基于林分生长量的沙地樟子松初植造林密度确定
2021-01-19袁春良张学利安宇宁孙晓辉
韩 辉,袁春良,张学利,宋 鸽,安宇宁,孙晓辉
(1.辽宁省沙地治理与利用研究所,辽宁 阜新 123000;2.辽宁章古台科尔沁沙地生态系统国家定位观测研究站,辽宁 阜新 123000)
樟子松Pinussylvestrisvar.mongolica主要天然分布在我国大兴安岭呼伦贝尔沙地,具有喜光、抗旱、耐贫瘠、耐寒、速生和适于沙地等优良特性,自20世纪50年代引种到科尔沁沙地南缘营建防风固沙林以来,发挥了巨大的防护功能[1-2],已经成为三北防护林工程中最重要的针叶造林树种之一。章古台引种樟子松以来,研究人员开展了樟子松经营密度[3-5]、生长进程[6-7]、生长量[8-12]、自然稀疏[12]、抚育间伐[13-14]、樟子松衰退原因[1,15-16]、造林密度[17-18]、立地影响[19-21]等诸多方面的研究。20世纪90年代以来,引种地章古台的中成熟林樟子松出现生长衰弱死亡以来,林分密度过大被认为是导致林分衰退或死亡的最直接的原因之一[1,16]。樟子松引种到章古台后生长进程加快,生命周期缩短,成熟期提前[7],探讨适宜的造林密度具有实践意义。樟子松的造林密度直接影响林分的生长[22-24],对章古台不同造林密度、不同林龄樟子松生长量的研究多有报道[18,25],但是对于不同造林密度及丛植模式条件下樟子松生长40年后的林分状况少有报道,本文拟通过对不同造林密度及丛植模式下樟子松成熟林林分胸径、树高、冠幅生长量的调查研究,提出适宜的造林密度及造林模式,为樟子松在三北地区的造林及经营提供参考依据。
1 材料与方法
1.1 研究区概况
研究区选择在科尔沁沙地南缘的辽宁省沙地治理与利用研究所章古台实验林场三家子实验区,地理位置:42°41′ N,122°33′ E,海拔204.5 m,年均气温6.3 ℃,全年无霜期150~160 d,年均降水量475.5 mm,年均蒸发量1 553.2 mm。土壤以风沙土为主,根据当地钻探资料,沙层厚度可达126~128 m[26],沙土瘠薄,樟子松林0~30 cm土层内有机质含量为4.2~5.0 g·kg-1[27]。主要代表性植物有色木槭Acermono、山里红Crataeguspinnatifida、榆树Ulmuspumila、山杏Prunussibirica、樟子松、彰武小钻杨Populus×xiaozhuanica‘zhangwu’、胡枝子Lespedezabicolor、中华隐子草Cleistogeneschinensis、拂子茅Calamagrostisepigeios、蒺藜梗Agriophyllumsquarrosum等。
1.2 试验设计
1980年用2年生樟子松裸根苗造林,造林设计采取不同株行距或者不同丛植模式(表1),每个小区面积大小为80 m×80 m,各小区标准地林分调查实际面积约为670 m2,共计27个小区。各小区实际调查面积之外的地块在1992年、1997年、2002年分别开展了透光抚育及抚育间伐。现存各小区的密度形成时间为近5 a。
1.3 数据分析方法
采用Excel 2007软件作图,SPSS 17.0软件进行差异性分析。采用SPSS 17.0软件进行单因素方差分析(ANOVA)得到不同初植密度试验林林分因子的差异性,运用最小显著差异法(LSD)进行多重比较分析。
2 结果与分析
2.1 不同初植密度林分因子生长量的差异性
不同小区间胸径、全树高、活枝下高、冠幅总体均值都存在显著性差异(表2)。
2.1.1 樟子松不同初植密度林分40年后现存林木胸径生长量的差异
随着密度的增大,各小区平均胸径呈现减小趋势,胸径最大值在下移,最小值也在下移,但在密度930株·hm-2(21小区)之后,胸径较小值稳定在一定范围,并没有随着密度的增大而继续减小(图1)。调查发现很多小区内存在个别林木胸径较大,作为异常值而存在,可见在密度较大林分中林木分化虽然更趋强烈,但依然有较大胸径的林木存在。随着密度的增大,各小区内胸径数值变化的差异也呈现出缩小的趋势。8小区(963 株·hm-2)林分胸径相对较大,该小区初植株行距为3 m×4 m,按1穴2株和1穴3株定植,该地立地条件处于坡底,地势较低,相当于洼坑里。

表1 樟子松不同初植密度试验林分标准地基本情况

表2 樟子松不同初植密度林分40年后各项因子调查结果
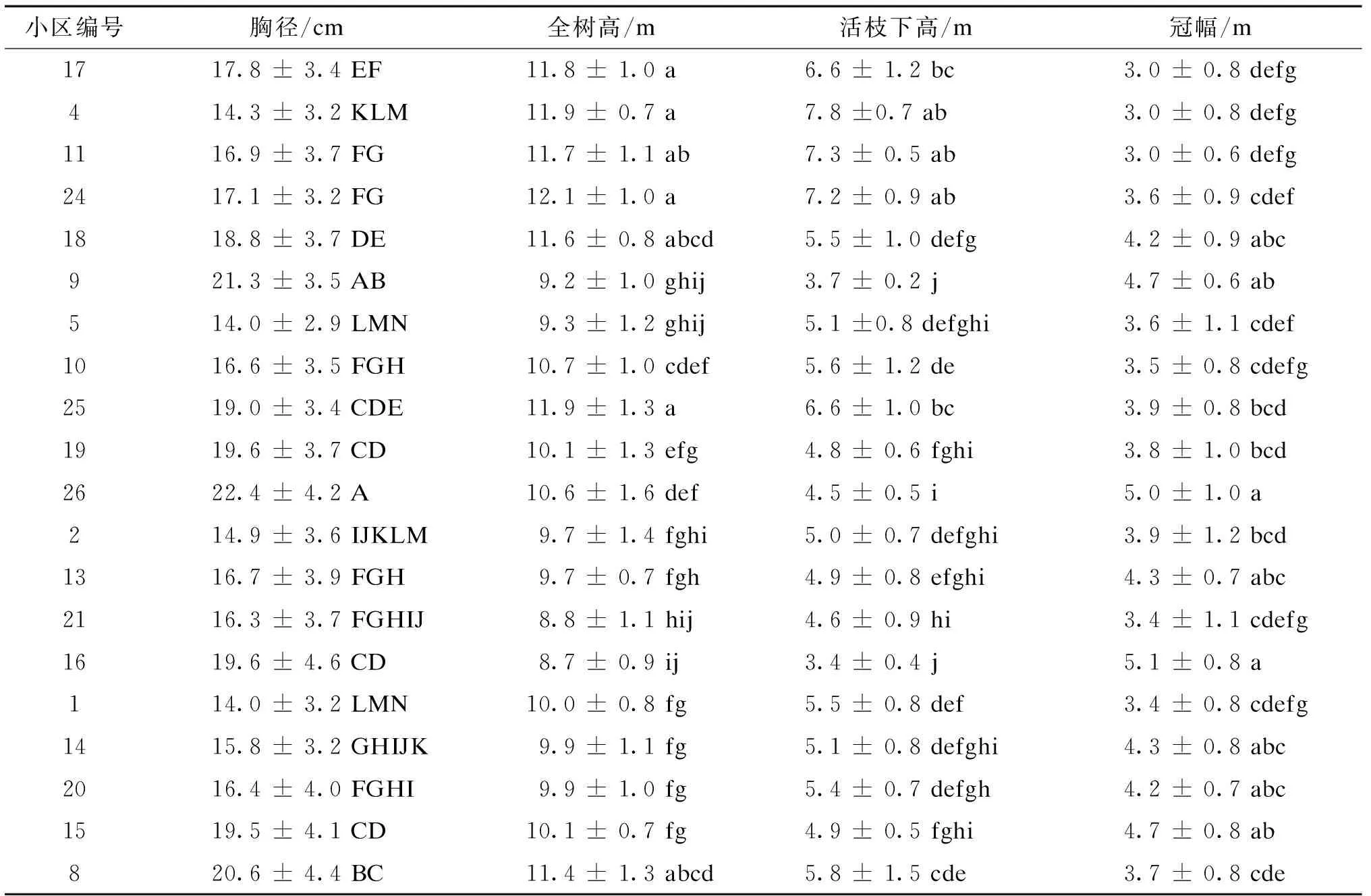
续表2 樟子松不同初植密度林分40年后各项因子调查结果

注:图中小区编号前后排序按照现存密度从大到小排序,下图同。图1 樟子松不同初植密度林分40 年后胸径生长量的差异
2.1.2 樟子松不同初植密度林分40年后全树高生长量的差异
随着密度的增大,全树高刚开始呈现增大走势,但上升到一定程度之后就不再随着密度的增大而增大,而是维持在一定水平,偶有上升或下降。树高(H)与现存密度(D现)的关系式为:H=0.000 5×D现+9.853 7(R2=0.05)。各小区之间全树高的差异性显著。在密度390~1 014 株·hm-2(9~25小区)范围内全树高随着密度的上升而上升,之后稳定在一定范围,偶尔有下降。调查发现,小区内全树高数值变化较大的有26、25、24小区,对应株行距分别为3 m×4 m、1.5 m×4 m、1.5 m×3 m;全树高数值变化较小的有22、4、12、3小区,对应株行距分别为1 m×1 m、0.5 m×3 m、1 m×2 m、0.5 m×2 m,即全树高数值变化大的株行距比较大,数值变化小的株行距比较小。密度较大而全树高又相对较小的有5、23、27、6小区,这几个小区的立地条件多为坡顶或丘顶(图2)。

图2 樟子松不同初植密度林分40年后全树高、冠幅生长量的差异
2.1.3 樟子松不同初植密度林分40年后冠幅生长量的差异
冠幅随着密度的增大呈现下降趋势,这与密度大妨碍树冠的生长有直接关系,密度增大到现存密度1 528 株·hm-2(23区)以上时,冠幅不再下降,而是趋于稳定在一定的低水平范围之内(图2)。
2.2 各因子间相关性分析
为了保证试验的科学性和连续性,各小区现存密度的形成完全是林分自然稀疏即自然选择的结果,不存在人为干扰因素,只是各小区有的树木死亡之后不定期地被人为清除。胸径随着冠幅的增大呈现增大趋势,呈现线性关系,胸径(DBH)与冠幅(W)的关系式为:DBH=3.336 5W+4.685 9(R2=0.71)(图3)。各小区按2020年6月调查时林分的平均胸径和保存率数据进行分析,胸径与现存密度(D现)的关系式为:DBH=-0.004 4D现+22.423(R2=0.75),胸径随着现保存密度的增大呈现减小趋势(图4)。各小区保存率(Rr)与初植密度(D初)的关系式为:Rr=-0.003 6D初+54.758(R2=0.61),保存率随着初植密度的增大呈现下降趋势,初植密度在3 000株·hm-2以上时保存率呈现大幅度下降趋势,在7 000~10 000株·hm-2时呈现出稳定的较低保存率。相同密度而保存率有高有低,这可能与立地条件有很大关系(图5)。现存密度与初植密度的关系式为:D现=705.651×ln(D初)-4 360.7(R2=0.81),现存密度随着初植密度的增大而增大,即初植密度小时,现存密度也小,初植密度大时,现存密度也大,但在初植密度7 000~10 000 株·hm-2时,现存密度趋于平缓和稳定(图6)。

图3 樟子松胸径与冠幅的关系

图4 樟子松胸径与现存密度的关系

图5 樟子松保存率与初植密度的关系

图6 樟子松现存密度与初植密度的关系
2.3 樟子松不同株行距对胸径生长量的影响
2.3.1 樟子松相同行距不同株距造林模式对胸径生长量的影响
相同行距时,行距1~4 m的造林模式,随着株距的增加,胸径都呈现出上升趋势(图7)。相同行距模式下不同株距之间,各小区间差异性显著(表2)。行距1 m时,22区(1 m×1 m)胸径高于27区(1 m×1 m)和6区(1.5 m×1 m),现场调查发现应该是因为22区处于坡底,是洼地,而27区和6区都位于坡顶,坡底与坡顶海拔相差近5~10 m。行距2 m时,12区(1 m×2 m)与23区(1.5 m×2 m)胸径差异性不显著。行距3 m时,11区(1 m×3 m)与24区(1.5 m×3 m)胸径差异性不显著。行距4 m时, 25区(1.5 m×4 m)与19区(2 m×4 m)胸径差异性不显著(表2)。
2.3.2 樟子松相同株距不同行距模式对胸径生长量的影响
相同株距时,株距0.5 ~3 m的造林模式,随着行距的增加,胸径都呈现出上升趋势(图8)。

图7 樟子松相同行距不同株距造林模式胸径平均生长量差异

图8 樟子松相同株距不同行距造林模式胸径平均生长量差异
相同株距模式下不同行距之间,各小区间差异性除株距2 m模式外都呈显著性(表2)。株距0.5 m时,3区(0.5 m×2 m)与4区(0.5 m×3 m)、5区(0.5 m×4 m)差异性显著,但4区和5区之间差异性不显著。株距1 m时,27区(1 m×1 m)胸径与其他小区差异性显著,22区(1 m×1 m)与12区(1 m×2 m)差异性不显著,11区(1 m×3 m)与10区(1 m×4 m)之间差异性不显著,22区、12区与11区、10区之间差异性显著。株距1.5 m时,各小区之间差异性都显著。株距2 m时,17区(2 m×2 m)与18区(2 m×3 m)、19区(2 m×4 m)差异性显著,18区与19区之间差异性不显著。株距3 m时,7区(3 m×1 m)与9区(3 m×3 m)、26区(3 m×4 m)差异性显著,9区和26区之间差异性不显著(表2)。
2.4 樟子松不同株行距对全树高生长量的影响
2.4.1 樟子松相同行距不同株距造林模式对全树高生长量的影响
相同行距时,行距1~4 m的造林模式,全树高没有呈现出完全一致的规律性(图9)。相同行距模式下不同株距之间,各小区间差异性显著(表2)。行距1 m时,22区(1 m×1 m)与7区(3 m×1 m)的全树高较高,两者之间差异性不显著,都与27区(1 m×1 m)和6区(1.5 m×1 m)差异性显著,27区全树高最低,与其它小区差异性都显著。27区位于坡顶,前面已说明。行距2 m时,23区(1.5 m×2 m)树高最低,与其它小区树高差异性都显著,3区(0.5 m×2 m)和17区(2 m×2 m)的树高较高,两者之间差异性不显著,12区(1 m×2 m)与3区之间差异性不显著,但都与23区和17区差异性显著。行距3 m时,9区(3 m×3 m)树高最低,与其它4个小区差异性显著,而其它4个小区之间差异性不显著。行距4 m时,25区(1.5 m×4 m)全树高最高,与其它4个小区差异性显著,5区(0.5 m×4 m)全树高最低,与19区(2 m×4 m)树高差异性不显著,与其它小区差异性显著,10区(1 m×4 m)、19区、26区(3 m×4 m)之间差异性不显著(表2)。

图9 樟子松相同行距不同株距造林模式全树高生长量差异
2.4.2 樟子松相同株距不同行距模式对全树高生长量的影响
相同株距时,株距0.5~3 m的造林模式,随着行距的增加,全树高没有呈现出一致的规律性,有的上升,有的下降(图10)。相同株距模式下不同行距之间,各小区间差异性显著(表2)。株距0.5 m时,5区(0.5 m×4 m)最低,与3区(0.5 m×2 m)和4区(0.5 m×3 m)差异性显著,但3区和4区之间差异性不显著。株距1 m时,27区(1 m×1 m)树高最低,与其他小区差异性显著,22区(1 m×1 m)与11区(1 m×3 m)较高,两小区之间差异性不显著,但与其它小区之间差异性显著,12区(1 m×2 m)与10区(1 m×4 m)之间差异性不显著,但与其它小区之间差异性显著。株距1.5 m时,6区和23区树高较低,两小区之间差异性不显著,但与其它小区之间差异性显著,24区(1.5 m×3 m)与25区(1.5 m×4 m)树高较高,两小区之间差异性不显著。株距2 m时,19区(2 m×2 m)全树高最低,与17区(2 m×2 m)、18区(2 m×3 m)差异性显著,17区与18区之间差异性不显著。株距3 m时,9区(3 m×3 m)树高最低,与7区(3 m×1 m)和26区(3 m×4 m)差异性显著,7区和26区之间差异性不显著(表2)。
2.5 不同丛植模式对胸径生长量的影响
2.5.1 1穴2株丛植模式对胸径生长量的影响
1穴2株丛植模式,随着株行距从2.5 m × 2.5 m到4 m × 4 m,胸径呈上升走势(图11)。不同小区间存在显著性差异。16区胸径最大,与其它区存在显著性差异。13区和21区株行距相同,两者间胸径无显著性差异。2区、13区、21区、16区保存密度分别为1 293、994、930、581 株·hm-2,随着现保存密度的降低,胸径呈上升趋势。小区间随着初植密度的降低,保存率在升高(表2)。
2.5.2 1穴3株丛植模式对胸径生长量的影响
1穴3株丛植模式,随着株行距从2.5 m × 2.5 m到4 m × 4 m,胸径呈上升走势(图11)。

图10 樟子松相同株距不同行距造林模式全树高生长量差异

图11 樟子松不同丛植模式对胸径生长量的影响
不同小区间存在显著性差异。15区胸径最大,与其它区存在显著性差异。14区和20区株行距相同,两者间胸径无显著性差异。1区、14区、20区、15区保存密度分别为1 528、1 457、1 105、705 株·hm-2,随着现保存密度的降低,胸径呈上升趋势。小区间随着初植密度的降低,保存率在升高(表2)。
统计丛植模式的胸径(DBH丛)与密度(D现)的关系式为DBH丛=-0.005 4×D现+ 22.471(R2=0.85)。
2.6 樟子松胸径断面积与密度之间的关系
胸径断面积随着现存密度的增大而增加,呈对数形式,在密度1 500 株·hm-2以上时,趋于平缓,可以断定胸径断面积可能不再随着密度的增大而增加,应当存在一个相对的最大值,而后稳定(图12)。
在初植密度800~2 200 株·hm-2这一区间内,胸径断面积随着初植密度的增大而快速升高,其后不再随着密度的增大而增大,而是趋于稳定在一定的数值范围内(图13)。
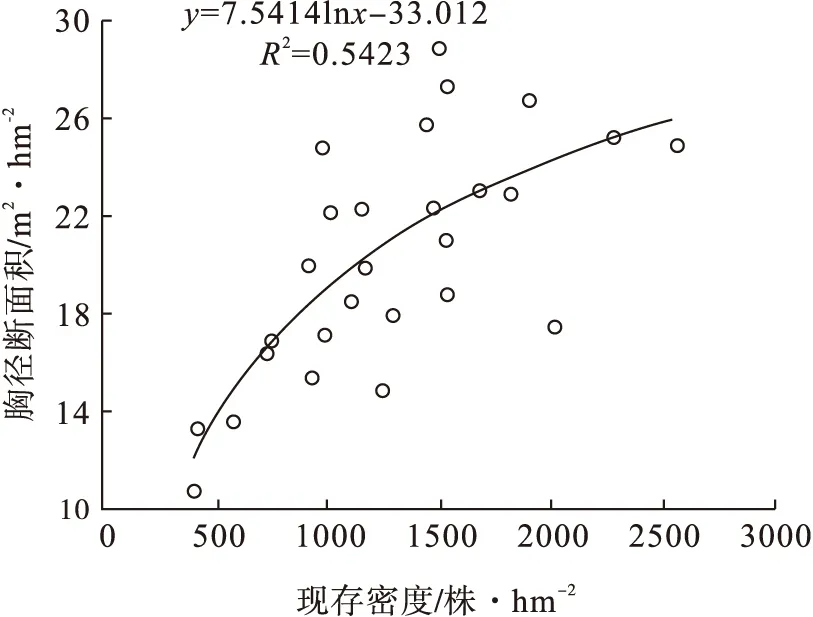
图12 樟子松胸径断面积与现存密度的关系

图13 樟子松胸径断面积与初植密度的关系
3 讨论与结论
3.1 不同初植密度沙地樟子松成熟龄人工林生长量及波动范围
1980年采用樟子松2年生裸根苗造林,到2020年,林龄达到42 a,处于成熟龄阶段。27块不同初植密度10 000~833 株·hm-2演变到现在的密度2 542~390 株·hm-2,平均胸径变动幅度在11.8~22.4 cm,胸径最大值31.3 cm,位于17区,初植株行距2 m×2 m;最小值6.6 cm,位于27区,初植株行距1 m×1 m。平均全树高变动幅度在8.5 ~12.1 m,全树高最大值13.7 m,位于26区,初植株行距3 m×4 m;最小值6.5 m,位于27区和5区,初植株行距分别为1 m×1 m和0.5 m×4 m。平均冠幅变动幅度在2.5~5.1 m;最大冠幅6.15 m,位于26区,初植株行距3 m×4 m;最小冠幅1.05 m,位于4区,初植株行距0.5 m×3 m,这与以往在章古台地区调查的樟子松同龄阶的生长量研究结果相一致[7,25,28]。不同初植密度林分的胸径、树高、冠幅生长量在造林40年后存在显著性差异。樟子松为强阳性树种,树木分化比较强烈,即使在密度较大林分内也有个别胸径较大的林木存在,这为樟子松培育大径材提供重要依据。
本研究樟子松胸径生长量随着密度的增大而减小的结论与相关研究结果一致[29-30],树高没有表现出随着密度的增大而增高,而是呈现出较稳定的结果,可能有立地条件的影响[19,21],也可能有病虫害的影响[31]。
3.2 丛植模式对沙地樟子松成熟龄人工林的影响
不论是1穴2株还是1穴3株,随着株行距从2.5 m × 2.5 m到4 m × 4 m,胸径生长量都呈上升趋势。随着初植密度的升高,保存率在降低。调查发现,虽然造林时为丛植,经过40年生长,70%~80%植穴坑里只保留1株,具体是在什么时候保留1株已经无从考证,但是从较高的保存率以及较大的平均胸径角度考虑,以4 m × 4 m丛植造林模式为好,这为沙地樟子松的初植造林密度和大径材培育提供了良好的借鉴经验。
3.3 沙地樟子松人工林初植造林密度确定的探讨
综合考虑沙地樟子松成熟龄生长量各项指标,在章古台地区的初植造林密度以833株·hm-2为宜,株行距以3 m×4 m为宜,这与毛乌素沙地较好的初植密度以833株·hm-2为好相一致[32-33]。有学者研究认为毛乌素沙地幼龄时较好的株行距为2 m×2 m,中龄时较好的株行距为3 m×4 m[32-33],章古台较好的初植密度为1 667~2 550 株·hm-2,株行距为2 m×3 m[18],结论的差别应该与沙地地理位置不同、调查时樟子松林龄不同有一定关联,但在一定时期都具有很好的借鉴意义。为了培育大径材可以考虑株行距为4 m × 4 m,因为现在造林苗木基本都为容器杯苗,不考虑1穴多株的丛株模式。
1980-2020年各试验地经历了干旱年份,没有一块试验地的林木全部死亡,说明樟子松具有很强的耐旱性和抗旱性,本身具有强烈的分化因素,这对于在适宜地区造林具有重要意义。相反,在生产经营时要特别注意控制病虫害的发生。
不同初植密度樟子松人工林在生长40年后,胸径、全树高、活枝下高、冠幅总体均值都存在显著性差异。现存密度和初植密度之间存在对数关系,胸径和保存率随现存密度的增大而降低,树高随着现存密度的增大而增高,当密度增大到一定程度之后树高不再增加,而是维持在一定范围之内上下波动。以樟子松成熟龄人工林的胸径生长量为指标,考虑到樟子松更持久的固沙防护周期,较好的初植密度以833 株·hm-2为宜,株行距以3 m×4 m或者更大株行距为宜。樟子松造林不提倡丛植。单位面积的樟子松胸径断面积在密度超过1 500株·hm-2后不会再增加,而是稳定在一定范围之内,这就决定了以减少单位面积的株数来达到稳定。在樟子松林分经营过程中,生长抚育间伐以单株树冠之间互不相搭为标准进行不同时段的作业操作。因为胸径与冠幅之间存在一定的线性关系,通过间伐有利于保留林木的生长,有利于提高冠幅。综合不同密度不同模式下各林分的生长量指标,在同一立地条件下,用克拉夫特林木分级法对沙地樟子松人工林同龄林中的林木进行分级时,可以用胸径生长量大小的指标进行划分。
