.94%~99.52%,说明样品基因序列被检出的概率很高,能够反映检测样品土壤细菌群落的真实情况(表3)。12 个样品的曲线趋向平坦时,说明测序数据量渐进合理,更多的数据量只会产生少量新的OTUs(图1)。
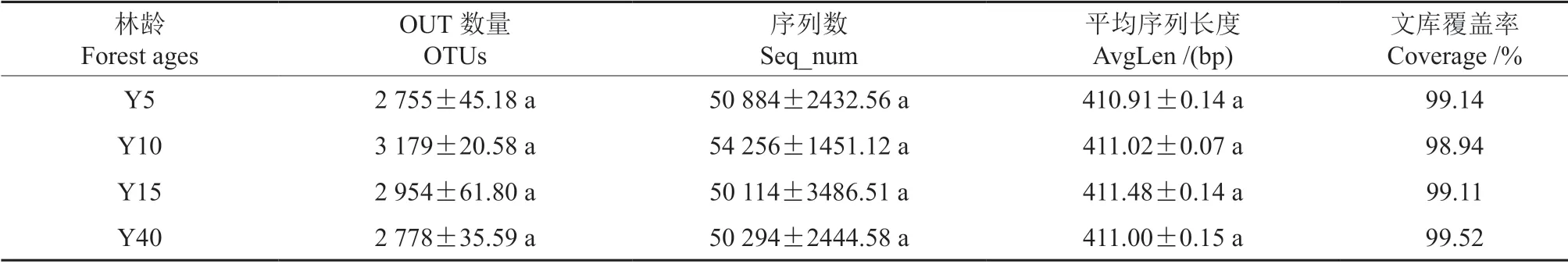
表3 不同林龄闽楠人工林土壤细菌测序数据Table 3 Sequencing data of soil bacteria in different ages of Phoebe bournei
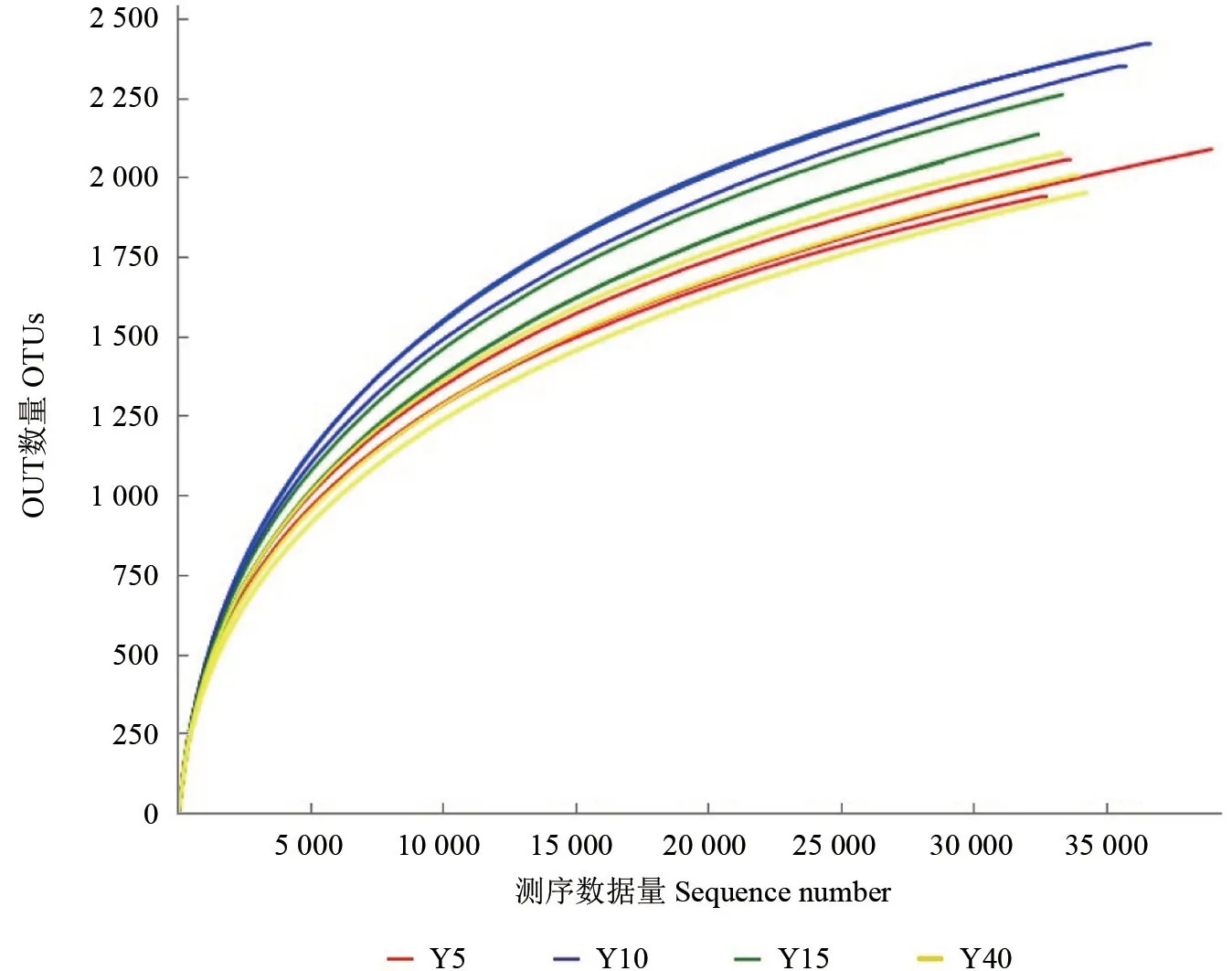
图1 OUT 稀释曲线Fig.1 OUT dilution curve
2.3 不同林龄闽楠人工林土壤细菌群落组成和结构分析
不同林龄闽楠人工林细菌群落组成存在显著差异,尤其是5年生和10年生闽楠人工林差异最为显著(图2)。主坐标分析(PCoA)结果显示,PC1 的贡献率为41.21%,PC2 的贡献率为23.75%,两者共解释了64.96%。从样点的细菌群落组成来看,在PC1 中,5年生、15年生和10年生闽楠人工林细菌群落组成差异较大,尤其是5年生和10年生闽楠人工林差异最为显著,而40年生与10年生和15年生闽楠人工林细菌群落组成差异不显著;在PC2 中,15年生和10年生闽楠人工林细菌群落组成差异较大,而40年生和5年生闽楠人工林细菌群落组成差异不显著。
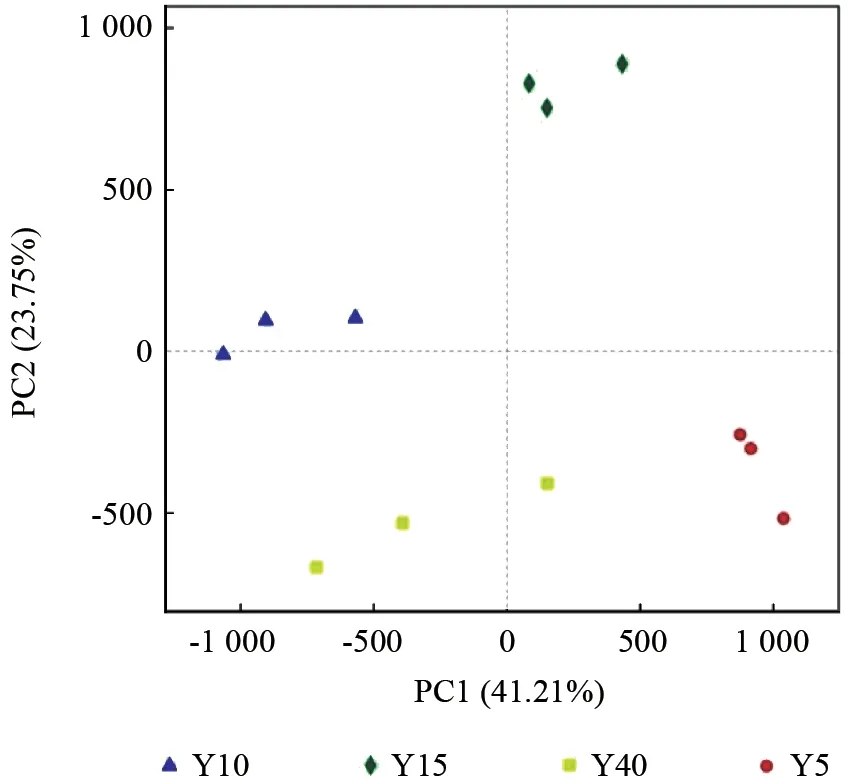
图2 土壤细菌OTU 主坐标分析Fig.2 Analysis of the main coordinates of soil bacteria OTU
对OTU 代表序列进行物种注释并归类,12个样品共识别出21 门,43 纲,88 目,153 科,204 属和395 种。12 个样品的土壤细菌门分类群落饼图显示:排名前10 主要菌门中,变形菌门(Proteobacteria)(30.87%)、酸杆菌门(Acidobacteria)(26.98%)、绿弯菌门(Chloroflexi)(16.49%)、放线菌门(Actinobacteria)(9.80%)为主要菌门(图3)。同时,根据不同林龄闽楠人工林土壤细菌群落优势菌门相对丰度的热图显示:细菌群落菌门的相对丰度在不同林龄条件下存在差异,且各菌门的差异性均不相同(图4)。
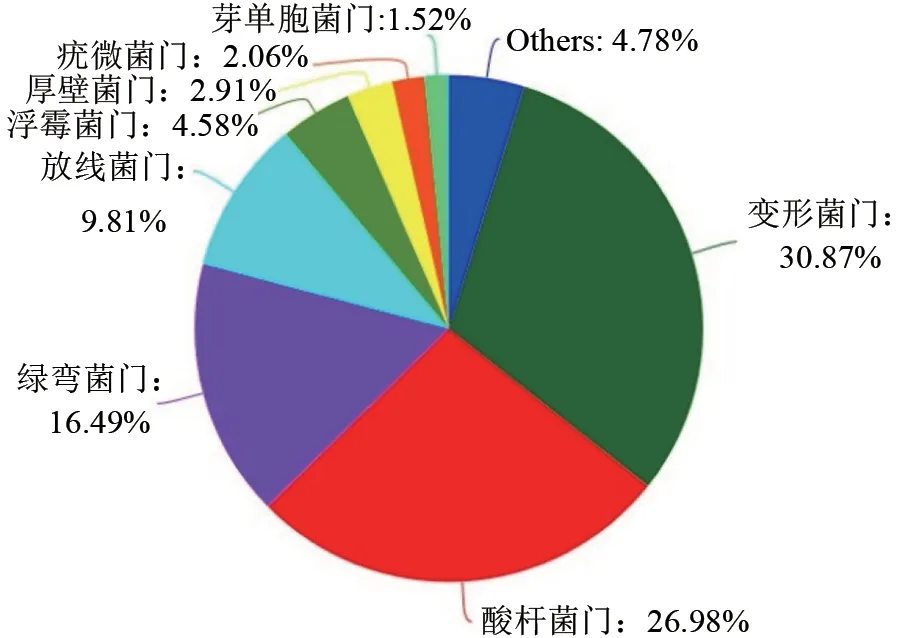
图3 林龄闽楠人工林土壤细菌门分类群落饼Fig.3 Soil bacterium gate classification community pie chart of Pinnacle sinensis plantation
不同林龄闽楠人工林土壤细菌群落的相对丰度在门水平下绿弯菌门、厚壁菌门和GAL15 均有显著性差异(P<0.05),而在变形菌门、酸杆菌门、放线菌门、浮霉菌门、疣微菌门、芽单胞菌门、拟杆菌门和衣原体中无显著性差异(P>0.05,表4)。其中,绿弯菌门和GAL15 的相对丰度随着林龄增加呈现先增高后下降再增高的趋势,在10年生时最高,分别为23.47%和1.49%,显著高于5年和15年生闽楠人工林,厚壁菌门的相对丰度随着林龄增加总体呈下降的趋势,在5年生时最大为3.69%,显著高于15年和40年生闽楠人工林。此外,变形菌门在15年生闽楠人工林中含量最丰富,酸杆菌门和绿弯菌门在10年生闽楠人工林中含量最丰富,而浮霉菌门和厚壁菌门在5年生闽楠人工林中含量最丰富,放线菌门在40年生闽楠人工林中含量最丰富(图5)。
2.4 不同林龄闽楠人工林土壤细菌群落多样性分析
不同林龄闽楠人工林土壤细菌群落多样性Shannon 均匀度指数、Simpson 均匀度指数、Ace丰富度指数和Chao1 丰富度指数均存在显著差异(P<0.05,表5),其中Simpson 均匀度指数和Ace 丰富度指数随着林龄增加呈现先下降后增高再下降的趋势,在15年生时最高,分别为0.24和32.31。Shannon 均匀度指数随着林龄增加呈现先增高后下降再增高的趋势,在10年生时最高为1.92。Chao1 丰富度指数随着林龄增加先增高后下降,在15年生时最高为26.33。
2.5 土壤细菌群落结构和多样性与化学性质的冗余分析
蒙特卡洛检验表明土壤化学性质与优势细菌菌门的相对丰度显著相关(P<0.05),其中,有机碳、有效钾和全磷与土壤细菌群落结构相关性较强。第一主轴解释了62.89%的变化,第二主轴解释了23.93%的变化,两者共解释了86.82%的变化。有机碳、有效钾和全磷与RDA1 正相关性最强,主要表现在:其与芽单胞菌门、变形菌门和放线菌门较强的正相关,与厚壁菌门、酸杆菌门、疣微菌门、绿弯菌门和芽单胞菌门较强的负相关性。碱解氮和全钾与RDA2正相关性最强,主要表现在:其与浮霉菌门、变形菌门和放线菌门较强的正相关,与GAL15 和绿弯菌门较强的负相关性。此外,全氮与RDA2 负相关性最强,其与土壤主要优势菌门的相关性与碱解氮和全钾的相反(图6)。

图4 不同林龄闽楠人工林土壤细菌群落优势菌门相对丰度的热图Fig.4 Heat map of the relative abundance of dominant bacterial species in soil bacterial communities of different ages of Phoebe bournei
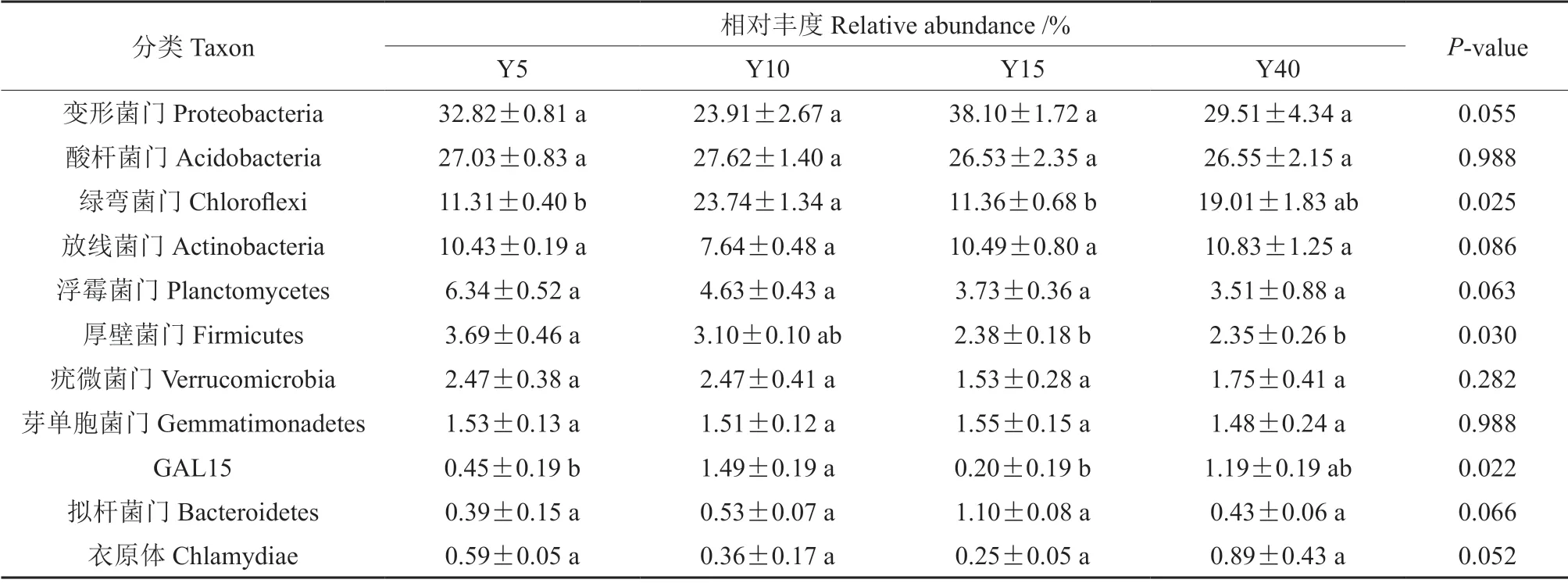
表4 不同林龄闽楠人工林土壤优势细菌菌门的相对丰度Table 4 Relative abundance of dominant bacteria in the soil of different ages of Phoebe bournei
蒙特卡洛检验表明土壤化学性质与优势细菌菌门的相对丰度显著相关(P<0.05),其中,有机碳、有效钾和全磷与土壤细菌群落多样性相关性较强。第一主轴解释了66.15%的变化,第二主轴解释了21.62%的变化,两者共解释了86.77%的变化。有机碳、有效钾和全磷与RDA1 正相关性最强,主要表现在:其与Simpson 均匀度指数和Chao1 丰富度指数较强的正相关,与Ace 丰富度指数和Shannon 均匀度指数较强的负相关性。碱解氮和全钾与RDA2 正相关性最强,主要表现在:其与Ace 丰富度指数、Simpson 均匀度指数和Chao1 丰富度指数的较强正相关,与Shannon 均匀度指数较强的负相关性。此外,全氮与RDA2 负相关性较强,其与土壤细菌多样性指数的相关性与碱解氮和全钾的相反(图7)。
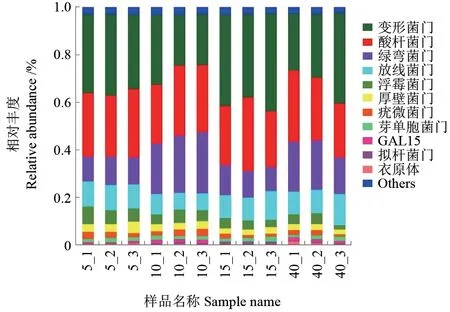
图5 不同林龄闽楠人工林土壤细菌群落优势菌门相对丰度分布Fig.5 Relative abundance distribution of dominant bacteria in soil bacterial communities of different ages of Phoebe bournei

表5 不同林龄闽楠人工林土壤细菌群落多样性差异Table 5 Differences in soil bacterial community diversity in different plantations of Pinus sylvestris
3 讨 论
3.1 林龄对闽楠人工林土壤化学性质的影响
本研究发现,不同林龄闽楠人工林土壤pH值、有机碳、全氮、全磷、全钾、碱解氮和速效钾均存在显著差异,其中15年生闽楠人工林有机碳、碱解氮和速效钾的含量显著高于其它林龄,而5年生闽楠人工林的有机碳、全氮、全磷和速效钾的含量显著低于其它林龄,这表明闽楠人工林在不同的发育周期,其土壤肥力质量变化比较大,在经营的过程中更应该注重施肥等管理措施。然而随着闽楠人工林的持续种植,40年生闽楠人工林土壤有机质、碱解氮和速效钾的含量显著小于15年生,这可能是由于持续经营方式不断消耗土壤地力,而在目前的经营方式下并未及时补充营养元素,仅靠凋落物分解难以维持林地的持续经营。胡士达[21]对不同林龄闽楠人工林生态化学计量特征的研究表明土壤中有机碳碳、全氮含量均表现为14 a>60 a>45 a,14 a 和60 a 显著高于45 a;全磷含量表现为45 a>60 a>14 a,这与本研究的研究结果相一致。此外,5 a 到10 a,土壤有机碳、全氮和速效钾均呈显著增加的趋势,而全钾和碱解氮呈显著减小的趋势;10 a 到15 a,土壤有机碳、全磷、全钾、碱解氮和速效钾均呈显著增加的趋势,而全氮呈显著减小的趋势;15~40 a,土壤全磷呈显著增加的趋势,而有机碳、碱解氮和速效钾全氮均呈显著减小的趋势,这可能是由于闽楠不同的生长发育周期,其对土壤元素的吸收利用效率不同导致的,而且这种利用机制的不同,导致土壤养分的不均匀效应。大量研究表明,不同树种在不同林龄类型下,土壤养分元素的增加和减小的趋势并不一致[22-23]。此外,在不同林龄条件下,土壤有效磷的含量无显著性差异,可能是由于在酸性土壤中极易形成磷酸盐沉淀,这也通常是亚热带酸性土壤中磷的有限性的限制性因子。
3.2 林龄对闽楠人工林土壤细菌群落结构和多样性的影响
大量研究表明,随着森林种植林龄的增加土壤微生物群落结构会发生巨大影响[24]。在本研究中,不同林龄闽楠人工林细菌群落组成存在显著差异,尤其是5年生和10年生闽楠人工林差异最为显著。这可能是由于随着林分的增加植物—土壤之间的交互作用发生改变,导致植物生理生化机制进行不同类型的分化,及其与土壤的适应和相互协调能力的改变,从而导致土壤微生物群落结构的差异[25]。同时,本研究检测出变形菌门(30.87%)、酸杆菌门(26.98%)、绿弯菌门(16.49%)和放线菌门(9.80%)为闽楠人工林的主要优势菌门,其中绿弯菌门、厚壁菌门和GAL15 在不同林龄条件下存在显著差异(P<0.05)。随着林龄增加土壤微生物群落结构变化的差异与徐志霞等[26]的结果相似。此外,我们还发现,不同林龄闽楠人工林土壤细菌群落多样性Shannon 均匀度指数、Simpson 均匀度指数、Ace 丰富度指数和Chao1 丰富度指数均存在显著差异(P<0.05),其中,15年生闽楠人工林细菌多样性指数Simpson 均匀度指数、Ace 丰富度指数和Chao1 丰富度指数均显著高于其它林龄类型。这一结果说明随着林龄的增加,15年生闽楠人工林土壤细菌多样性指数达到了较高的水平,而在40年生闽楠人工林中土壤细菌多样性指数有所下降。
本研究中,测量的土壤化学因子分别解释了土壤微生物群落结构和多样性总变异系数的86.82%和86.77%,这表明土壤微生物的群落结构和多样性更容易受到土壤化学性质的影响,其通过微生物的呼吸,矿化和反硝化等代谢活动来调节地球的化学环境。我们的研究结果中,随着林龄的增加,土壤pH 值、有机碳、全氮、全磷、全钾、碱解氮和速效钾含量均呈现不同程度的增加或减少,土壤化学性质的变化可能是导致土壤微生物群落结构和多样性发生明显变化主要原因。此外,本研究还发现有机碳、有效钾和全磷与土壤细菌群落结构和多样性显著相关,表明土壤有机碳、有效钾和全磷可能是影响闽楠人工林土壤中微生物活动的重要因素。大量研究表明,微生物生长和活性通常受土壤有机碳,氮和磷的有效性的限制[27],而本研究中有机碳、有效钾和全磷是闽楠人工林土壤微生物生长的关键因子,与微生物群落结构和多样性密切相关。因此,在确定闽楠人工林土壤微生物群落结构和多样性中土壤环境因子的影响值得进一步研究,以了解如何更好地利用微生物,改善土壤环境,提高其在森林土壤环境的利用效率。此外,本研究仅对不同林龄闽楠人工林土壤真菌进行探讨分析,而未涉及真菌和土壤微生物功能多样性的探讨,在接下来的研究中重点是应该将土壤微生物细菌、真菌结构和多样性进行结合探讨,并与土壤微生物功能多样性进行衔接,以期更加深入了解闽楠人工林土壤微生物的演变对林龄的响应机制,同时本研究拟为闽楠人工林在以后新型肥料的研发提供一定的参考意见。
4 结 论
研究结果表明不同林龄闽楠人工林土壤化学性质、微生物群落结构和多样性均存在显著差异。随着林龄的增加,15年生闽楠人工林有机碳、碱解氮和速效钾的含量达到较高的水平,而随着林龄增加到40 a,土壤有机碳、碱解氮和速效钾全氮均呈显著减小的趋势。微生物群落结构和多样性在不同林龄闽楠人工林样地中具有显著差异,本研究检测出变形菌门(30.87%)、酸杆菌门(26.98%)、绿弯菌门(16.49%)和放线菌门(9.80%)作为闽楠人工林的主要优势菌门,其中绿弯菌门和GAL15 均在10年生闽楠人工林中的相对丰度显著高于5年和15年生,厚壁菌门在5年生闽楠人工林中的相对丰度显著高于15年和40年生。闽楠人工林细菌多样性指数Simpson 均匀度指数、Ace 丰富度指数和Chao1 丰富度指数均表现为15年生显著高于其它林龄类型。此外,从研究的土壤环境因子的范围来看,有机碳、有效钾和全磷是影响土壤微生物群落结构和多样性的关键决定性因素。因此,在进行闽楠人工林培育和种植的过程中,应该关注土壤微生物的演变特征,及其与土壤环境因子的相互作用机制,以期为闽楠人工林的高效经营以及以后新型肥料的研发提供一定的参考意见。
