苗期高温高湿影响番茄花芽分化进程的机理探讨*
2021-01-20黄琴琴杨再强刘显男王学林丁宇晖李佳佳郑芊彤
黄琴琴,杨再强,2**,刘显男,王学林,徐 超,丁宇晖,李佳佳,郑芊彤
苗期高温高湿影响番茄花芽分化进程的机理探讨*
黄琴琴1,杨再强1,2**,刘显男1,王学林3,徐 超1,丁宇晖1,李佳佳1,郑芊彤1
(1. 南京信息工程大学气象灾害预报预警与评估协同创新中心,南京 210044;2. 江苏省农业气象重点实验室,南京 210044,3. 合肥市气象局,合肥 230041)
以番茄品种“寿和粉冠”为试材,于2020年4−7月在南京信息工程大学农业气象试验站进行气温、空气相对湿度、处理天数的正交试验,气温(昼温/夜温)设4个处理水平:T1(32℃/22℃)、T2(35℃/25℃)、T3(38℃/28℃)和T4(41℃/31℃);空气湿度设3个处理水平:H1(50%)、H2(70%)和H3(90%),误差范围在±5个百分点;处理天数为2、4、6和8d。以昼温/夜温28℃/18℃、空气相对湿度45%~55%处理为对照(CK)。在番茄花芽分化各个时期分别测量顶芽内源激素、淀粉和可溶性糖含量,在现蕾期测量茎粗、单株干质量、壮苗指数和叶绿素含量,以研究苗期高温高湿影响番茄花芽分化进程的机理。结果表明:(1)随着温度升高,整个花芽分化过程随着温度的升高而延长,而空气相对湿度和处理天数对番茄花芽分化进程影响不大。(2)不同处理下番茄顶芽IAA和GA3含量随着花芽分化出现降—升—降的趋势,ZT和ABA含量出现与IAA完全相反的趋势。IAA、ZT、GA3含量均随着温度、相对湿度和处理天数的增加逐渐降低,ABA含量随着胁迫程度的增加逐渐升高。(3)番茄叶片淀粉和叶绿素含量随花芽分化进程逐渐降低,可溶性糖含量从未分化期到雄蕊分化期逐渐升高,雌蕊分化期间逐渐降低。随着胁迫程度的加深,各处理间差异显著。表明高温高湿对番茄花芽分化的抑制作用可能与内源激素含量变化、营养物质减少有关,花芽分化初期环境温度应控制在CK水平,温度越高越不利于番茄花芽分化。
番茄;高温;高湿;内源激素;花芽分化进程;营养物质
番茄()是一种富含维生素的蔬菜,其高含量的胡萝卜素对心脑血管具有较好的保护作用,营养价值和口感均是一般蔬菜无法比拟的[1]。番茄的消费,不仅在量上呈逐年增加的趋势,市场对其品质的要求也越来越高[2−4]。番茄在生长过程中喜欢日照,对温度敏感,尤其在营养生长阶段,其最佳生长气温在15~32℃[ 5−7]。日光温室是中国冬季蔬果进行反季节生产的主要设施类型,在不同日光温室温度条件下番茄的生长发育有显著差异。夜间温度低会影响根系发育,不利于植株正常生长和光合作用[8−10]。日间高温常导致设施番茄死棵,高温干旱条件下番茄花芽分化不良,从而导致开花坐果不良,还会诱发病毒病等[11−12]。番茄花芽分化不良是畸形番茄花果形成的主要原因,在花芽分化过程中若遭到不适宜的温度,导致花芽分化障碍,在花芽形成以后就处于畸形状态,最后形成不正常的花或果实[13]。大量研究表明,植物的花芽分化主要与植物体内内源激素有关,对于一些农作物和花卉,内源激素的分泌直接影响植物的花芽分化进程[14]。
Ahamed等[15]研究指出,高温抑制花芽分化进程。Bagamboula等[16]研究表明,在葡萄花芽分化前21d,植株对外界的温度极为敏感,在形成葡萄原基的前21d,30℃左右是最适宜的温度。Pense等[17]研究了桃花芽形成和花朵发育过程中涉及的内源性和外源性因素,表明桃的花芽形成过程在低温和温暖条件下有所不同。温暖条件下的芽诱导和分化是解决花蕾发育变化的重要基础。Pornpairin[18]实验得出,球芽甘蓝花芽分化有较低的温度要求,低温对甘蓝花芽分化的影响具有品种间差异。Hidaka等[19]研究了冠冕降温处理对6月草莓的花芽分化、开花特性和果实产量的影响,表明在适当的时期进行冠冕冷却处理能够在高温下稳定草莓的产量。杨丽等[20]研究表明,国庆小菊花花芽分化的主要影响因素是外界温度。袁慧敏等[21]研究指出,番茄的花芽分化及发育对温度有一定要求,白天气温20~25℃,夜晚15~17℃是番茄最佳的花芽分化温度,这种条件下分化的花芽质量好,开花数量多。赵玉芬等[22]研究指出,夜间温度的高低决定八仙花花芽分化完全与否,并直接影响开花质量。
目前关于高温单一因子对番茄生长发育和产量形成的影响研究较多,主要集中在植株生长、光合荧光特性、保护酶活性和可溶性糖等方面[23−25],而关于高温高湿复合胁迫对番茄花芽分化影响仍然缺乏系统性的研究。因此,如何调控设施小气候,优化番茄花芽分化是亟待解决的技术问题。本研究拟通过人工环境控制试验,研究不同温度对番茄花芽分化速率、内源激素、糖含量及叶绿素含量的影响,揭示番茄植株花芽分化机理,以期为设施番茄种植过程中温湿度的优化调控提供理论依据和实践指导。
1 材料与方法
1.1 试验设计
以番茄品种“寿和粉冠”为试材,在南京信息工程大学农业试验站玻璃温室(Venlo型)内进行人工控制试验。2020年3月16日使用基质土在温室(22~28℃、45%~55%)内苗床上育苗,秧苗处于二叶一心时(4月25日)定植至28cm(高)×25cm(底径)的花盆中,每盆一株。待秧苗长到四叶一心(4月30日)时选取长势相近的植株放入人工气候箱(TPG1260,Australia)进行不同温湿度组合处理。正交试验因素为气温和空气相对湿度,试验设计见表1。其中日最高气温/最低气温设置为32℃/22℃、35℃/25℃、38℃/28℃和41℃/31℃共4个水平,相对湿度设置50%、70%和90%共3个水平(误差范围±5个百分点),持续处理时间为2、4、6和8d,对照组温度设置为28℃/18℃,空气相对湿度为45%~55%。
番茄花芽分化等级确定参考《中国蔬菜栽培学》(表2)[26],即未分化期(US)、分化初期(ES)、萼片分化期(SS)、雄蕊分化期(XS)和雌蕊分化期(PS),同时分别在番茄幼苗花芽分化的5个时期测量顶芽内源激素、淀粉和可溶性糖的含量,在现蕾期测量茎粗、单株干质量、壮苗指数和叶绿素含量。南京地区玻璃温室内的夏季气温日内变化过程采用神经网络方法模拟[27],试验期间保证盆栽土壤的水分和养分条件均维持在适宜水平。各处理指标值均为3次重复的算术平均值,试验数据参照韦婷婷等[27]正交试验数据处理方法,求出各水平指标的均值并进行比较。
1.2 测定项目与方法
1.2.1 花芽分化进程的测定
从处理开始至结束,每隔两天取各处理10株番茄苗,进行花芽分化调查,在Olympus电子显微镜下解剖并观察其生长锥,使用Mshot MS60拍照,统计花芽分化时期,得出每隔两天的番茄花芽分化情况。记录花芽分化每个阶段开始和结束的日期。各分化阶段的确定以达到某一时期的植株超过3/5即6株为标准。

表1 人工气候箱高温高湿环境的正交试验方案L16(42×3)

表2 番茄花芽分化阶段判断标准
由图1可见,处于未分化期的番茄茎尖顶端未见明显分生组织(图1a);分化初期的顶端分生组织稍膨大,顶端分生组织明显可见(图1b);当观测到顶花芽四周萼片原基开始隆起时即为萼片形成期(图1c);雄蕊形成期的萼片原基的内侧开始形成齿状的雄蕊原基,番茄顶芽进一步隆起(图1d);雌蕊形成期的花芽中心部分进一步隆起,在电镜解剖图中可以看到中心隆起部分开始出现数个突起的小圆丘,即雌蕊心皮原基(图1e)。

(a)未分化期Undifferentiated period;(b)分化初期Early period of differentiation;(c)萼片分化期Sepal differentiation period;(d)雄蕊分化期Stamen differentiation period;(e)雌蕊分化期Pistil differentiation period
注:图中7−16为番茄叶序。
Note:7−16 in the figure are the tomato leaf sequence.
1.2.2 顶芽内源激素及糖含量的测定
取下顶芽用Olympus电子显微镜(南京产,型号SZX16)下观察花芽分化后,放入液氮罐进行速冻,随后放入−80℃的冰箱内冷藏。测定样品中的生长素(IAA)、赤霉素(GA3)、细胞分裂素(CTK)和脱落酸(ABA),共4种内源激素。采用韩佩汝等[28]的方法测定内源激素,采用蒽酮比色法测定淀粉和可溶性糖含量[29]。
1.2.3 叶绿素含量的测定
采用乙醇提取比色法。用电子天平称取0.2g叶片(选取番茄第7片真叶),将样品置于25mL95%乙醇中,放置48h直至叶片中的细胞色素完全被提取出,把叶绿体色素提取液倒入光径1cm的比色杯内。以95%乙醇为空白,用722nm分光光度计,在波长665nm、649nm下测定吸光度。各色素浓度为:Ca=13.95×D665−6.99×D649,Cb=24.96×D649− 7.32×D665。
1.3 数据统计与分析
采用SPSS23.0和Microsoft Excel软件对数据进行统计分析。运用Duncan检验法进行多重比较和差异显著性检验(P=0.05),用Microsoft Excel软件作图。数据以平均值±标准误差表示。
2 结果与分析
2.1 高温高湿对番茄花芽形态分化的影响
由图2可见,对照植株于处理开始后7d即5月7日开始花芽分化,此时取10株番茄苗样观察,其中8株处于分化初期,分化概率为80%,分化初期历时6d,萼片、雄蕊、雌蕊分化期分别历时10、6和8d,从观测日起历时30d。图2a显示,与CK相比,各高温处理水平下花芽开始分化的时间均有不同程度的延迟,32℃处理水平下于处理开始后32d进入分化初期,35℃处理水平下于38d后开始分化,38℃和41℃处理水平下分别于42d和48d后开始分化,分别比CK晚35d和41d;各高温处理水平下花芽分化到雌蕊分化总历时44、56、66和78d,分别比CK延长14、26、36及48d。可见,日最高温度高于32℃条件会使番茄幼苗花芽分化明显延迟,分化进程明显延长,而且温度越高,延迟或延长的程度越重。
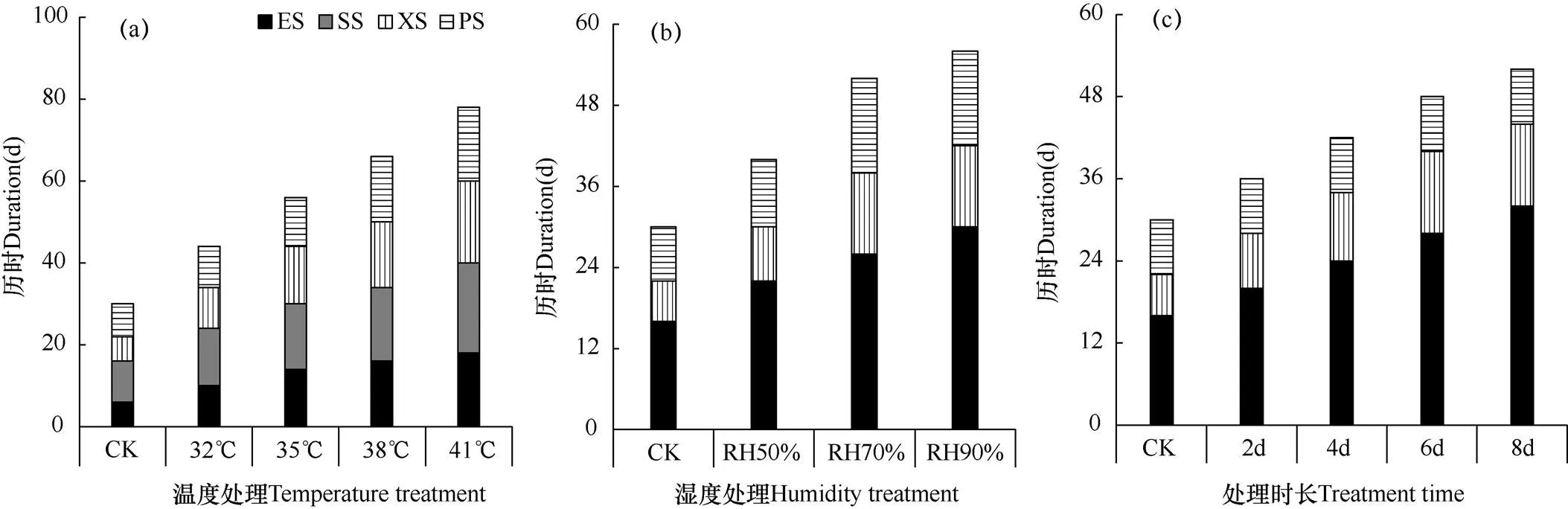
图2 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗花芽各分化阶段历时的比较Fig. 2 Comparison of the duration of flower bud differentiation of tomato seedlings under different high temperature (a), high humidity (b) and treatment time (c)
注:US:花芽未分化期,ES:分化初期,SS:萼片分化期,XS:雄蕊分化期,PS:雌蕊分化期。数据为平均值。下同。
Note: US: undifferentiated stage, ES: early differentiation stage, SS: sepal differentiation stage, XS: stamen differentiation stage, PS: pistil differentiation stage. The data are average values. The same as below.
图2b显示,与CK相比,各高空气湿度处理水平下花芽分化时间均有不同程度的延迟,RH50%处理水平下于处理开始后16d进入分化初期,RH70%处理水平下于22d后开始分化,RH90%处理水平下于26d后开始分化,分别比CK晚9、17和21d。各高湿度处理水平下花芽分化到雌蕊分化分别历时40、52和56d。可见,空气相对湿度高于45%会使番茄花芽分化进程明显延长与延迟。湿度越高,延迟或延长程度越重。
由图2c可见,与CK相比,各处理时长下花芽分化时间均有不同程度的延迟,2d处理水平下于处理开始后12d进入分化初期,4d处理水平下于18d后开始分化,6d处理水平下于24d后开始分化,8d处理水平下在26d后开始分化,分别比CK晚7、17和19d。2、4、6和8d处理时长下花芽分化到雌蕊分化分别历时36、42、48和52d。可见,处理时间越长,花芽分化延迟或延长程度越重。
2.2 高温高湿对番茄花芽顶芽内源激素的影响
生长素(Auxin,IAA)为首个被证实存在的植物内源激素,生长素参与调控植物几乎所有的生长发育进程,由图3a可看出不同高温处理对番茄花芽分化各阶段IAA含量的影响。32℃处理下,IAA含量与CK相比差异不显著,35℃以及更高的温度处理下,IAA含量显著降低,当环境温度为41℃时,IAA含量相对于CK下降了48.2%。在各处理中,随着处理时间的延长,IAA含量均出现先降后升的趋势,分化初期IAA含量最低,雄蕊分化期IAA含量达到峰值,这说明分化初期低水平的IAA有利于番茄花芽由营养生长向生殖生长转变。
由图3b可看出,湿度对番茄顶芽内源激素的影响不如高温处理显著。在50%空气湿度处理下,IAA含量与CK差异不显著,当空气相对湿度上升到70%时,IAA含量下降25.7%。相对湿度为90%时,IAA含量相对于RH70%处理有所上升,相比于CK下降24.1%。各处理水平下,番茄幼苗在未分化期IAA含量均保持较高水平,IAA含量最低值出现在分化初期,花芽分化阶段内呈降—升—降的变化趋势,说明低水平的IAA有助于番茄由营养生长向生殖生长转变,高水平的IAA有利于花芽的形态分化,特别是番茄雌蕊的分化。
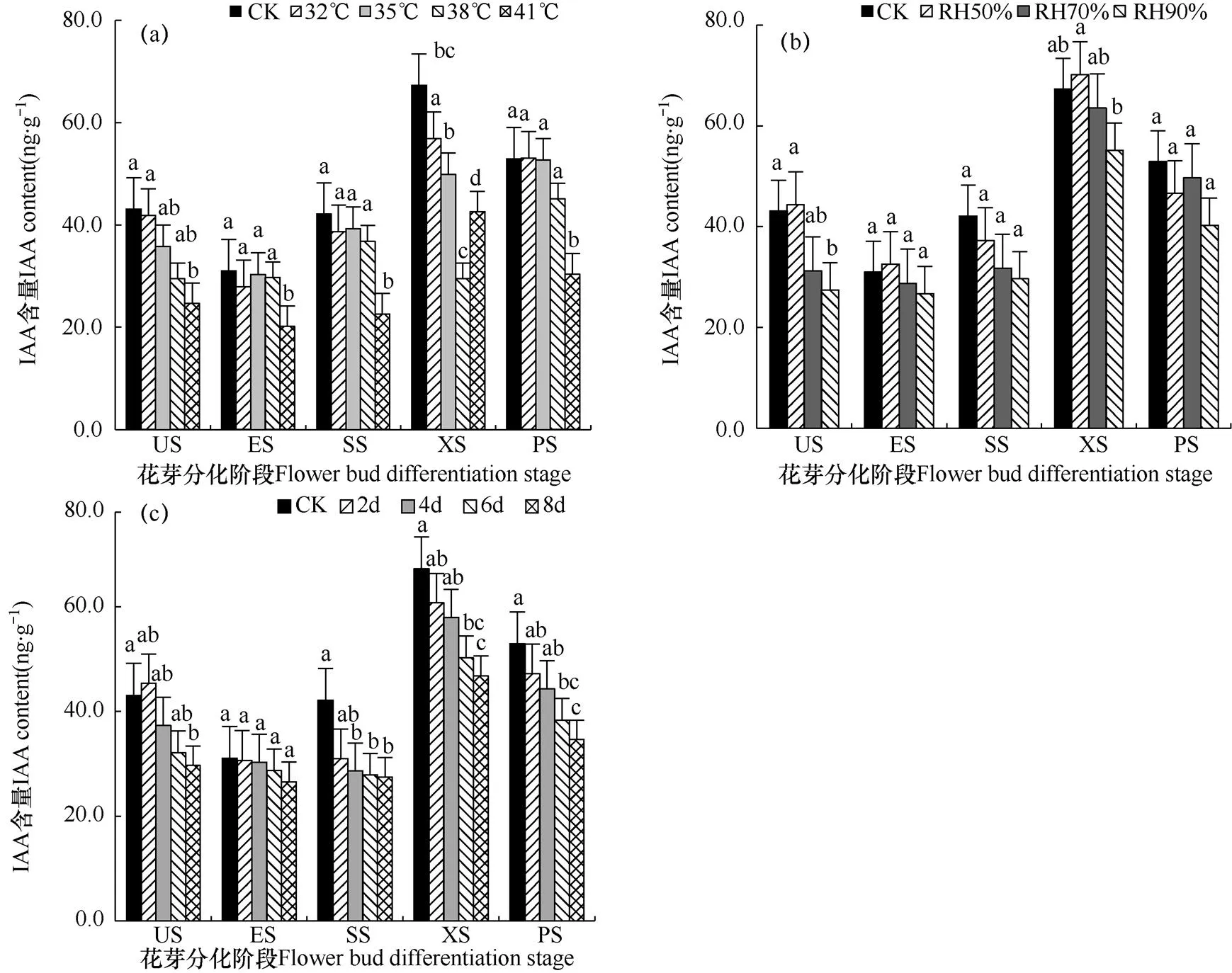
图3 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗顶芽花芽分化过程中生长素(IAA)含量的比较
注:小写字母表示处理间在0.05水平上的差异显著性,短线为标准误差。下同。
Note:Lowercase indicates the difference significance among treatments at 0.05 level. The short line is the standard error bar. The same as below.
由图3c可见,IAA含量在未分化阶段和分化初期各处理均与CK差异不显著。在萼片、雄蕊和雌蕊分化期各处理均与CK差异显著。在花芽分化阶段内IAA含量整体呈现降—升—降的趋势。
细胞分裂素(Cytokinin,CTK)在植株内分布较广,生理功能多样,比如引起细胞分裂、诱导芽的形成、防止离体叶片衰老等。反玉米素(ZT)作为细胞分裂素的一种,在植物体内分布最广,是一种具有较高活性的细胞分裂素,对植物生长发育起重要作用。由图4a可看出,高温处理对番茄顶芽ZT含量影响不显著,当温度上升到38℃和41℃,ZT含量分别比CK下降了26.0%和67.7%。CK处理下ZT含量均显著高于3种高空气湿度处理,而各湿度处理之间ZT含量差异不显著。对于各处理,ZT含量最低值均出现在US阶段,最大值均出现在ES阶段,花芽分化阶段内呈现升—降变化趋势。说明低水平的ZT促进花芽分化的开始,高水平的ZT有利于番茄花芽形态的转变,特别是促进分化初向萼片分化。
脱落酸(Abscisic acid,ABA)是植物生长抑制型激素,有助于碳水化合物向库的运输,高含量ABA易造成芽休眠、落叶等。由图5a可看出,各处理ABA含量随着温度升高而升高,各高温处理ABA含量均显著高于CK,可见高温诱导植物内源ABA的大量合成。由图5b可看出,在50%相对湿度处理下ABA含量最高,为0.62μg·g−1,而CK处理下含量最低,说明空气湿度增加会显著提高番茄顶芽中ABA含量。从各花芽分化阶段看,ABA最低值出现在XS阶段,最高值出现在ES阶段,呈现出升—降—升的趋势,说明高水平的ABA能促进番茄花芽从分化初期向萼片分化期转变,而在雄蕊分化期顶芽较低含量的ZT有利于雌蕊的分化。
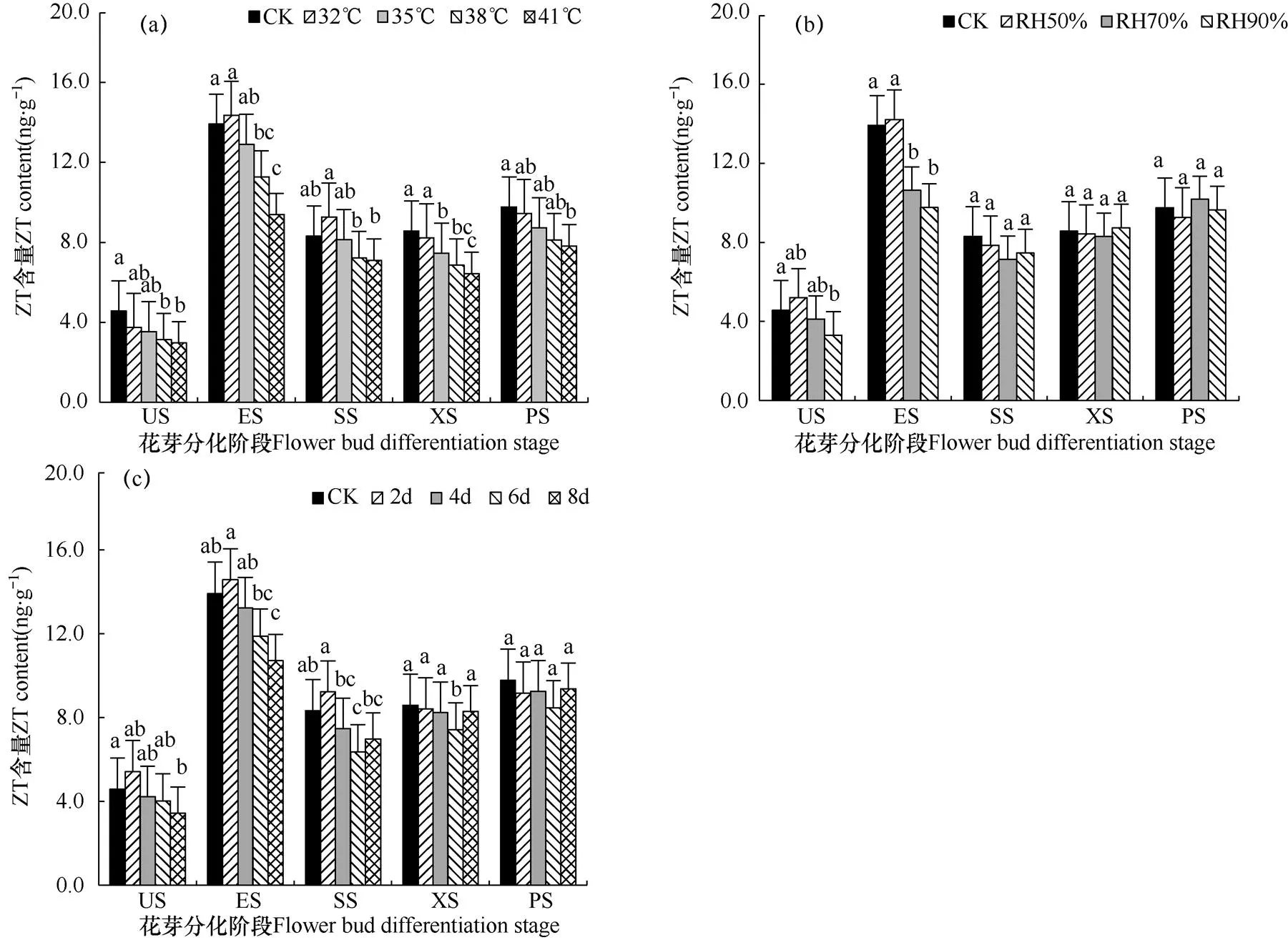
图4 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗顶芽花芽分化过程中顶芽反玉米素(ZT)含量的比较
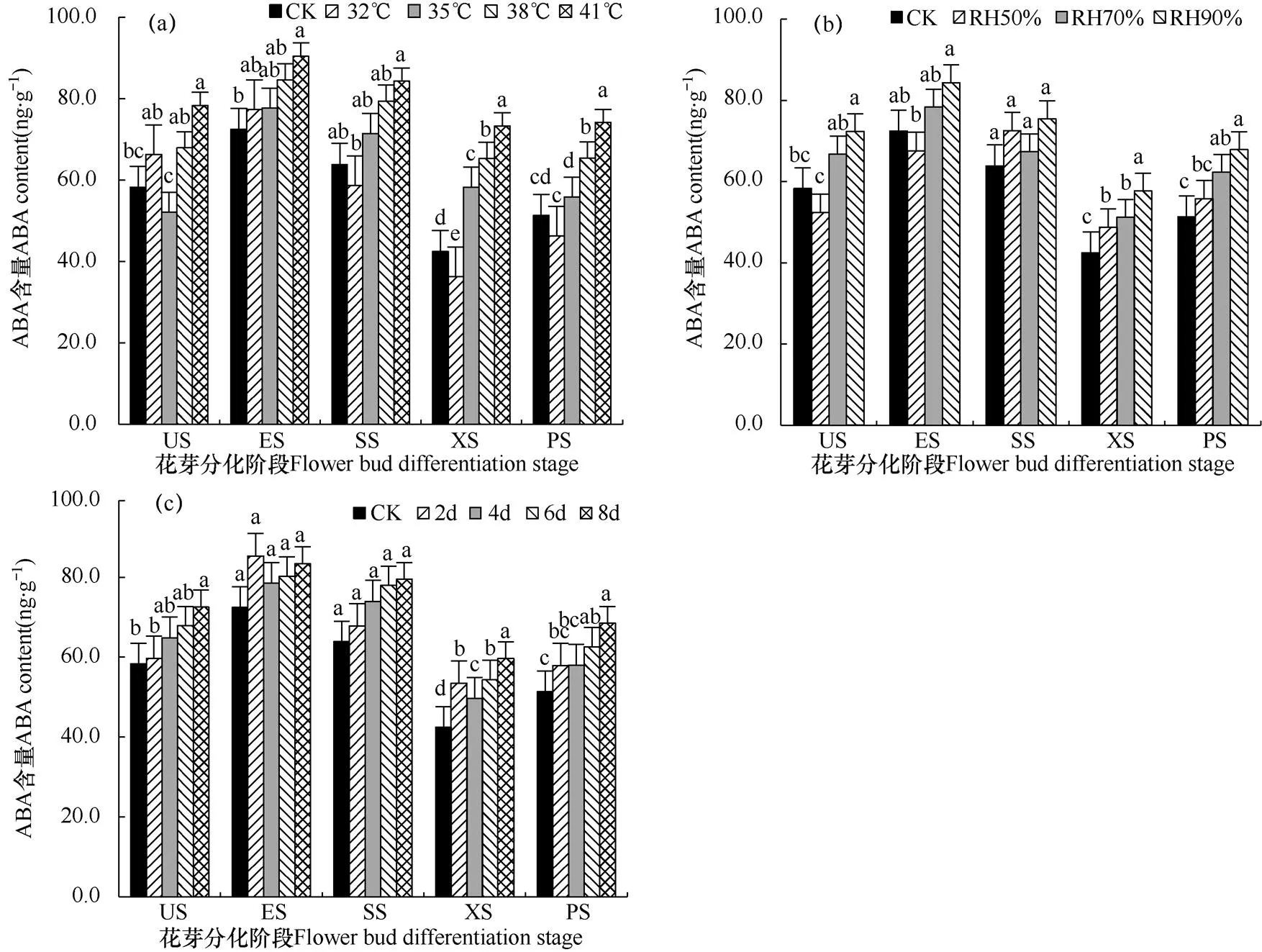
图5 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗花芽分化过程脱落酸(ABA)含量的比较
赤霉素(Gibberellicacid,GA3)多存在于生长旺盛的植物器官内,其主要生理作用是促进茎、叶的伸长,还可以调控IAA的水平,诱导植物开花。从图6a可看出,32℃处理下,GA3含量与CK 处理差异不显著,随着温度持续升高,GA3含量显著下降,35、38和41℃下GA3含量分别比CK下降10.17%、21.3%和33.8%。由图6b可看出,随着空气湿度的增加,GA3含量逐渐降低。随着花芽各等级分化,GA3含量呈现降—升—降的趋势,与IAA、ZT含量变化趋势类似,说明形态分化开始后,高水平的GA3有利于各个器官的分化和发育。
2.3 高温高湿对番茄幼苗花芽分化过程中糖及叶绿素含量的影响
如图7所示,随着温度、湿度和处理天数的增加,番茄幼苗淀粉含量呈现直线下降趋势,最大值出现在CK处理下的US阶段,为131mg·g−1,最低值为7d处理下的PS阶段,为35mg·g−1。可溶性糖随着温湿度的增加,呈现单峰曲线变化趋势,均为CK处理在XS阶段达到最大值,为99mg·g−1。最低值为41℃处理下的US,为17mg·g−1。由表3可看出,不同高温高湿处理对番茄幼苗叶绿素含量的影响不同。光合色素以叶绿素a为主,含量在4.40~10.37mg·g−1,叶绿素b含量为2.06~3.52mg·g−1,叶绿素a的降低引起叶绿素b的变动。高温高湿胁迫下番茄叶片中叶绿素a、叶绿素b、叶绿素总量均低于对照。空气湿度对叶绿素a和叶绿素b含量影响显著,但各高温处理之间叶绿素含量差异不显著,高温对叶绿素a含量影响较大,在41℃处理下,叶绿素a含量相对于CK下降了71.6%。高温对叶绿素b含量影响较小,从而导致叶绿素a/b降低。降低程度与叶绿素a一致。说明高温高湿胁迫引起番茄幼苗叶绿素a、叶绿素b和叶绿素总含量下降程度与胁迫程度成正比。
糖类是番茄花芽分化及发育的能量基础,高水平可溶性糖促进番茄花芽分化和成花。而高温高湿条件抑制了番茄光合色素的合成,抑制了光合作用,使其呼吸作用增强,叶片气孔阻力增加,碳水化合物的积累导致番茄中淀粉和可溶性糖含量显著下降,不利于番茄花芽分化的能量储备,从而使番茄幼苗地上部分营养生长受阻,生殖生长必然受到影响,表现为花芽分化延迟和分化进程延长。

图6 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗花芽分化进程中GA3含量的比较
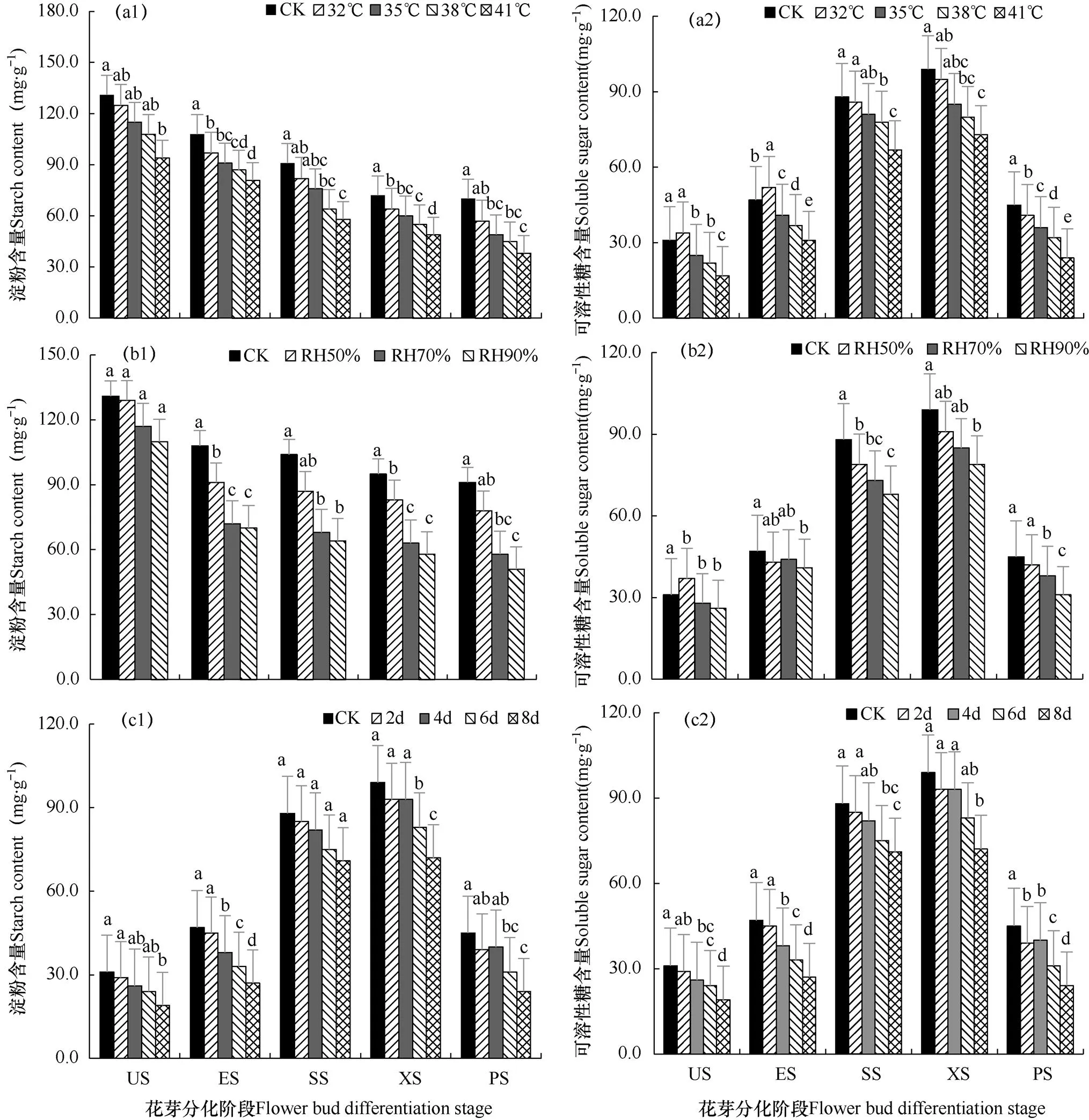
图7 不同高温(a)、高湿(b)和处理时长(c)下番茄幼苗花芽分化期淀粉(1)和可溶性糖(2)含量的比较

表3 不同处理番茄幼苗叶绿素含量的比较
3 结论与讨论
3.1 结论
在番茄品种“寿和粉冠”苗期花芽分化过程中,高温高湿复合影响下,植株花芽分化开始日期均有不同程度的推迟,整个花芽分化进程也会不同程度延长,高温高湿影响时间越长,花芽分化推迟和延长越明显。
花芽分化过程中,高温高湿复合影响下,花芽顶芽内源激素生长素(IAA)、赤霉素(GA3)、细胞分裂素(CTK)和脱落酸(ABA)也发生明显变化,IAA、ZT和GA3含量均明显减少,而ABA含量明显增加,抑制了花芽分化和进程。
高温高湿环境影响下,番茄植株体内糖和叶绿素含量均明显减少,影响光合作用进行,从而使有机物质合成不足,抑制了番茄植株地上部分营养生长与生殖生长,从而导致花芽分化进程延迟与延长。
3.2 讨论
关于高温高湿对番茄花芽分化速率的研究,目前认为植物处于高温高湿逆境时,植株叶片气孔关闭频率较张开频率高,植株进行光合作用和蒸腾作用速率下降,植物呼吸作用增强,植物体内光合色素合成量减少,最终使光合作用能力下降,淀粉及可溶性糖等营养物质含量下降,植物体内能量储备减少,生殖器官中同化物分配量减少,植物发育关键阶段的可利用营养物质短缺,致使生殖生长处于饥饿状态,最终导致花芽分化速率减慢[30−31]。本实验中,导致花芽分化速率减慢可能原因是即将进入生殖生长阶段的番茄幼苗遭受高温高湿胁迫后,生殖器官中碳水化合物供应不足,生殖生长长期处于饥饿状态下,最终导致高温高湿处理下的花芽分化各个阶段的分化速率相比对照显著减慢,这与聂梅等[32]研究的高温对地被菊“紫娇”花芽分化影响的结论一致。
大部分学者认为IAA是花芽形成的抑制因子之一,但也有学者研究发现高水平IAA抑制植株花芽分化可能与植物种类有关,在本研究中对番茄品种“寿和粉冠”而言,高水平IAA有利于分化初期花芽分化的启动,萼片、雄蕊和雌蕊分化期花芽形态的膨大和细胞的增殖。而在高温和高湿度条件下,番茄中IAA的形成会逐渐减少,说明高温高湿环境对番茄初期花芽分化的启动,后期萼片、雄蕊和雌蕊分化都具有抑制作用。
对于不同的植物种类和品种,ZT的调控作用也有所不同,对本实验中的番茄品种“寿和粉冠”而言,高水平的ZT有利于番茄花芽的分化,属于促花激素,在对邓恩桉[33]和油茶花芽分化[34]的研究中有相似的结果。而在高温高湿条件下,番茄顶芽中ZT的生成受到抑制,温湿度越大,ZT含量越少。
ABA作为重要的植物生长调节剂,在植物响应非生物胁迫时也扮演着重要的作用,且在植物的器官中一直存在。大量研究证实,在成花过程中ABA可能从两个方面进行调控:(1)与IAA、GA3产生协同作用,促进营养物质在花芽中积累,促进成花。(2)诱导花芽休眠,抑制成花[35−36]。ABA在番茄花芽分化过程中的变化,与赵通等[37]的研究结果相似,高水平的ABA能够抑制顶端优势,促进花芽分化,低水平ABA有利于花芽分化后期各花器官的形态分化和发育,避免休眠的诱导。在番茄顶芽中,ABA含量在高温高湿环境下分泌量减少,削弱了ABA对花芽分化的促进和抑制作用。
植株顶芽中GA3与IAA通常对植物产生协同作用,在植物中共同生成与消亡。这一结论可在艾星梅等研究马铃薯花芽分化与内源激素动态关系[38]的结果中得以证实。本实验中结果与王磊等[35]结论一致,即GA3含量随着环境温湿度的上升而下降。
高温高湿胁迫下番茄幼苗的糖及叶绿素含量均显著低于对照。高温作为影响植物生长发育的常见非生物胁迫之一,高温胁迫下诸多农艺性状都予以响应,高温环境会影响植物光合作用,进而影响植物干物质的积累,植物需要消耗自身营养物质来抵御高温伤害,表现为生长缓慢,甚至停滞。高温会使植株徒长,通过增加空气湿度可以在植物的某个特定时期起到缓解高温胁迫的作用,但是在植物的整个生长发育过程中,加湿的影响微乎其微,并且超过一定的湿度,不仅不能缓解高温胁迫,反而不利于植株的生长发育。总体来说,随着温度的升高,番茄营养物质均呈下降趋势,这与张海娜等[39]在其它植物上的研究结果一致。Silvia等[40]认为高温严重影响了水稻的生长形态,降低了植株叶面积、地上和地下生物量;Semenov等[41]研究认为,高温导致植物生理指标受阻,生长迟缓。本实验研究结果表明,高温胁迫使番茄幼苗茎粗、单株干质量和壮苗指数下降,植株徒长,这与Figueroa等[42]研究中高温阻碍植物正常生长结果一致。造成这一结果的主要原因可能是高温胁迫抑制了番茄幼苗光合作用,使其呼吸作用增强,叶片气孔阻力增加,光合作用下降,碳水化合物的积累降低。高温胁迫破坏了细胞膜稳定性,从而使番茄幼苗地上部分营养生长受阻。综上所述,由于番茄幼苗在高温胁迫条件下促花激素分泌量减少,使植株生殖发育速率减缓,即使进入生殖生长,萼片、雄蕊和雌蕊的分化和细胞膨大速率也会减缓甚至停滞。
本研究以番茄品种“寿和粉冠”为试材,系统研究了高温高湿环境对番茄花芽分化进程的影响及花芽分化过程中番茄体内营养物质及生长指标的变化,是否适应于其它作物与品种还需进一步研究,高温高湿环境下番茄幼苗花芽分化阶段内源激素分泌的机理以及其它发育时期的生理响应机制亦需进一步研究与探讨。
[1] 李英楠,白亚丽,杜南山,等.有机复混肥替代化肥减施对日光温室番茄生长及土壤环境的影响[J].中国瓜菜,2019, 32(11):43-47. Li Y N,Bai Y L,Du N S,et al.Effects of organic compound fertilizer instead of chemical fertilizer on tomato growth and soil environment in solar greenhouse[J].Chinese Melon and Vegetable,2019,32(11):43-47.(in Chinese)
[2] 项荣,张杰兰.基于改进PCNN的番茄植株夜间图像分割算法[J].农业机械学报,2020,51(3):130-137. Xiang R,Zhang J L.Tomato image night segmentation algorithm based on improved PCNN[J].Journal of Agricultural Machinery,2020,51(3):130-137.(in Chinese)
[3] 蒋程瑶,宋羽,李玉姗.不同叶背补光模式对戈壁温室番茄叶片光合性能与固碳效应的影响[J].中国蔬菜,2019(10): 32-38.Jiang C Y,Song Y,Li Y S.Effects of different back-filling modes on photosynthetic performance and carbon sequestration of tomato leaves in Gobi greenhouse[J]. Chinese Vegetables, 2019(10): 32-38.(in Chinese)
[4] 王丰建.夏季番茄病毒病的发生与防治[N].山东科技报,2019-07-01(005). Wang F J.Occurrence and prevention of tomato virus disease in summer[N].Shandong Science and Technology News,2019-07-01(005).(in Chinese)
[5] 赵伟,白青,张凯,等.减磷施肥对番茄光合特性的影响[J].陕西农业科学,2019,65(6):27-29. Zhao W,Bai Q,Zhang K,et al.Effect of phosphorus reduction and fertilization on tomato photosynthetic characteristics[J]. Shaanxi Agricultural Sciences, 2019,65(6):27-29.(in Chinese)
[6] 游薇,战鸿彬,金朝霞.拮抗菌诱导番茄根系分泌物鉴定及作用研究[J].生物学杂志,2020,37(1):54-58. You W,Zhan H B,Jing Z X.Antagonism and antibacterial induction of tomato root secretions and their effects[J]. Journal of Biology,2020,37(1):54-58.(in Chinese)
[7] 马盼,王学成,朱长华,等.不同温度对番茄育苗效果的影响[J].上海蔬菜,2019(6):44-45. Ma P,Wang X C,Zhu C H,et al.Effects of different temperatures on tomato seedling raising effect[J].Shanghai Vegetables,2019(6):44-45.(in Chinese)
[8] 程雯,王妍.引发技术对不同温度下番茄种子萌发和幼苗生长的影响[J].粮食科技与经济,2019,44(1):87-89. Cheng W,Wang Y.Effect of priming technology on tomato seed germination and seedling growth at different temperatures[J].Food Science and Technology and Economy,2019,44(1):87-89.(in Chinese)
[9] 张启好.番茄温度胁迫响应中组蛋白去乙酰化酶HDACs的功能研究[D].杭州:浙江大学,2019. Zhang Q H.Functional study of histone deacetylase HDACs in response to tomato temperature stress[D]. Hangzhou: Zhejiang University,2019.(in Chinese)
[10]张淑杰,孙立德,马成芝,等.日光温室番茄低温冻害指标确定及温度预报模型建立[J].气象与环境学报,2016, 32(4): 98-105. Zhang S J,Sun L D,Ma C Z,et al.Determination of low-temperature freezing injury index of tomato and establishment of temperature prediction model in sunlight greenhouse[J].Journal of Meteorology and Environment, 2016,32(4):98-105.(in Chinese)
[11]尹赜鹏,鹿嘉智,魏雪莹,等.适当遮光对高温强光下番茄幼苗叶片光合功能的影响[J].沈阳农业大学学报,2020, 51(1): 43-51. Yin Z P,Lu J Z,Wei X Y,et al.Effect of proper shading on photosynthetic function of tomato seedling leaves under high temperature and strong light[J].Journal of Shenyang Agricultural University,2020,51(1):43-51.(in Chinese)
[12]袁昌洪,杨再强,赵和丽.番茄高温高湿胁迫后的补偿生长[J].生态学杂志,2020,39(2):487-496. Yuan C H,Yang Z Q,Zhao H L.Compensatory growth of tomato after high temperature and high humidity stress[J].Journal of Ecology,2020,39(2):487-496.(in Chinese)
[13]李胜利,夏亚真,孙治强.冷激对高温胁迫下番茄幼苗生长及花芽分化的影响[J].应用生态学报,2016,27(2):477-483. Li S L,Xia Y Z,Sun Z Q.Effects of cold shock on tomato seedling growth and flower bud differentiation under high temperature stress[J].Chinese Journal of Applied Ecology, 2016,27(2):477-483.(in Chinese)
[14]李胜利,毕明明,陈菲,等.冷激诱导高温胁迫下番茄幼苗矮化机理[J].应用生态学报,2015,26(7):2063-2068. Li S L,Bi M M,Chen F,et al.Mechanism of dwarfing tomato seedlings under high temperature stress induced by cold shock[J].Chinese Journal of Applied Ecology,2015, 26(7): 2063-2068.(in Chinese)
[15]Ahamed M T,Suvo TP,Chakrobarti M,et al.Effects of different media based hydroponics on yield and nutrient of tomato[J].Bangladesh J,2016,30:51-54.
[16]Bagamboula C F,Uyttendaele M,Debevere J,et al.Inhibitory effect of thyme and basil essential oils,carvacrol, thymol, estragol,linalool and p-cymene towardsand[J].Food Microbiol,2004,21:33-42.
[17]Penso G A,Citadin I,Scariotto S,et al.Development of peach flower buds under low winter chilling conditions[J]. MDPI, 2020,10(3):1003-1032.
[18]Pornpairin R,Yukihiro F,Satoshi T,et al.Low temperature requirements for flower bud differentiation of brussels sprouts(var.Zenk.)[J]. Japanese Society of Agricultural Technology Management, 2019,15(1):757-778.
[19]Hidaka K,Dan K,Imamura H,et al.Crown-cooling treatment induces earlier flower bud differentiation of strawberry under high air temperatures[J]. Environmental Control in Biology,2017,55(1):261-283.
[20]杨丽,杨际双.‘神马’菊花花芽分化与内源多胺的关系[J].西北植物学报,2008(10):2067-2072. Yang L,Yang J S.Relationship between flower bud differentiation and endogenous polyamines in 'Shenma' chrysanthemum [J].Acta botany Sinica, 2008(10): 2067- 2072. (in Chinese)
[21]袁慧敏,王革伏,樊佳茹等.高温对番茄幼苗生长和花芽分化的影响[J].西北植物学报,2019,39(10):1768-1775. Yuan H M,Wang G F,Fan J R,et al.Effects of high temperature on tomato seedling growth and flower bud differentiation[J].Journal of Northwestern Plants,2019, 39(10):1768-1775.(in Chinese)
[22]赵玉芬,储博彦,曾春凤,等.GA3对盆栽八仙花促成栽培生长的影响研究[J].北方园艺,2007(1):109-110. Zhao Y F,Chu B Y,Zeng C F,et al.Study on the effect of GA3on the growth of potted hydrangea flowers[J].Northern Horticulture,2007(1):109-110.(in Chinese)
[23]韦婷婷,杨再强,王明田,等.高温与空气湿度交互对花期番茄植株水分生理的影响[J].中国农业气象,2019, 40(5): 317-326. Wei T T,Yang Z Q,Wang M T,et al.The effect of the interaction of high temperature and air humidity on the moisture physiology of tomato plants during flowering[J]. Chinese Agricultural Meteorology, 2019, 40(5): 317-326.
[24]杨再强,朱静,张波,等.高温处理对结果期草莓叶片衰老特征的影响[J].中国农业气象,2012,33(4):512-518. Yang Z Q,Zhu J,Zhang B,et al.The effect of high temperature treatment on the senescence characteristics of strawberry leaves in fruiting stage[J]. Chinese Agricultural Meteorology, 2012, 33(4): 512-518.
[25]王丽娟,孙嘉星,韩卫华,等.水肥减量对土壤硝态氮和番茄产量品质的影响[J].灌溉排水学报,2020,39(3):1-7. Wang L J,Sun J X,Han W H,et al.Effect of water and fertilizer reduction on soil nitrate nitrogen and tomato yield and quality[J].Journal of Irrigation and Drainage, 2020, 39(3):1-7.(in Chinese)
[26]程智慧.中国蔬菜栽培学[J].园艺学报,2015,42(5):978.Cheng Z H. Chinese vegetable culture[J].Journal of Horticulture, 2015, 42(5):978.(in Chinese)
[27]韦婷婷,杨再强,王琳,等.玻璃温室和塑料大棚内逐时气温模拟模型[J].中国农业气象,2018,39(10):644-655.Wei T T,Yang Z Q,Wang L,et al.Hourly temperature simulation model in glass greenhouses and plastic greenhouses[J]. Chinese Journal of Agometeorology, 2018, 39(10):644-655.
[28]韩佩汝,张正伟,郑静,等.低温对草莓花芽分化的影响[J].中国农业大学学报,2019,24(1):30-39. Han P R,Zhang Z W,Zheng J,et al.Effect of low temperature on strawberry flower bud differentiation[J].Journal of China Agricultural University,2019,24(1):30-39.(in Chinese)
[29]杨盛,白牡丹,郭黄萍.环境因子与花芽分化关系研究进展[J].内蒙古农业大学学报(自然科学版),2018,39(5):97-100. Yang S,Bai M D,Guo H P.Research progress on the relationship between environmental factors and flower bud differentiation[J].Journal of Inner Mongolia Agricultural University(Natural Science Edition),2018,39(5):97-100.(in Chinese)
[30]王海波,赵君全,王孝娣,等.新梢内源激素变化对设施葡萄花芽孕育的影响[J].中国农业科学,2014, 47(23): 4695- 4705. Wang H B,Zhao J Q,Wang X D,et al.The effect of endogenous hormone changes of new shoots on the inoculation of grape buds in facilities[J].Scientia Agricultura Sinica, 2014,47(23):4695-4705.(in Chinese)
[31]冯枫,杨际双.切花秋菊‘神马’花芽分化与内源激素的关系[J].中国农业科学,2011,44(3):552-561. Feng F,Yang J S.The relationship between the flower bud differentiation of cut flower autumn chrysanthemum and the endogenous hormone[J].Chinese Journal of Agricultural Sciences,2011,44(3):552-561.(in Chinese)
[32]聂梅.温度对地被菊‘紫娇’生长发育及花芽分化的影响研究[D].太古:山西农业大学,2019. Nie M.The effect of temperature on the growth and development and flower bud differentiation of ground cover chrysanthemum ‘Zijiao’ [D].Taigu:Shanxi Agricultural University,2019.(in Chinese)
[33]郭东强,卢陆峰,卢晨升,等.邓恩桉开花结实期间内源激素含量变化[J].桉树科技,2019,36(4):22-26. Guo D Q.Lu L F,Lu C S,et al.Deng Enchanges in endogenous hormone content during flowering and fruiting period[J]. Eucalyptus Science and Technology, 2019,36(4):22-26.(in Chinese)
[34]邹玉玲,钟秋平,葛晓宁,等.复硝酚钠对油茶花芽内源激素和花芽分化率的影响[J].经济林研究,2020,38(2):69-76. Zou Y L,Zhong Q P,Ge X N,et al.The effect of sodium nitrophenolate on the endogenous hormones and the differentiation rate of flower buds of[J]. Economic Forest Research,2020,38(2):69-76.(in Chinese)
[35]王磊,汤庚国,刘彤.石蒜花芽分化期内源激素和核酸含量的变化[J].南京林业大学学报(自然科学版),2008(4): 67-70. Wang L,Tang G G,Liu T.Changes in endogenous hormones and nucleic acid content during flower bud differentiation of[J].Journal of Nanjing Forestry University (Natural Science Edition),2008(4):67-70.(in Chinese)
[36]李广,王建红,车少臣,等.侧柏花芽分化期间内源激素变化规律[J].北方园艺,2019(22):90-96. Li G,Wang J H,Che S C,et al.Changes of endogenous hormones during flower bud differentiation of[J].Northern Horticulture,2019(22):90-96.(in Chinese)
[37]赵通,陈翠莲,程丽,等.‘李光杏’花芽分化时期内源激素及碳氮比值的动态研究[J].干旱地区农业研究,2020,38(3): 97-104. Zhao T,Chen C L,Cheng L,et al.Research on the dynamics of endogenous hormones and carbon-nitrogen ratio during the flower bud differentiation of 'Liguang Apricot'[J]. Agricultural Research in the Arid Areas,2020,38(3): 97-104.(in Chinese)
[38]艾星梅,何睿宇,胡燕芳.马铃薯花芽分化与内源激素动态变化的关系[J].西北植物学报,2018,38(1):87-94. Ai X M,He R Y,Hu Y F.The relationship between potato flower bud differentiation and dynamic changes of endogenous hormones [J].Journal of Northwestern Plants, 2018,38(1): 87-94.(in Chinese)
[39]张海娜,鲁向晖,金志农,等.高温条件下稀土尾砂干旱对4种植物生理特性的影响[J].生态学报,2019,39(7): 2426- 2434. Zhang H N,Lu X H,Jin Z N,et al.Effect of drought of rare earth tailings on physiological characteristics of four plants under high temperature conditions[J].Acta Ecologica Sinica,2019,39(7):2426-2434.(in Chinese)
[40]Silvia L,Natasa K,Andrea P,et al.The total path length hydraulic resistance according to known anatomical patterns: what is the shape of the root-to-leaf tension gradient along the plant longitudinal axis[J].Elsevier Ltd, 2020,502: 2208-2224.
[41]Semenov M V,Krasnov G S,Semenov V M,et al.Long-term fertilization rather than plant species shapes rhizosphere and bulk soil prokaryotic communities in agroecosystems[J]. Elsevier B.V.,2020,154:757-779.
[42] Figueroa L L,Grab H,Wee H N,et al.Landscape simplification shapes pathogen prevalence in plant- pollinator networks[J]. Ecology Letters,2020,23(8): 326-354.
Discussion on the Mechanism of Effects of High Temperature and Humidity on Tomato Flower Bud Differentiation in Seedling Stage
HUANG Qin-qin1, YANG Zai-qiang1,2,LIU Xian-nan1, WANG Xue-lin3,XU Chao1, DING Yu-hui1, LI Jia-jia1, ZHENG Qian-tong1
(1. Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters, Nanjing University of Information Science & Technology, Nanjing 210044, China;2. Jiangsu Provincial Key Laboratory of Agrometeorology, Nanjing 210044; 3.Hefei MeteorologicalBureau, Hefei 230041)
In order to study the mechanism of high temperature and humidity affecting the differentiation of tomato flower buds, the tomato variety "Shouhe Fenguan" was used as the test material. The orthogonal test of air temperature, air relative humidity and treatment days was conducted in the agricultural meteorological experimental station of Nanjing University of information technology from April to July 2020. The air temperature (day temperature / night temperature) was set with four treatment levels: T1 (32℃/22℃), T2 (35℃/25℃), T3 (38℃/28℃), T4 (41℃/31℃); The air humidity was set at three levels: H1 (50%), H2(70%) and H3(90%), error range is ±5 percentage points. The treatment time was 2,4,6 and 8 days. The treatments of day / night temperature 28℃/18℃ and air relative humidity 45% − 55% were used as control (CK). The contents of endogenous hormones, starch and soluble sugar were measured at different stages of tomato flower bud differentiation, and stem diameter, dry weight of single plant, strong seedling index and chlorophyll content were measured at budding stage to study the mechanism of high temperature and high humidity on flower bud differentiation of tomato. The results showed that: (1) with the increase of temperature, the whole process of flower bud differentiation was prolonged with the increase of temperature, while the air relative humidity and treatment days had little effect on the process of tomato flower bud differentiation. (2) Under different treatments, the contents of IAA and GA3in the top buds of tomato decreased, increased and decreased with the flower bud differentiation, while the contents of ZT and ABA showed the opposite trend with IAA. The contents of IAA, ZT and GA3decreased with the increase of temperature, relative humidity and treatment days, while ABA content increased with the increase of stress degree. (3) The content of starch and chlorophyll in tomato leaves decreased gradually with the process of flower bud differentiation. Soluble sugar content increased gradually from non differentiation stage to stamen differentiation stage, and decreased gradually during pistil differentiation stage. With the deepening of stress degree, there were significant differences among the treatments. The results showed that the inhibitory effect of high temperature and high humidity on tomato flower bud differentiation might be related to the change of endogenous hormone content and the decrease of nutrients. The environmental temperature should be controlled at the level of CK at the initial stage of flower bud differentiation. The higher the temperature, the more unfavorable it would be. The results can provide some scientific basis for tomato growth environment regulation and disaster warning.
Tomato; High temperature; High humidity; Endogenous hormones; Flower bud differentiation process; Nutrients
10.3969/j.issn.1000-6362.2021.01.006
黄琴琴,杨再强,刘显男,等.苗期高温高湿影响番茄花芽分化进程的机理探讨[J].中国农业气象,2021,42(1):56-68
2020−07−19
国家自然科学基金面上项目“高温高湿对设施番茄果实品质影响机理及风险评估模型的研究”(41775104);“高温胁迫下氮素对设施番茄果实糖酸代谢影响机理与品质调控的研究”(41975142);科技部重点研发计划“主要经济作物气象灾害风险预警及防灾减灾关键技术”(2019YFD100021)
杨再强,教授,研究方向为设施农业气象、特色农业气象、农业生物环境调控,E-mail:yzq@nuist.edu.cn
黄琴琴,E-mail:1510141893@qq.com
