海岸带生态恢复区不同林龄红树林对CH4和CO2排放通量的影响
2021-01-28李森蔡厚才陈万东林利倪孝品伍尔魏曾贵候唐剑武李香兰
李森,蔡厚才,陈万东,林利,倪孝品,伍尔魏,曾贵候,唐剑武,李香兰*
1. 北京师范大学全球变化与地球系统科学研究院,北京 100875;2. 北京师范大学遥感科学国家重点实验室,北京 100875;3. 南麂列岛国家海洋自然保护区管理局,浙江 温州 330326;4. 华东师范大学河口海岸学国家重点实验室,上海 200062
甲烷(CH4)是仅次于二氧化碳(CO2)的最重要的温室气体。根据IPCC第五次评估报告,1750—2011年大气中CO2和CH4体积分数分别从278×10-6和 772×10-9增加到 390×10-6和 1803×10-9,增加了40%和150%(IPCC,2013)。虽然空气中CH4体积分数远远小于CO2,但是其全球变暖潜能在100年时间尺度内是CO2的28倍(IPCC,2013)。通过陆地生态系统和海洋生态系统吸收大气中 CO2和CH4,形成了当前的陆地碳汇(1.5 Pg·a-1)和海洋碳汇(2.3 Pg·a-1),剩余 4.0 Pg·a-1停留在大气中(IPCC,2013)。通过提高海洋和陆地碳汇功能,可以有效减少大气中温室气体浓度,从而减缓全球气候变暖。受潮汐影响的海岸带生态系统较为脆弱,对于海陆交界的碳汇功能研究才刚刚起步。
海岸带蓝碳被定义为海岸带盐沼湿地、红树林和海草床等海岸带植物固定的碳(Mcleod et al.,2011;章海波等,2015;Howard et al.,2016;唐剑武等,2018),这三类生态系统是生物多样性最丰富,生产力最高、生态服务功能最大的湿地生态系统,其单位面积固碳能力是陆地生态系统的 10倍以上(章海波等,2015)。红树林是热带、亚热带海岸潮滩上由红树科植物为主组成的一种特殊植被类型,一般分布在 32°S—32°N 的滨海湿地。全球约有红树林152000 km2,占陆地森林面积的0.4%,分布在 118个国家,占全球热带森林湿地 30%—35%(Bhattarai et al.,2011)。中国现有红树林总面积 220 km2,主要分布于东部及东南沿海滩涂(但新球等,2016);红树林拥有强大的根系,是海岸防护林体系的第一道防线,是海边鸟类、鱼虾的重要栖息地,从而维持海岸带生态平衡(Nellemann et al.,2008)。以往研究大都关注红树林湿地碳储量,探索红树林湿地碳循环过程及其调控机制,从而对红树林湿地进行合理的保护和利用。全球红树林湿地碳汇能力在0.18 Pg·a-1,其中东南亚地区深达3 m的热带红树林湿地的碳储量平均高达 102.3 kg·m-2(Donate et al.,2012),而中国红树林的平均碳汇能力在 209—661 g·m-2·a-1(张莉等,2013)。受海岸带发展、水产养殖的扩张和过度砍伐等因素影响,海岸带红树林面积和分布范围在过去的半个世纪减少了30%—50%(Bouillon et al.,2008)。受海平面上升影响,2080年全球20%的海岸带蓝碳生态系统将面临消失,将导致生态功能损失、碳释放潜力增大、台风等极端气候、搬迁和移民等问题产生的经济损失与社会压力(Jachowski et al.,2013)。人类活动破坏红树林蓝碳生态系统,但通过保护和恢复能够增加蓝碳碳汇功能,以作为减缓气候变化的重要手段。
林龄是影响红树林蓝碳功能的重要影响因素。中国在九龙江口的研究中发现,24年生的秋茄(Kandelia candel)与48年生的秋茄年净固碳量分别为 1851、701 g·m-2·a-1,高于中亚热带杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)和楠木(Phoebe zhennan)等林分及针叶阔叶林的平均水平(258 g·m-2·a-1)(金亮等,2013)。陈海生等(2017)对中国分布最北的红树林种植区西门岛海洋特别保护区的研究发现,17年生的秋茄幼林树红树林湿地土壤呼吸作用始终高于60年生的成树林红树林湿地,例如,秋茄幼林树红树林湿地在夏季 7月土壤呼吸日平均速率为 5.8 μmol·m-2·s-1,而成林树红树林湿地土壤呼吸日平均速率为 5.35 μmol·m-2·s-1,其他季节同样表现为幼林高于成树林(陈海生等,2017)。通过比对不同林龄红树林温室气体排放通量进行分析,可进一步提高蓝碳收支的精度提供重要参考,为发展“蓝色港湾”的国家战略提供重要支撑。
浙江属于中国红树林分布的最北端,受气温和土壤环境影响,其海岸带生态恢复区红树林温室气体排放通量与变化趋势尚不明了。鳌江是浙江省八大水系之一,与钱塘江、闽江并称全国三大涌潮江。随着气候变化与人类活动加剧,鳌江入海口的喇叭口面积正逐年减少。全球气候变暖引起的海洋风暴、台风、海啸等极端气候现象增多,其直接后果就是侵蚀沙滩和滩涂;人类围海造田、围海养殖、过度捕捞和污染、人工砍伐红树林等严重侵害海岸带蓝碳生态系统。本文以鳌江口不同期移植红树林生态恢复区为研究对象,通过比较不同林龄红树林群落和土壤源温室气体排放通量,为完善蓝色碳汇的定量估算方法提供重要数据基础。
1 材料与方法
1.1 研究区域
本研究观测平台位于浙江南部鳌江镇“浙南蓝碳生态过程监测试验站”(27°34′58″E,120°34′13″S),分别于2014年和2018年在海岸带滩涂种植了秋茄红树林,林龄分别为移植5年和移植1年。该试验站位于浙江温州湾鳌江流域,属于中亚热带海洋性季风气候区,鳌江流域面积为1580.40 km2,年均降水量为1500—1800 mm,降水多集中在5—8月,年平均气温17.6 ℃,夏季多台风暴雨。鳌江滨海湿地位于温州湾东南部,呈东西条带状分布于鳌江两岸,潮汐属非正规半日潮,在一个月内分别在阴历月初与月中会出现两次最大潮差,最大潮差可达 603 cm。基于“浙南蓝碳生态过程监测试验站”现有观测平台,结合试验站周边新种植的红树林幼苗区,通过地面原位观测数据定量分析不同林龄红树林的温室气体排放通量和植被生长量,旨在探究不同林龄红树林碳汇功能及影响机制。
1.2 试验设计
1.2.1 CO2与CH4温室气体排放通量
通过理加便携式温室气体分析仪(LGR SF-3000,U.S.A.,Lica United)监测移植1年和移植5年红树林群落与土壤源 CO2与 CH4温室气体排放通量。理加便携式温室气体分析仪包括明箱(又称“同化箱”)与暗箱(又称“呼吸室”),明箱观测的是生态群落CO2和CH4通量,每半小时收集一个数据,持续自动观测。由江边至江岸间隔30 m为一个梯度,每个梯度随机选择一个观测点位,共3个观测点,其中高潮滩和低潮滩为5年移植林龄红树林群落,中潮滩为1年移植林龄红树林群落。每个观测点位分别固定安放一套透明同化箱(80 cm×80 cm×100 cm),箱体内是被观测的红树林植被。
理加便携式温室气体分析仪3个气室依次共同连接在同一台LGR主机,测定获取频率1 Hz,设置每个气室测定时间 5 min,每个气室间隔半个小时可自动进行一次气体数据的收集采样,每台气室每天可获取48个通量数据。由于电力供应有限,新生红树林群落与老红树林群落无法同步测定,但充分保证老与新生红树林气体数据保持连续不间断的收集,且时间相邻、同期进行。由于受潮水和仪器故障影响,本文研究周期分别包括2019年9月1—20日和10月11—17日沿水位梯度对移植5年红树林进行观测,9月20—24日和10月18—20日对移植1年红树林进行观测。
1.2.2 红树林生物量观测
沿着水位梯度(低潮滩、中潮滩和高潮滩)对移植5年的红树林种植区域随机设置3个边长为10 m的样方,每个样方内随机选取 50株红树林进行挂牌标记,共计150株。对新移植的1年的红树林随机选取2个样方,共计100株。每个小区5—10 m边长。每一株用带号码的铝标签标注。
1.2.3 土壤样品采集与测定
不干扰植被生长的情况下,在红树林恢复区,由低潮滩至高潮滩随机选取3条样带。每条样带每隔20 m布设1 m×1 m的样方。利用土钻随机采集3个深度为0—100 cm的土柱,每次采样分为5个土层剖面(每层厚度为20 cm)作为土壤样品,放入自封袋密封保存,运回实验室,用于相应样地土壤理化性质的测定。土壤有机碳的测定采用重铬酸钾容量法-外加热法,利用LWY84B型控温式铝体消煮炉。采用残渣烘干——质量法进行土壤水溶性盐总量的测定;硫酸根离子测定采用EDTA间接滴定法;土壤酸碱度(pH)采用电位测定法与电导法测定——电导率仪为DDS-307进行测定。
1.3 数据分析
运用 Excel 2016软件对原始数据进行初步统计,将原始通量数据进行连续变化的图像处理。利用SPSS 17.0统计分析软件,以小时为单位,将不同林龄红树林的同一月份内每天(24 h)同一小时内的通量进行统计并计算出均值,得出结果为不同林龄的红树林在不同月份的日变化,同时进行标准误差的计算。根据计算结果,运用SPSS17.0的单因素方差(One-way ANOVA)分析不同树龄的红树林温室气体通量之间的差异性。运用Excel 2016软件对土壤理化因子的数据进行结果整理与绘图。
2 结果
2.1 不同林龄红树林群落CH4和CO2通量动态变化
采用理加透明同化箱观测了2019年9—10月连续两月的红树林生态恢复区移植1年中潮位以及移植5年高潮位和低潮位红树林 CH4和CO2动态变化趋势(图1)。结果表明,浙江南部海岸带不同林龄红树林恢复区 CH4通量变化区间为-5—70 nmol·m-2·s-1,总体为大气 CH4的排放源。移植 5 年红树林 CH4通量变化为 3—70 nmol·m-2·s-1,移植 1年红树林 CH4通量变化为-5—35 nmol·m-2·s-1。
整个红树林恢复区 CO2通量变化为-25—15 μmol·m-2·s-1,总体而言,移植 5 年红树林为大气 CO2的吸收汇,而移植 1年红树林为大气 CO2的排放源。移植5年红树林CO2通量变化区间为-25— -14 μmol·m-2·s-1,高潮滩和低潮滩红树林平均值分别为-2.59 μmol·m-2·s-1和-0.87 μmol·m-2·s-1。移植 1 年红树林中潮位 CO2排放通量为-3—15 μmol·m-2·s-1,平均值为 2.85 μmol·m-2·s-1。移植 1 年红树林群落,9月呼吸作用大于光合作用CO2通量表现为碳源,平均排放通量为 5.06 μmol·m-2·s-1,而 10 月随着红树林植被生长,其光合作用增强,CO2吸收量增大,总体释放量明显降低,平均值为 0.63 μmol·m-2·s-1。
2.2 不同林龄红树林群落CH4和CO2排放通量比较
比较观测期内红树林恢复区CH4和CO2平均气体通量,结果发现,鳌江口海岸带红树林生态恢复区不同林龄的红树林都为大气 CH4的排放源(0.67±0.06) mg·m-2·h-1和 CO2的吸收汇 (-13.81±18.68) mg·m-2·h-1(图 2)。对 CH4排放通量而言,移植 5年红树林的高潮滩 CH4排放通量 (1.06±0.11)mg·m-2·h-1显著高于移植 5年红树林的低潮滩(0.45±0.03) mg·m-2·h-1和移植 1 年红树林的中潮位CH4通量 (0.38±0.07) mg·m-2·h-1(P<0.01),其中移植5年低潮滩和移植一年的中潮滩没有显著性差异(P>0.05)。对CO2通量而言,移植5年的红树林吸收大气中的 CO2,高潮滩和低潮滩红树林 CO2通量分别 为 (-82.63±22.9) mg·m-2·h-1和 (-26.05±17.94)mg·m-2·h-1。移植 1 年的红树林幼苗表现为 CO2的源,其碳通量为 (196.95±46.94) mg·m-2·h-1。移植 1 年的红树林与移植5年的红树林在CO2碳通量存在极显著差异性(P<0.01)。随着红树林幼苗成长,吸收和固定大气中 CO2能力加强,可能发挥更大的碳汇作用。林龄是影响红树林碳汇功能的重要影响因素。
2.3 不同林龄红树林群落CH4和CO2通量日变化
整个观测期内,CH4通量日变化在移植5年与1年的红树林存在明显差异。移植5年的高潮滩与低潮滩红树林CH4通量的日变化整体呈现“M”型(图3),CH4排放通量在06:00—08:00出现第一个排放最大值点,在 11:00—12:00,CH4排放通量出现最低值。正午之后,CH4通量逐渐增加。其中,高潮滩的红树林植被 CH4排放量的最大值出现在06:00,低潮滩红树林群落最大值出现在 07:00。移植5年红树林的CH4通量时隔半天出现两个峰值,同时本研究区为半日潮区,潮水变化可能是影响红树林CH4通量的重要因素。移植1年的红树林日变化趋势较为稳定,最大值出现在14:00,04:00出现最小值。另外发现,同一红树林植被9月与10月CH4通量日变化存在差异。移植5年的红树林(高潮滩与低潮滩)在10月的日变化CH4通量明显高于9月,但移植1年的红树林表现为降低,CH4通量日变化出现季节性差异。
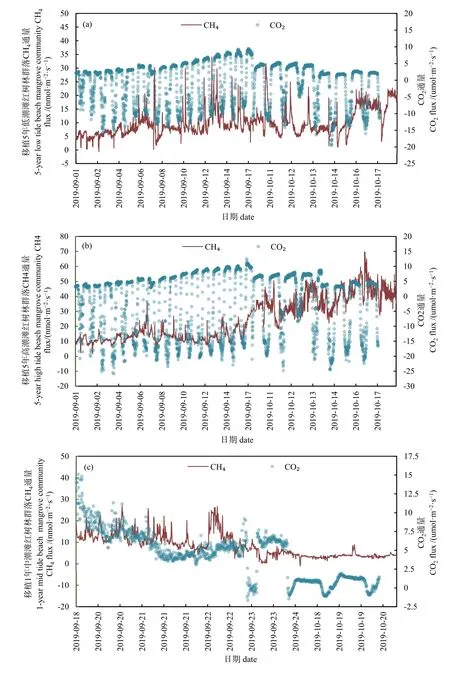
图1 红树林移植5年低潮滩(a)和高潮滩(b)以及移植1年中潮位(c)CH4和CO2排放通量变化趋势Fig. 1 CH4 and CO2 emission fluxes trend of 5-year mangroves at low tide beach (a), high tide beach (b) and 1-year at mid tide beach (c)

图2 不同林龄红树林CH4和CO2通量对比Fig. 2 Comparisons of CH4 and CO2 fluxes of mangroves in different ages

图3 2019年9月和10月不同林龄红树林CH4排放通量日变化Fig. 3 Diurnal variation of CH4 emissions of mangroves in different ages in September and October 2019
移植5年的红树林的CO2通量日变化呈现“U”型变化趋势(图 4)。观测期内每天 19:00至次日07:00,CO2排放以群落呼吸为主,释放CO2;受红树林光合作用影响,白天光照条件下CO2吸收能力高于排放能力;移植5年红树林恢复区白天为CO2的“汇”,夜晚为CO2的“源”。移植1年的红树林的9月与10月的CO2通量日变化有明显差异:在9月观测期,由于幼苗光合作用强度较弱,以呼吸作用为主,11:00—12:00,CO2通量出现最大值,为CO2的“源”。经过9月一个月的生长期,移植1年的红树林在10月的CO2通量日变化,同5年的红树林一样,呈现“U”型变化趋势,在07:00—16:00,CO2通量为负值,表现为碳汇作用。
3 讨论
3.1 红树林生长状况对温室气体排放的影响
移植5年红树林植被植株高度可达160 cm,根茎深度达25 cm,根系发达。移植1年红树林植株平均高度为37 cm,冠幅平均为23 cm,根茎深度为8 cm,根系较小(图5)。移植5年的红树林的CO2与CH4两种气体温室气体通量均大于移植1年的红树林(图2)。由红树林的样方调查结果可知,移植5年的红树林植株高度与冠幅明显高于移植1年的红树林。生物量、根系发达程度与凋落物随林龄呈正相关关系。年长的红树林植被生物量明显较大,植被根系的分泌作用为甲烷菌提供充足的底物(陈海生等,2017;黄国宏等,2001),植物的生物量与CH4排放量之间存在明显的正相关性(Whiting,1991)。红树林土壤中的有机质含量与凋落物的碳储量随林龄增长而增大(金亮等,2017),提供充足的甲烷基质。另外,土壤中植被根系附近产生的CH4通过植株的通气组织向进行释放,林龄越大的红树林的通气组织越发达,CH4通量自然也就更高。

图4 2019年9月和10月不同林龄红树林CO2排放通量日变化Fig.4 Diurnal variation of CO2 emissions of mangroves in different ages in September and October 2019
红树林群落的CO2由植被的光合作用和群落的呼吸作用组成。移植5年的红树林植被植株个体较大,在白天进行光合作用,吸收的CO2远高于移植1年的幼苗。在夜间,植被不再进行光合作用,群落只进行呼吸作用。土壤中的有机质含量与凋落物层的碳储量与林龄呈正相关性,同时年长的红树林根系组织较发达。因此,呼吸作用产生的CO2相比较移植1年的红树林更多(图4、5)。
3.2 潮位和土壤属性对温室气体排放的影响
移植5年的高潮滩红树林CH4通量高于移植5年的低潮滩通量,高潮水位比低潮滩水位较低,但CH4通量高于低潮滩(图2)。已有研究表明,CH4通量与水位无正相关性(叶勇等,2000a)。潮水的水位高度不等同于土壤的含水量,潮水上涨的短暂效应不会增加CH4通量(卢昌义等,2000;叶勇等,2000a)。潮汐通过影响土壤因子,如土壤有机碳、盐度与pH等(图5),影响CH4的产生率。红树林植被生物量、土壤碳密度均随着潮滩高程的增加而增加(陈瑶瑶等,2019)。大多数研究已表明,土壤丰富的有机质含量,提供了产生CH4的充足分解底物,同时有机质也有利于增加甲烷菌的数量(姜欢欢等,2012)。本研究发现,垂直深度20—40 cm土壤有机碳含量出现最大值,与海南红树林土壤有机碳分布规律一致(辛琨等,2014)。高潮滩的红树林土壤的有机质含量最高,与CH4排放通量变化趋势一致(图2和图6)。
红树林湿地作为海岸带湿地,潮水中含有一定量的盐分,盐分通过影响甲烷菌的活性进一步影响CH4通量。研究表明,CH4通量与盐度呈反比(Sun et al.,2013;李杨杰等,2014),随盐度的增加CH4通量降低。有学者的研究发现,盐度超过一定范围(盐度>18×10-3)时,沼泽的CH4通量明显受到抑制(Poffenbarger et al.,2011)。已有研究表明红树林土壤的pH与盐度呈正相关关系(沙聪等,2018),CH4排放量较低的土壤pH相对较高。SO42-与CH4通量呈负相关关系(叶勇等,2000b;王纯等,2015),本研究结果发现在土壤 20 cm处,高潮滩土壤的SO42-浓度最低(图6)。潮水淹没期间土壤含水量会发生变化,在水分蒸发、消退的过程中会引起土壤温度的变化,一定程度上会间接影响土壤中微生物的活性,从而改变CH4通量。在CH4传输过程中低潮滩有较高的水位,形成一道“屏障”阻断了CH4的排放通道。另外,土壤排放的CH4通量一部分会被潮水氧化和分解(卢昌义等,2000)。潮汐对CH4通量的影响主要通过对其产生率与传输两个方面进行影响。本文研究CH4通量结果发现移植5年的高潮滩红树林>移植5年的低潮滩红树林>移植1年的中潮滩红树林,其中移植1年的红树林处于低潮滩和高潮滩之间,但通量明显较低,由此推论,除潮位梯度因素之外有其他因素共同影响CH4通量。
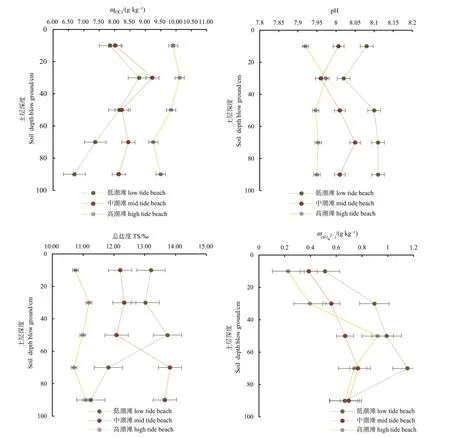
图6 红树林恢复区不同滩面土壤理化特征的不同土层深度变化Fig. 6 Variations of soil physical and chemical characteristics with different soil depths at different beaches in mangrove ecological restoration zone
红树林群落CO2通量由地上的植被与土壤呼吸共同构成。在白天,红树林群落以光合作用为主,吸收大气中的 CO2,夜晚只进行呼吸作用,释放CO2。本研究采用的透明同化箱观测,涨潮期间,箱内潮水不能淹没红树林植株叶片。潮水主要通过群落的土壤呼吸间接影响红树林群落的CO2通量。本研究结果发现,红树林群落的CO2通量表现为移植5年的高潮滩红树林>低潮滩红树林>移植1年的红树林。有研究表明,潮汐淹没对红树林土壤的CO2排放具有显着影响,高潮滩红树林CO2通量明显高于低潮滩红树林(Chen et al.,2010)。第一,分子在气体的扩散速度快于流体,低潮滩的水位较高,不利于气体的扩散,例如,当水位降低时,日本中尾湖的沿海盐沼,CO2通量趋于增加(Hirota et al.,2007)。第二,在潮水淹没后,形成厌氧环境,氧化还原电位相对更低,土壤中微生物有氧呼吸和有机质的化学氧化作用减弱,进而导致 CO2通量降低(Adam,2011)。第三,潮汐通过潮水的动力,以物理转移的方式影响不同潮位的有机质积累。有研究表明,潮汐动力能够转移低潮滩植被的生物量可高达75%(Cranford et al.,1989),在潮汐的转移作用下,低潮滩提供给土壤可利用的有机质相对高潮滩少,CO2通量相对较低。综上,潮位梯度主要通过周期性的水位高低变化、土壤氧化电位等因子进而影响气体通量的变化。潮水对植被温室气体通量的影响,本质上是由于潮汐改变了不同梯度下的环境因子,例如水位高度和土壤因子等,形成环境梯度差异,进而影响植被不同的温室气体通量大小(郭海强等,2010)。
4 结论
海岸带生态恢复区不同林龄的红树群落均为CH4排放“源”,移植5年的红树林的CH4排放通量高于移植1年红树林。受不同林龄植被红树林光合作用影响,移植5年红树林为大气CO2吸收汇,而移植1年红树林为大气CO2排放源。
土壤属性与潮汐淹水是影响红树林恢复区CH4和CO2产生排放的重要因子。受潮汐淹水的影响,红树林恢复区土壤属性存在较为明显的梯度变化。由靠近江水的滩涂到岸边,土壤有机碳含量呈现升高的趋势,土壤pH、土壤总盐度和土壤SO42-含量呈现降低的趋势。移植5年红树林高滩位CH4排放通量和CO2吸收能力显著高于低滩位。
