甜菜碱脂在14 种海洋微藻中的分布研究
2021-01-15李艳荣牟桐黄莉莉徐继林周成旭严小军
李艳荣,牟桐,黄莉莉,徐继林,周成旭,严小军
( 1. 宁波海洋研究院,浙江 宁波 315832;2. 宁波大学 应用海洋生物技术教育部重点实验室,浙江 宁波 315832)
1 引言
甜菜碱脂是一类不含磷的甘油脂类,由两分子的脂肪酸和一分子的极性头部组成,Dembitsky[1]对甜菜碱脂的生化结构和生物学功能做了详细的报道。海洋微藻中的甜菜碱脂主要包括3 大类:二酰甘油-N-三甲基高丝氨酸(diacylgycerol-N-trimethylhomoserine,DGTS),二酰甘油-羟甲基-N,N,N-三甲基-β-丙氨酸(diacylglyceryl-hydroxymethy-N,N,N- trimethylβ-alanine,DGTA)和二酰甘油-O-羧羟甲基胆碱(diacylglyceryl-carboxyhydroxymethylcholine, DGCC), 是海洋微藻中的三大膜脂之一。甜菜碱脂的组成和代谢对于微藻的化学分类具有重要的意义,甜菜碱脂的脂肪酸组成和完整极性脂分子可以作为营养标记,可进一步确认初级生产者在食物网中的作用[2–3]。Banskota等[4]在微绿球藻(Nannochloropsis granulata)中分离出6 种 DGTS, 即 (2S)-1,2-bis-Oeicosapentaenoylglyceryl-3-O-4′-(N,N,N-trimethyl)-homoserine, (2S)-1-O-eicosapentaenoyl-2-O-arachidonoylglyceryl-3-O-4 ′-(N,N,N-trimethyl)-homoserine, (2S)-1-Oeicosapentaenoyl-2-O-myristoylglyceryl-3-O-4′-(N,N,N-trimethyl) -homoserine, (2S)-1-O-eicosapentaenoyl-2-O-palmitoylglyceryl-3-O-4′-(N,N,N-trimethyl)-homoserine, (2S)-1-O-eicosapentaenoyl-2-O-palmitoleoylglyceryl-3-O-4′-(N,N,N-trimethyl)-homoserine 和(2S)-1-O-eicosapentaenoyl-2-O-linoleoylglyceryl-3-O-4′-(N,N,N-trimethyl)-homoserine。这些DGTS 可以通过下调一氧化氮合成酶的表达来抑制RAW264.7 巨噬细胞中NO 的产生,首次表明DGTS 或许具有抗炎活性。此外,甜菜碱脂与微藻的生长和耐盐性有关,DGTS 的含量测定有助于了解微藻生长或耐盐的机制,进而有益于培养出更多使用性微藻[5–6]。
海洋微藻是海洋中最主要的光合生物,硅藻、绿藻、甲藻和定鞭藻是4 种常见的微藻种类,其季节、种类的变化在海洋生态环境、海区生产力、全球碳循环等方面起着重要作用[7–8]。甜菜碱脂是一类极性脂,是生物膜的重要组成成分,在磷胁迫时,甜菜碱脂可以替代磷脂酰胆碱(PC)维持藻类细胞正常的生命活动[9–10]。对于甜菜碱脂的研究,大多数研究者是将甜菜碱脂经过复杂的衍生化或酸化后用液相色谱、气相色谱或气相色谱−质谱联用的方法测定甜菜碱脂的组成[11–13]。近年来,液相色谱和质谱联用系统为海洋微藻中甜菜碱脂的研究提供了一种新的方法[3,14–15]。本文利用超高效液相色谱−四级杆−飞行时间质谱联用分析系统(UPLC-Q-TOF MS)建立了一种同时检测海洋微藻中3 类甜菜碱脂的分析方法,对4 个微藻门中的14 种微藻的甜菜碱脂分子结构及其相应脂肪酸的组成及分布进行分析。为海洋微藻生态学研究提供理论依据,是进一步阐明脂质代谢必然要做的一项工作,有助于更好地了解甜菜碱脂在人类健康领域的作用;另一方面,对不同种类海洋微藻中甜菜碱脂分布的研究,也可以为微藻的系统发育及化学分类提供新的见解。
2 材料与方法
2.1 实验材料
DGTS 标品1,2-Dipalmitoyl-sn-glycero-3-O-4-(N,N,N-trimethyl) homoserine (16:0-16:0 DGTS) 以及DGTS 的同位素标准品(1,2-di-palmitoyl-sn-glycero-3-O-4'-[N,N,N-trimethyl(D9)-homoserine)均购买自美国Avanti Polar Lipids 公司;所有用于样品提取、液质分析的试剂均为色谱级;纯水来自于Milli-Q 纯水仪;抗氧化剂2,6-二叔丁基对甲酚(BHT,>99.9%),购自美国Sigma公司。
2.2 样品的培养和收集
表1 所示的14 种微藻均由宁波大学浙江省海洋生物重点实验室微藻种质库提供。用于微藻培养的海水均经脱脂棉过滤、煮沸消毒,所有容器均高温灭菌。培养液采用f/2 配方[16],藻种在5 000 mL 锥形瓶中(20±2)℃自然光培养,每天摇动数次,并用颗粒粒度计数分析仪(上海求精生化试剂仪器有限公司)测量藻类密度,14 种微藻均在平台期收获,将收获的藻液在5 000 r/min 的条件下高速离心10 min 获取藻泥,冷冻干燥机(美国Labconco 公司)干燥后备用。样品平行培养3 份。

表1 14 种海洋微藻的种名和种质编号Table 1 The strain number and species name of 14 species of microalgae
2.3 样品制备
分别称取各藻类细胞50 mg 干粉,加入氯仿/甲醇(1∶1,V/V)混合溶液提取总脂,氮气浓缩仪(美国ATR 公司)吹干,甲醇复溶后经孔径为0.22 μm 的超滤膜(美国Mil-lipore 公司)离心过滤,然后进行UPLC-Q-TOF-MS(美国Waters 公司)分析。所有溶剂中均加入50 μg/mL BHT。
2.4 UPLC-Q-TOF 分析
用于样品分析的液相色谱和质谱条件见参考文献[17]。
2.5 甜菜碱脂的半定量分析
甜菜碱脂的半定量:分别在每一种微藻样品中加入5 μg/mL 的1,2-dipalmitoyl-sn-glycero-3-O-4’-[N,N,Ntrimethyl(D9)] homoserine DGTS 同位素内标(由于目前市场上只有DGTS 的同位素内标和标准品,微藻中所有甜菜碱脂的半定量分析参照相应DGTS 内标的峰面积)对样品进行半定量分析。根据流动相比例的改变,获得相应甜菜碱脂的离子化效果,从低碰撞能量下的色谱图中提取样品中每一种甜菜碱脂的离子色谱图;根据不同流动相比例下的甜菜碱脂的离子化效果,获得相应峰面积积分值。根据峰面积与浓度的线性关系,利用同位素标准品的浓度和峰面积半定量样品中每毫克干藻粉中甜菜碱脂含量,取3 个平行结果的均值。
3 结果与讨论
3.1 甜菜碱脂质谱规律及鉴定方法研究
由于目前市场上购买不到DGCC 的相关标准品,文中选用文献已报道过的富含DGCC 的微藻进行DGCC 结构质谱碎裂规律的分析。据文献[15]报道,假微型海链藻中可检测到分子组成为16:0/22:6 DGCC,其母离子质荷比为799.59。利用UPLC-Q-TOF MS 对假微型海链藻中16:0/22:6 DGCC 的分子离子峰进行一级和二级质谱扫描。ESI 源正离子模式的一级质谱图上(图1a),m/z=800.61 代表的是16:0/22:6 DGCC的[M+H]+的母离子峰;在碰撞能量为45 eV 时的二级质谱图中(图1b),m/z=104.11、m/z=132.10 和m/z=178.11的碎片离子的元素组成分别为[C5H14NO]+、[C6H14NO2]+和[C7H16NO4]+,这与Kato 等[18]的研究结果一致,其中m/z=104.11 碎片离子的丰度较高,可以作为DGCC 的特征碎片离子,用于DGCC 的定性。此外,m/z=472.37([M+H-R2COOH]+)和m/z=544.36([M+H-R2COOH]+)的碎片离子峰,可用于DGCC 脂肪酸链的分析。
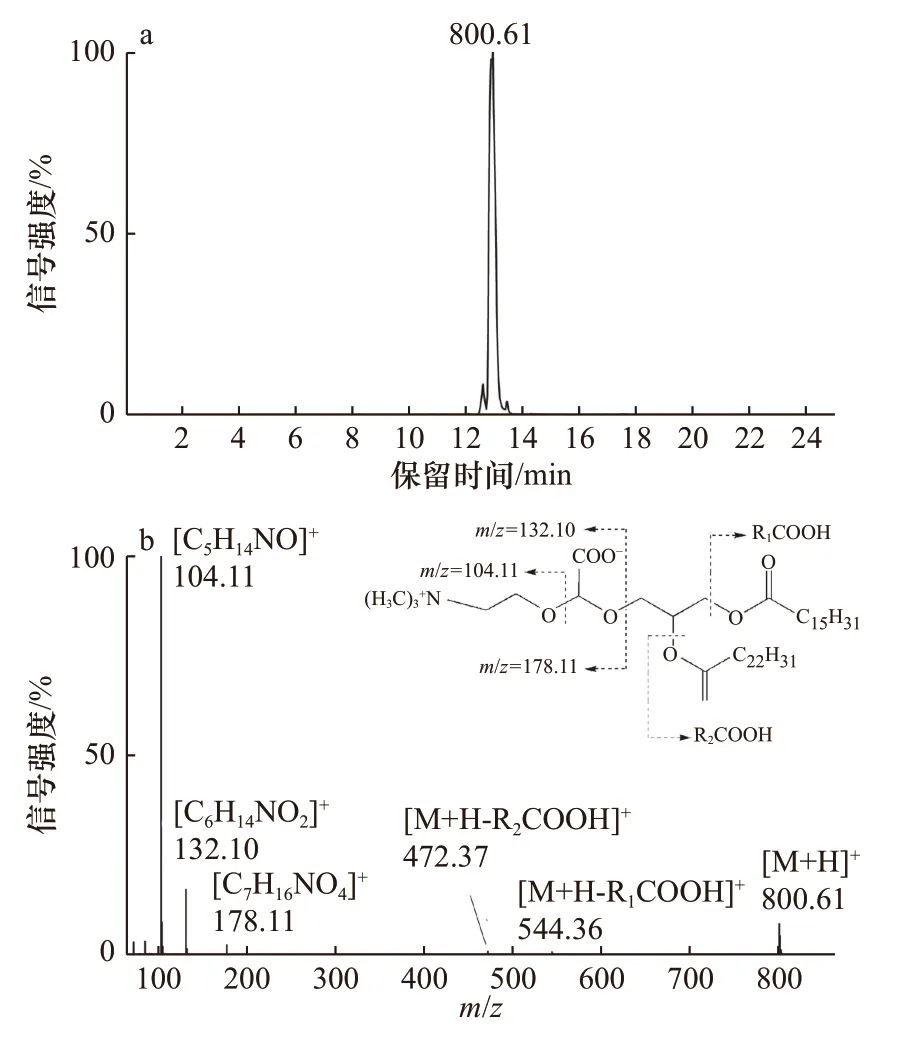
图1 正离子MSE 模式,假微型海链藻中16:0/22:6 DGCC ([M+H]+,m/z=800.61)低能量下提取离子流色谱图 (a);正离子模式下假微型海链藻保留时间为12.98 min16:0/22:6 DGCC [M+H]+(m/z=800.61)二级质谱图(b)Fig. 1 EIC of 16:0/22:6 DGCC ([M+H]+, m/z= 800.61) in T. pseudonana at the low collision energy of 6 V in the positive mode with the MSE technique (a); the MS2 spectrum of the[M+H]+ ion at m/z=800.61 of 16:0/22:6 DGCC at retention time 12.98 min extracted from T. pseudonana (b)
基于已总结的DGCC 的质谱碎裂规律,采用MSE技术对微藻样品中的DGCC 进行分析,可以在一次进样全扫中通过高低能量转换得到样品中每一个物质的母离子和对应子离子精确质量数。在MSE的二级质谱图中,母离子和子离子可以同时存在,从高碰撞能量扫描的总离子流图中提取含有m/z=104.11 的特征碎片离子的色谱图;然后在相同保留时间点,从低碰撞能量扫描的总离子流图中找到相应碎片离子的[M+H]+的母离子;最后针对每一个母离子进行二级质谱分析,进一步确定DGCC 的结构。例如,在颗石藻的高碰撞能量扫描的总离子流图中提取m/z=104.11碎片离子的色谱峰(图2b),从低碰撞能量扫描的总离子流图的对应保留时间找到每一个DGCC 的母离子(图2a),然后针对每一个母离子分别进行二级质谱分析,对每一个DGCC 的分子组成进行鉴定。通过进一步的二级质谱分析,在颗石藻中一共检测到17 种DGCC 甜菜碱脂(表2)。

图2 正离子MSE 模式,颗石藻总脂混合物的低能量总离子流色谱图(a);高能量MSE 色谱图中提取DGCC 特征碎片离子m/z=104.11 的离子流色谱图(b)Fig. 2 TIC of the lipid mixture extracted from P. carterae at the low collision energy of 6 V in the positive mode with the MSE technique (a); EIC of m/z=104.11 for characteristic product ion of DGCC in P. carterae from high collision energy scans (ramp of 30–60 V) in the positive mode with the MSE technique (b)
DGTS 和DGTA 是两种同分异构体,[M+H]+的二级质谱碎裂方式非常相似,均可以产生特征碎片离子m/z=236.15 和m/z=144.10。为了更有效地鉴定和区分这两种甜菜碱脂的分子组成,本文采用[M+H]+和[M+Li]+两种加合形式相结合的方法。在流动相中加入0.01% LiCl 和0.1% HCOOH,可以同时检测到DGTS和DGTA 的[M+H]+和[M+Li]+的母离子峰。首先在[M+H]+的条件下通过特征碎片离子和脂肪酸链的碎片离子鉴定出DGTS 或DGTA 的分子组成,然后在[M+Li]+的二级质谱图中根据碎片离子区分DGTS 和DGTA。关于DGTS 和DGTA 具体的二级碎裂规律及详细鉴定方法参照本实验室已发表文献[17]。
3.2 14 种海洋微藻中甜菜碱脂的组成和分布
根据以上研究的甜菜碱脂的特征碎片离子及其分析鉴定方法,对4 个门的14 种微藻中的甜菜碱脂组成进行分析。共鉴定出133 种甜菜碱脂,包括53 种DGCC、41 种DGTS 和39 种DGTA(表2 至表4)。本研究基于MSE技术建立的系统分析海洋微藻中甜菜碱脂的检测方法灵敏度更高,检测到甜菜碱脂的种类更多。Cañavate 等[15]在假微型海链藻中共检测到8 种DGCC,而本研究在假微型海链藻中共检测出26 种DGCC 分子,其中还包括6 种只含有一条脂肪酸链的甜菜碱脂,该方法鉴定的种类和数量具有明显优势。
Armada 等[3]对两种定鞭金藻Pseudoisochrysis paradoxaVLP(等鞭金藻纲,Isochrysidaceae)和Diacronema vlkianumVLP(巴夫藻纲,Pavlovaceae)中的甜菜碱脂的研究发现,等鞭金藻纲的P. paradoxaVLP可以同时检测到3 种类型的甜菜碱脂,而巴夫藻纲的D. vlkianumVLP 中仅检测到DGCC 和DGTS 两种类型的甜菜碱脂,且14:0/18:1 DGTS 占D. vlkianumVLP中总DGTS 的90.9%以上。本研究的3 种定鞭金藻中,湛江等鞭金藻和球等鞭金藻属于等鞭金藻纲,而颗石藻属于颗石藻纲,在湛江等鞭金藻和球等鞭金藻中除了DGCC 外均检测到14:0/18:1 DGTS,这与Armada 等[3]的研究结果一致,而颗石藻种检测到的DGTS 脂肪酸链组成为16:0/22:6。研究结果表明,等鞭金藻纲主要的甜菜碱脂为DGCC,且仅含有或主要含有14:0/18:1 DGTS,进一步证明14:0/18:1 DGTS 及其含量占比可以作为等鞭金藻纲的特征甜菜碱脂,为等鞭金藻门等鞭金藻纲藻的系统分类提供一种新的依据。
假微型海链藻中发现26 种DGCC,威氏海链藻中发现25 种DGCC 和1 种DGTS,角毛藻中发现16 种DGCC;在小新月菱形藻和三角褐指藻中没有发现DGCC,但分别检测到27 种和30 种DGTA。由结果可知,中心硅藻纲(centric ditom)的3 种硅藻,即假微型海链藻、威氏海链藻和角毛藻中检测到的甜菜碱脂主要为DGCC,没有检测到DGTA,而在羽纹硅藻纲(pennate diatom)的两种硅藻小新月菱形藻和三角褐指藻中检测到甜菜碱脂仅有DGTA。根据Vogel 和Eichenberger[11]的研究,DGTS 是DGTA 合成的前体,即DGTA 是通过DGTS 极性头部脱羧基和重新形成羧基,以及甘油分子脱酰化和再酰化形成的。因此,所有含有DGTA 的微藻原则上都应含有少量或痕量的DGTS,DGTS 的水平取决于它转换成DGTA 的速率。而小新月菱形藻和三角褐指藻中仅检测到DGTA,可能是因为DGTS 含量太低,没有达到相应分析的检测限。双眉藻(Amphorasp.)属于羽纹硅藻纲的硅藻,Li 等[17]在双眉藻(Amphorasp.)中共检测到16 种DGTA和9 种DGTS,没有检测到DGCC,且DGTA 的种类和含量多于DGTS。说明这种生化组成的不同有助于鉴别硅藻的两个不同纲,相关的研究也证明了这一结论[9–10,15,18]。因此,海洋微藻进化过程中两个不同纲的硅藻甜菜碱脂的合成机制仍需要大量的研究去探索。
绿藻门的3 种微藻中检测到的甜菜碱脂仅为DGTS,其种类和含量远远高于其他门微藻中的DGTS。目前关于甲藻门中甜菜碱脂分布的研究较少,Cañavate 等[15]在环沟藻中仅检测到1 种DGCC(16:0/16:0),Flaim 等[19]和Anesi 等[20]在甲藻中仅检测到DGCC 存在。本研究中,东海原甲藻和微小亚历山大藻分别发现3 种和10 种DGCC,而在卡罗藻中同时检测到3 种甜菜碱脂的存在,即11 种DGCC、2 种DGTS 和2 种DGTA。根据已有报道,本研究的卡罗藻是目前唯一报道过的同时含有3 种甜菜碱脂的甲藻。
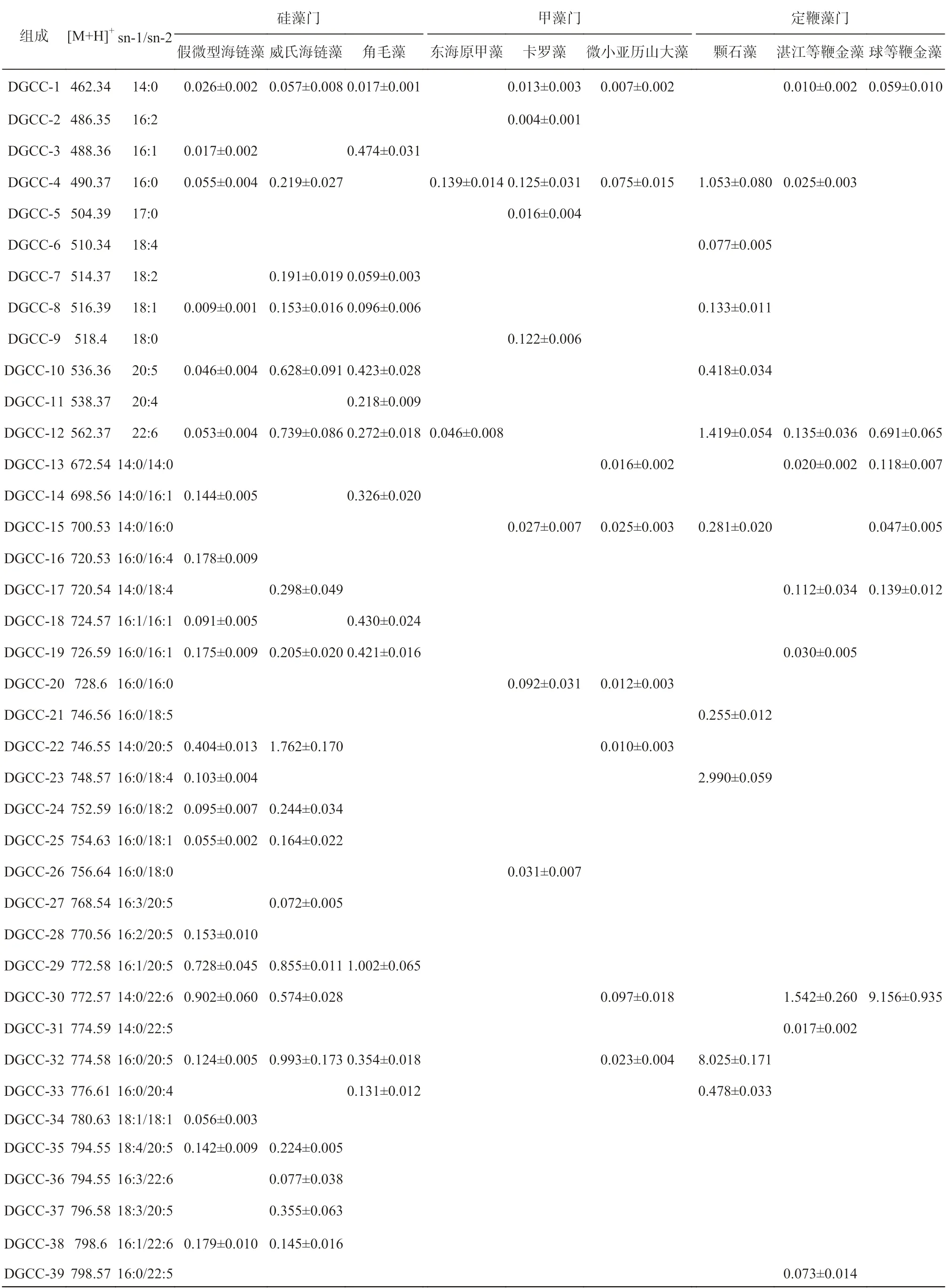
表2 9 种海洋微藻中DGCC 的组成及含量(nmol/mg 干藻粉)Table 2 The composition and content of DGCC in 9 microalgae(nmol/mg dry microalgae)
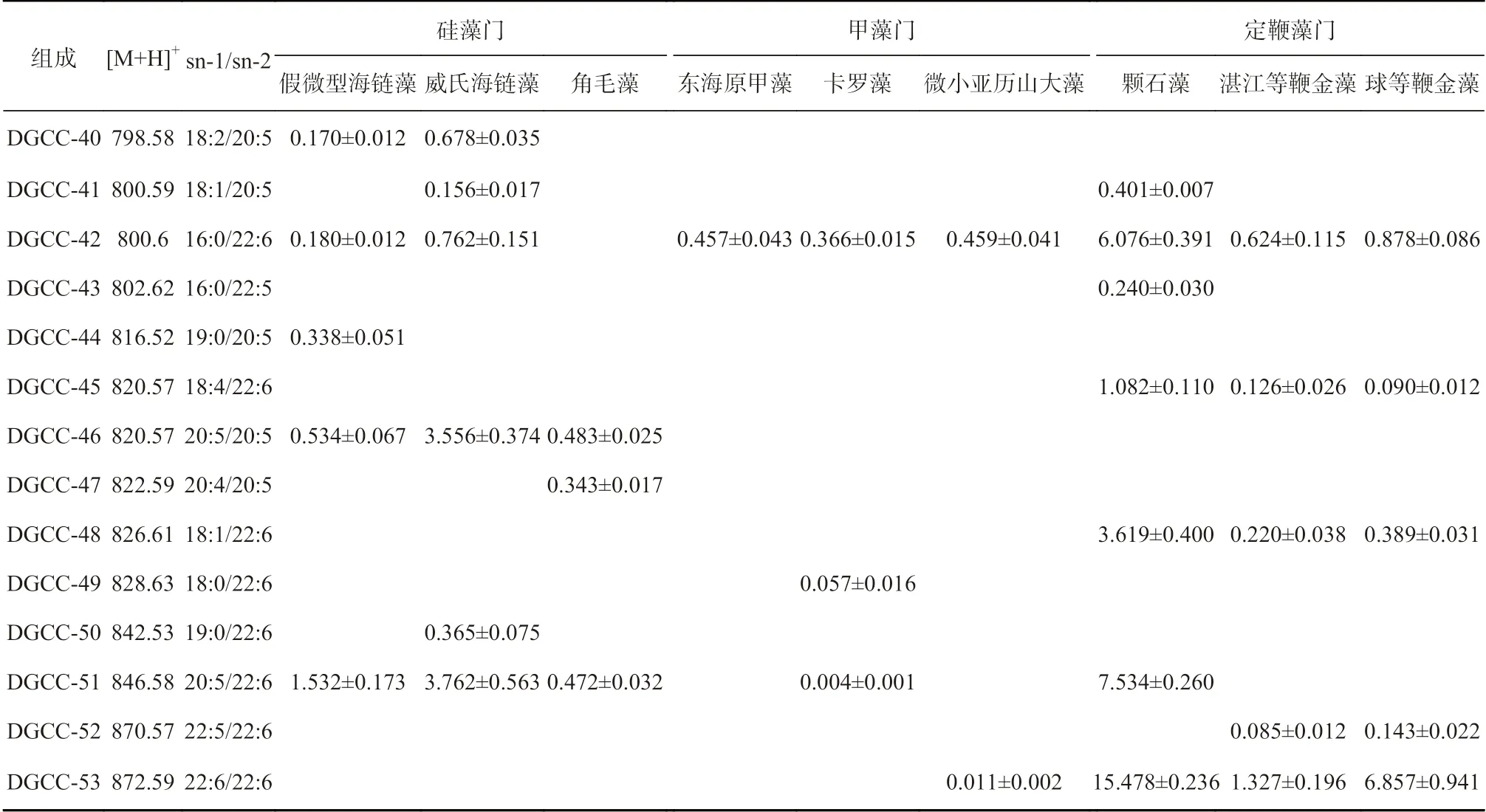
续表2
Cañavate 等[15]利用主坐标分析(PCO)方法对微藻中的甜菜碱脂进行研究,结果表明绿藻纲、共球藻纲和大眼藻纲的微藻聚在第一坐标轴,与DGTS 含量成正相关;DGTA 和DGCC 在第二坐标轴上分开,隐藻门和硅藻门的甜菜碱脂主要是DGTA,间藻门和甲藻门唯一的甜菜碱脂为DGCC。包括本研究在内的甜菜碱脂的研究都仅是一个初步的研究,还不能全面了解甜菜碱脂在微藻中的分布,但可以将甜菜碱脂作为完整极性脂的生物标记物,用于评估浮游植物天然的种群结构和生理状态[21]。
3.3 微藻中甜菜碱脂的脂肪酸组成
为了更好地了解不同门的微藻中甜菜碱脂分子的脂肪酸组成特点,将本研究中甜菜碱脂的脂肪酸组成检测结果与部分文献资料[3,15,18,22]报道过的微藻中甜菜碱脂的脂肪酸组成相结合,对甲藻门、绿藻门、定鞭藻门和硅藻门四大类微藻中甜菜碱脂的脂肪酸组成进行了总结(表5)。由结果可知,硅藻门的甜菜碱脂DGTS 中脂肪酸组成主要是C14−C18 偶数个碳原子的饱和以及低不饱和短链脂肪酸,而DGTA和DGCC 中的脂肪酸组成除了短链脂肪酸外,还含有多不饱和的脂肪酸和长链的多不饱和脂肪酸,如C20:4、C20:5 和C22:6,该结果与Li 等[17]对双眉藻Amphorasp.的脂肪酸组成研究结果一致。定鞭藻门3 种类型甜菜碱脂的脂肪酸组成差别不大,基本为C14 脂肪酸、C16 脂肪酸、C18 脂肪酸以及C20:5 脂肪酸和C22:6 脂肪酸;甲藻门中DGCC 脂肪酸组成为C14 饱和短链脂肪酸、C16 饱和短链脂肪酸、C17 饱和短链脂肪酸和C18 饱和短链脂肪酸以及C20:5 多不饱和脂肪酸和C22:6 多不饱和脂肪酸,而DGTA 和DGTS 中仅含有短链脂肪酸。
此外,对不同门微藻中同一种类型甜菜碱脂的脂肪酸分布进行了分析,结果表明,硅藻门、甲藻门和定鞭藻门中均含有DGCC,但是甲藻门C14−C18 脂肪酸链均为饱和的,而硅藻门中含有C19 奇数碳原子的脂肪酸链。对于DGTS,硅藻中主要含有C14−C18的脂肪酸,甲藻中只含有C18 的脂肪酸,C20 的脂肪酸仅在绿藻门中检测到,而C22:6 脂肪酸仅在定鞭藻中发现,这些特殊的脂肪酸组成的甜菜碱脂可在水产食物网研究中用作营养标记。
以上研究结果表明,不同门的微藻甜菜碱脂的脂肪酸组成有其各自的特点,同一个门内的不同微藻间甜菜碱脂脂肪酸的组成差别不大。相关研究表明,海洋大型植物中脂肪酸分布在科的水平上就可以很好地区分[23],而微藻中甜菜碱脂的脂肪酸分布的特点与海洋大型植物中脂肪酸分布存在不同。Dalsgaard 等[24]研究表明,微藻的脂肪酸组成在较高的分类水平上表现出一定的特点,在水产食物网研究中被广泛用作营养标记;Thomas 等[25]研究也表明不同微藻脂肪酸组成差异只能限制在门和纲的水平上,在较低的分类水平上差异不明显。甜菜碱脂作为一种完整的极性脂已用于浮游植物自然群落结构的研究[26–27],对甜菜碱脂脂肪酸组成的研究有望提高不同物种区分的分辨度,可以为微藻的系统发育及化学分类提供新的见解。

表3 8 种海洋微藻中DGTS 的组成及含量(nmol/mg 干藻粉)Table 3 The composition and content of DGTS in 8 microalgae(nmol/mg dry microalgae)

表4 3 种海洋微藻中甜菜碱脂的组成及含量(nmol/mg 干藻粉)Table 4 The composition and content of betaine lipid in 3 microalgae(nmol/mg dry microalgae)

表5 4 个门的微藻中甜菜碱脂脂肪酸的组成特点Table 5 Characteristics of fatty acids of betaine lipids in marine microalgae
4 结论
本研究运用超高效液相色谱−四级杆−飞行时间质谱建立了海洋微藻中甜菜碱脂的分析鉴定方法,对4 个微藻门中的14 种微藻的甜菜碱脂分子结构及其相应脂肪酸的组成及分布进行了研究。研究结果表明,这种简单快速的方法可以同时检测出微藻中3 种类型的甜菜碱脂。不同门的微藻中甜菜碱脂分布的种类不同,定鞭藻和甲藻中主要的甜菜碱脂是DGCC,绿藻中主要的甜菜碱脂为DGTS,而在硅藻中的甜菜碱脂主要包括两种,即DGCC(中心硅藻纲)和DGTA(羽纹硅藻纲)。不同门的微藻甜菜碱脂的脂肪酸组成有其各自的特点,根据脂肪酸组成特点对微藻分类只能限制在门和纲的水平上,在较低的分类水平上差异不明显。本研究对微藻中甜菜碱脂的种类和分布特性进行了研究,对于进一步研究这些缺磷极性脂类的生理、生态作用和功能特性具有重要的意义,也可以为微藻的系统发育及化学分类提供新的见解。
