云斑斜线天蛾在我国的适生性及限制性环境因子分析
2021-01-13刘博王晔楠唐超刘丽马光昌彭正强阎伟
刘博 王晔楠 唐超 刘丽 马光昌 彭正强 阎伟



摘 要:云斑斜线天蛾是一种为害抗风桐的迁飞性昆虫,在我国西沙群岛多个岛屿发生。阐明云斑斜线天蛾在我国的适生区分布及其主要限制环境因子,可为该害虫扩散预警与防控提供理论依据。本文利用MaxEnt模型对云斑斜线天蛾在我国的潜在适生区进行预测,通过调整调控倍频和特征组合参数建立最优模型,使用刀切法及环境变量响应曲线对影响云斑斜线天蛾分布的环境因子进行评估。结果表明:云斑斜线天蛾在我国的潜在适生区位于海南、台湾、广东、广西、云南、贵州、湖南、江西、福建、浙江、安徽、湖北、四川、重庆、西藏;最冷月最低温(bio06)、昼夜温差月均值(bio02)、最暖季降水量(bio18)是影响云斑斜线天蛾潜在地理分布的主导环境因子。云斑斜线天蛾在我国南海诸岛具有较高危险性,建议建立监测预警与防控系统,将该害虫控制在合理水平。
关键词:云斑斜线天蛾;MaxEnt模型;适生区;南海诸岛;抗风桐
中图分类号:S31 文献标识码:A
Abstract: Hippotion velox is a migratory insect that damages Pisonia grandis. It occurs in many islands of China Paracel Islands. MaxEnt model was used to to clarify the potential suitable area and the main environmental factors of H. velox in China and to provide a theoretical basis for the early warning and prevention of the insect. An optimal model by adjusting the regularization multiplier and feature combination parameters was established. The environmental factors affecting the distribution of H. velox were evaluated using the jackknife method and the response curve of environmental variables. The potential suitable area of H. velox are distributed in Hainan, Taiwan, Guangdong, Guangxi, Yunnan, Guizhou, Hunan, Jiangxi, Fujian, Zhejiang, Anhui, Hubei, Sichuan, Chongqing, Tibet. The min temperature of the coldest month (bio06), the mean diurnal range (bio02) and the precipitation of warmest quarter (bio18) are the dominant environmental factors affecting the potential suitable area of H. velox. H. velox has a high risk in the South China Sea islands. It is recommended to establish an early warning and control system to to control the pest at a reasonable level.
Keywords: Hippotion velox; MaxEnt model; potential suitable area; South China Sea Islands; Pisonia grandis
DOI: 10.3969/j.issn.1000-2561.2021.12.027
云斑斜线天蛾[Hippotion velox (Fabricius, 1793)]属鳞翅目(Lepidoptera)天蛾科(Sphingidae),是一种主要分布于亚洲和澳洲热带、亚热带地区的活跃的迁徙害虫[1]。目前已知的斜线天蛾幼虫寄主植物主要为紫茉莉科(Nyctaginaceae)避霜花属(Pisonia)、黄细心属(Boerhavia)、叶子花属(Bougainvillea),天南星科(Araceae)海芋属(Alocasia)、芋属(Colocasia),十字花科(Brass¬icaceae)芸薹属(Brassica),旋花科(Convol¬vulaceae)虎掌藤属(Ipomoea)及茜草科(Ru¬biaceae)巴戟天属(Morinda)等多种植物,如:抗风桐(Pisonia grandis)、腺果藤(Pisonia aculeata)、胶果木(Pisonia umbellifera)、Pisonia morindifolia、叶子花(Bougainvillea spectabilis)、热亚海芋(Alocasia macrorrhizos)、番薯(Ipomoea batatas)、海滨木巴戟(Morinda citrifolia)等[2-12]。云斑斜线天蛾在全球分布于中国(海南、广东、台湾、香港、广西、云南、贵州、四川)、日本、越南、泰国、缅甸、马来西亚、菲律宾、印度尼西亚、印度、斯里兰卡、澳大利亚、新喀里多尼亚、巴布亚新几内亚、所罗门群岛、北马里亚纳群岛、关岛、帕劳、斐济、瓦努阿图[13-14]。
抗风桐(Pisonia grandis)属紫茉莉科常绿乔木,是西沙群岛自然森林群落的第一优势种,在多个岛屿上都有分布,常形成纯林,其生长速度快,断枝可再植,在防风、海岸固沙以及海岛植被恢复方面具有重要作用[15]。在澳大利亚昆士兰州图勒加和北领地达尔文,云斑斜线天蛾为害会导致Pisonia和Bougainvillea屬寄主植物不同程度的落叶;在澳大利亚昆士兰州桅顶岛、东北先驱礁、东南马格德莱恩礁等岛礁抗风桐林爆发,会造成抗风桐完全脱叶,其危害爆发呈现一定的周期性[9]。云斑斜线天蛾在我国西沙群岛和东沙群岛主要为害抗风桐和海滨木巴戟,并在东岛、永兴岛、赵述岛、晋卿岛、珊瑚岛、琛航岛、广金岛、金银岛、中建岛、东沙岛等多个岛屿有分布[16]。该虫大发生时,会导致岛内抗风桐树叶全部被食光,对岛屿生态景观及生态系统稳定造成破坏。周先涌[17]于2017—2018年使用高空测报灯系统对永兴岛空中昆虫进行监测调查,根据总的诱虫量、恒有指数和年季间种群动态情况,认为云斑斜线天蛾在永兴岛属于关键性害虫。云斑斜线天蛾大发生呈现一定的周期性,但目前对该虫在我国的适生环境、迁徙特点及成灾机制知之甚少,亟需进行相关研究工作。分析云斑斜线天蛾在我国的适生性,对该虫的风险性评估、扩散预警与防控具有重要意义。
MaxEnt模型是一种基于最大熵理论预测物种地理分布的生态位评价模型[18]。因其具有所需样本量小、预测精度高等优点被广泛应用于外来入侵物种潜在分布区的研究[19-20]。本文利用MaxEnt模型對云斑斜线天蛾在我国的潜在适生区进行预测,通过调整调控倍频和特征组合参数建立最优模型,使用刀切法及环境变量响应曲线对影响云斑斜线天蛾分布的环境因子进行评估,以期为该虫的早期预警、风险评估、扩散与防控提供数据支撑。
1 材料与方法
1.1 数据收集
分布数据:通过全球生物多样性信息服务网络平台(Global Biodiversity Information Facility, GBIF)物种分布信息数据下载、国内外公开发表文献查阅及实地调查的方式,获得斜线天蛾在全球分布点数据[13]。
环境数据:从世界气象数据库(Worldclim, https://www.worldclim.org/)下载全球气候数据,该数据共包含1970—2000年的19个生物气候变量,版本为Version 2.0,空间分辨率为5 min(~10 km)。
地图数据:中国国界和省级行政区划图矢量底图(1∶4 000 000)来源于国家基础地理信息中心(http://ngcc.sbsm.gov.cn/)。
1.2 数据处理
1.2.1 分布数据 通过全球生物多样性信息服务网络平台等方式收集的物种分布数据,由于采集者与采集时间不同、物种分布地区的易到达程度、人为研究偏向性等因素会造成一定程度的采样偏差,这些分布数据可以造成分布模型的不确定性[21]。为了在一定程度上消除采样偏差对预测模型的影响,首先去除重合及不准确的分布点,然后在每个5′×5′的空间栅格数据中只保留一个分布点,经过筛选最终获得104个云斑斜线天蛾分布点用于模型预测。将分布数据按照MaxEnt格式要求保存为.CSV文件备用。
1.2.2 环境数据 气候数据基于温度和降水,环境变量间存在自相关和多重线性重复等问题[22],为减少变量间冗余信息对模拟结果的干扰,需对环境变量进行降维处理。使用R软件对云斑斜线天蛾分布点环境变量数据进行Pearson相关性分析,对于∣r∣≥0.75的环境变量,参考初次建模时环境变量贡献率,只选择一个代表性的变量作为预测模型的环境变量[23]。经过筛选最终使用昼夜温差月均值(bio02)、等温性(bio03)、最暖月最高温度(bio05)、最冷月最低温度(bio06)、年降水量(bio12)、最干月降水量(bio14)、最暖季降水量(bio18)等7个环境变量用于模型预测。
1.3 模型设置
MaxEnt采用机器学习算法,在默认参数条件下,模型对采样偏差敏感,易产生过度拟合,需要采取适当的措施来限制模型的复杂性和过度拟合,提高模型转移能力[24]。利用kuenm数据包调整MaxEnt模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)参数,调控倍频设置为1~4,递增0.1,共31个水平,特征选择线性(linear-L)、二次型(quadratic-Q)、乘积型(product-P)、阈值型(threshold-T)、片段化(hinge-H)5种参数,31种特征组合,分析各种参数条件下模型的复杂度,选择最优组合参数用于模型预测[25]。根据最佳模型标准选择统计上显著、低遗漏率及低ΔAICc值的候选模型,最终确定调控倍频1.5与特征组合LQP为最优模型参数。将筛选处理后的分布数据和环境变量文件导入MaxEnt软件,设置线性、二次型、乘积型特征参数,调控倍频设置1.5。随机选取25%的分布点作为测试集,剩余的75%分布点作为训练集,重复训练次数(replicates)设置为10,重复运行模式选择“Bootstrap”;创建响应曲线(create response curves)、绘制变量重要性刀切图(do jackknife to measure variable importance),预测结果使用“Logistic”格式和“asc”文件类型输出。
1.4 模型评价
ROC曲线(receiver operating characteristic curve)以真阳性率为纵坐标,假阳性率为横坐标所形成的曲线,曲线分析通过改变阈值,获得多对真(假)阳性率值。AUC(area under the ROC curve)值是ROC曲线与横坐标围成的面积,AUC值越大表示与随机分布相距越远,环境变量与预测的物种地理分布模型之间的相关性越大,模型预测效果越好,AUC值不受阈值影响,评价更客观[19]。ROC曲线的评估标准为:AUC值低于0.5,失败;0.5~0.7,较差;0.7~0.9,一般;0.9~1.0,很好[26]。
遗漏率(omission rate, OR)是指测试样本中没有被正确分类的异常样本数与测试样本中异常样本总数的百分比,其值越小表示预测准确性越高[27]。遗漏率提供模型差别和过度拟合的信息,对使用的数据在特定的阈值上进行评估。根据累积阈值的定义,良好的模型,测试遗漏率应该与预测遗漏率保持一致,如果测试遗漏率过高或低于理论遗漏率,说明用于测试和训练的数据是不独立的,建模数据存在空间自相关[28]。通过计算最低存在阈值(lowest presence threshold)遗漏率OR0和第10百分位存在阈值(10th percentile presence threshold)遗漏率OR10来评价模型灵敏度。理想模型OR0和OR10值分别为0和0.1,超过理想模型OR值,则认为模型存在过度拟合[29]。
1.5 适生区确定
将MaxEnt模型预测结果文件导入ArcGIS中,经过转栅格、提取分析,获得云斑斜线天蛾在中国的适生性栅格图。利用ArcGIS“自然間断点分级法(Jenks)”将适生性栅格图划分为4个适生区:高度适生区、中度适生区、低度适生区及非适生区[30]。利用ArcGIS空间分析功能统计各适生区的面积。
1.6 刀切法测试
变量重要性刀切图用来判断各生物气候变量对分布预测的贡献,可以帮助筛选主导气候因子。模型生成的各气候因子响应曲线显示预测分布概率随环境变量的变化趋势,响应曲线反映各环境变量对模型预测的影响[28]。
2 结果与分析
2.1 模型准确性检验
ROC曲线(图1)结果显示,测试集AUC值为0.981,标准差为0.002,依据AUC曲线评价标准,本次预测模型达到“很好”效果。OR0值为0.009,OR10值为0.105,非常接近理想模型OR值,表明模型拟合情况较好。AUC值与OR值的检验结果说明模型具有较高的灵敏度和准确性。
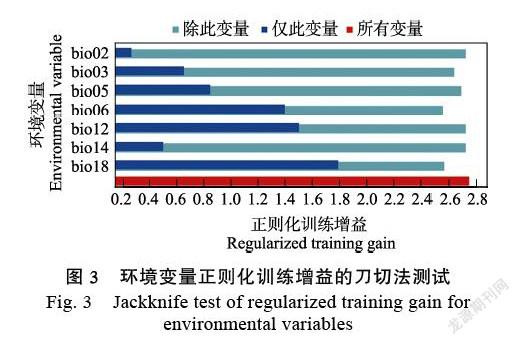
2.3 刀切法测验与主导环境因子
根据MaxEnt模型训练集环境变量正则化增益刀切图(图3)显示,使用单环境变量建模时,最暖季降水量(bio18)模型具有最高的正则化增益值,其值为1.799,说明最暖季降水量(bio18)具有最多影响云斑斜线天蛾分布的有效信息。去除单个环境变量最冷月最低温(bio06)时,模型增益减少最多,其值为2.564,说明最冷月最低温(bio06)具有最多其他变量没有的影响云斑斜线天蛾分布的信息。
MaxEnt环境变量贡献分析结果(表1)显示,最冷月最低温(bio06)、昼夜温差月均值(bio02)、最暖季降水量(bio18)的贡献率分别为43.3%、17.9%、16.9%。当环境变量对模型的贡献率大于15%时,则认为该气候变量是影响物种分布模型结果的重要变量[31],故最冷月最低温(bio06)、昼夜温差月均值(bio02)、最暖季降水量(bio18)是影响云斑斜线天蛾潜在地理分布的主导环境变量。
主导环境变量的响应曲线结果(图4)显示:云斑斜线天蛾分布概率随最冷月最低温(bio06)的增加而提高,当bio06值低于–10 ℃时,分布概率趋于0,当bio06值高于10 ℃时,分布概率显著提高,当bio06值高于24 ℃后分布概率达到最大值。分布概率随昼夜温差月均值(bio02)的增加而逐渐降低。最暖季降水量(bio18)曲线呈单峰曲线,极值约为1300 mm,高于或者低于该值时,分布概率降低,当bio18值低于0 mm或高于3000 mm时,分布概率趋于0。由主导环境变量响应曲线结果说明,云斑斜线天蛾更适应昼夜温差较小的较为湿润的温热环境。
3 讨论
MaxEnt预测模型基于最大熵理论,首先通过物种已知分布区域的空间特征,寻找限制物种分布的约束条件(环境变量),然后构筑约束集合,最后建立二者之间的相互关系[18]。MaxEnt模型在物种样本量较少的情况下仍有较高的准确率,现已被广泛应用于入侵物种潜在分布区的研究[32-34]。MaxEnt采用机器学习算法,模型对采样偏差敏感,易产生过度拟合,通过调整特征组合和调控倍频参数可以约束模型的复杂度。根据最优模型评估标准,选择调控倍频为1.5,特征组合为LQP用于模型预测。建模后得到云斑斜线天蛾在我国的适生区,模型测试集AUC值达到0.981,标准差为0.002,表明预测模型准确性高,预测效果“很好”。MaxEnt预测结果表明,云斑斜线天蛾在我国的适生区主要位于长江以南地区,环境变量贡献率及刀切法测试结果显示,最冷月最低温、昼夜温差月均值、最暖季降水量是影响云斑斜线天蛾分布的主导环境因子,主导环境变量响应曲线表明,云斑斜线天蛾更适应昼夜温差较小的较为湿润的温热环境。
物种的地理分布受多种环境变量的限制,是物种与环境长期相互作用的结果。影响物种分布格局的因素主要有气候因素、生物地理因素、物种自身的迁移能力、物种间的相互作用、物种生态幅及人为因素等[35]。气候因素、物种间的相互作用和物种的迁移能力是影响物种分布的3个主要因素,气候因素(温度和降水等相关因子)主要在大尺度空间影响物种的分布,物种间的相互作用及物种自身的迁移能力主要在较小的空间尺度下影响物种的分布[36]。就本研究的空间尺度而言,物种间的相互作用和物种自身的迁移能力对物种大尺度的分布范围影响相对较小,而对小尺度范围内的种群动态、短距离的迁移和危害严重程度等有较大影响。云斑斜线天蛾最嗜寄主为抗风桐等紫茉莉科避霜花属植物,云斑斜线天蛾的发生记录分布与抗风桐的全球分布具有较高的重合度,表明云斑斜线天蛾的适生范围可能与抗风桐等避霜花属植物的分布密切相关。本研究使用MaxEnt生态位模型仅利用物种分布点所关联的气候变量来模拟物种的潜在分布范围,模拟的生态位属Grinnell生态位(grinnellian niche),该范围小于基础生态位(fundamental niche),大于现实生态位(realized niche)。除气候因素外,寄主、天敌和人类活动等因素也对云斑斜线天蛾的分布与发生产生影响,这些因子作为环境变量可以提高物种分布模型的真实性和准确性。
我国南海诸岛基本上都是珊瑚岛,其中西沙群岛自然岛屿最多,植被最具代表性,自然乔木植被主要是抗风桐纯林和海岸桐纯林。抗风桐具有耐强光、干旱和贫瘠等热带珊瑚岛礁环境的生态适应性,根部的根瘤菌能够进行生物固氮,利于海岛植物繁育以及海岛土壤养分的提升,是热带珊瑚岛礁环境的主要植被恢复工具种[37]。云斑斜线天蛾是一种分布于东洋界和澳新界沿海地区及岛屿的迁飞性害虫,该虫在西沙群岛严重危害抗风桐,导致整株叶片全部脱落。云斑斜线天蛾在西沙群岛有周期性大发生的现象,近年来爆发危害的周期在缩短,危害发生更频繁。目前已知云斑斜线天蛾严重发生的地点多为热带、亚热带海岛,我国南海诸岛具有高度适宜的气候环境,云斑斜线天蛾在我国南海诸岛具有较高的潜在危险性,加之岛屿纯林生态系统的脆弱性,极易造成害虫的大爆发。云斑斜线天蛾是迁飞性害虫,极易随风扩散入侵其他岛屿,对我国南海诸岛森林生态系统具有潜在威胁。鉴于云斑斜线天蛾对我国南海诸岛森林生态系统的危险性,建议开展该虫的风险评估、扩散及成灾机制、绿色防控技术等方面的研究,建立适合热带岛礁的监测预警与防控系统,将虫害控制在合理水平,确保我国南海岛礁森林生态系统的健康与稳定。
参考文献
[1] Eitschberger U. Zur Kenntnis von Hippotion velox (Fab-ricius, 1793) (Lepidoptera, Sphingidae)[J]. Atalanta, 2015, 46: 153-164.
[2] Barnett L K, Emms C W, Holloway J D. The moths of the Chagos Archipelago with notes on their biogeography[J]. Journal of Natural History, 1999, 33(7): 1021-1038.
[3] Bell T R D, Scott F B. Fauna of British India, Including Ceylon and Burma, Moths, Vol. 5, Sphingidae[M]. Taylor and Francis Ltd., London, 1937: 415-417.
[4] Burwell C J, McDougall A, Nakamura A, et al. New but-terfly, hawkmoth (Lepidoptera) and dragonfly (Odonata) records from vegetated coral cays in the southern Great Barrier Reef, Queensland[J]. Australian Entomologist, 2011, 38(2): 75-88.
[5] Common I. Moths of Australia[M]. Melbourne University Press, 1990.
[6] Eitschberger, U, Ihle T. Raupen von schwärmern aus laos und Thailand -4. Beitrag (Lepidoptera, Sphingidae)[J]. Neue Entomologische Nachrichten, 2019, 76: 136-144.
[7] Hippotion velox (Fabricius, 1793)[DB/OL]. (2021-03-01). http://lepidoptera.butterflyhouse.com.au/sphi/velox.html.
[8] Iyer G, Kitching I J. A preliminary study of the hawkmoth diversity (Lepidoptera: Sphingidae) of Kanyakumari Dis-trict, Tamil Nadu, India[J]. Journal of Threatened Taxa, 2019, 11(5): 13592-13604.
[9] Moulds M, Tuttle J, Lane D. Hawkmoths of Australia: Identification, Biology and Distribution[M]. CSIRO Pub-lishing, 2020.
[10] Pittaway A R, Kitching I J. Sphingidae of the Eastern Pa-laearctic[DB/OL]. (2021-03-01). https://tpittaway.tripod.com/ china/h_vel.htm.
[11] Robinson G S, Ackery P R, Kitching I J, et al. Hostplants of the moth and butterfly caterpillars of the Oriental Re-gion[M]. Kuala Lumpur, Malaysia: United Selangor Press. 2001.
[12] Robinson G S, Ackery P R, Kitching I J, et al. HOSTS-a database of the world’s Lepidopteran hostplants[DB]. Natural History Museum, London, 2010.
[13] GBIF. Occurrence: Hippotion velox Fabricius, 1793[DB/OL]. [2021-03-11]. https://www.gbif.org/occurrence/search?q=Hi-ppotion%20velox%20(Fabricius,%201793).
[14] 蔣卓衡. 中国长喙天蛾属及斜纹天蛾属团的分类学研究[D]. 上海: 上海师范大学, 2019.
[15] 王馨慧, 刘 楠, 任 海, 等. 抗风桐(Pisonia grandis)的生态生物学特征[J]. 广西植物, 2017, 37(12): 1489-1497.
[16] 廖一璋, 江东权, 杨曼妙. 南海之岛—东沙昆虫相调查[J]. 国家公园学报, 2018, 28(1): 6-16.
[17] 周先涌. 海南省三沙市空中昆虫群落结构与种群动态[D]. 福州: 福建农林大学, 2019.
[18] Phillips S J, Anderson R P, Schapire R E. Maximum en-tropy modeling of species geographic distributions[J]. Ecological Modelling, 2006, 190(3): 231-259.
[19] 王运生, 谢丙炎, 万方浩, 等. ROC 曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4): 365-372.
[20] 陈新美, 雷渊才, 张雄清, 等. 样本量对MaxEnt模型预测物种分布精度和稳定性的影响[J]. 林业科学, 2012, 48(1): 53-59.
[21] 雷军成, 徐海根. 基于MaxEnt的加拿大一枝黄花在中国的潜在分布区预测[J]. 生态与农村环境学报, 2010, 26(2): 137-141.
[22] 朱耿平, 刘 强, 高玉葆. 提高生态位模型转移能力来模拟入侵物种的潜在分布[J]. 生物多样性, 2014, 22(2): 223-230.
[23] Kumar S, Neven L G, Yee W L. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment[J]. Ecosphere, 2014, 5(7): 1-23.
[24] 朱耿平, 原雪姣, 范靖宇, 等. MaxEnt模型参数设置对其所模拟物种地理分布和生态位的影响——以茶翅蝽为例[J]. 生物安全学报, 2018, 27(2): 118-123.
[25] Cobos M E, Peterson A T, Barve N, et al. Kuenm: an R package for detailed development of ecological niche models using MaxEnt[J]. PeerJ, 2019, 7: e6281.
[26] Peterson A T, Soberón J, Pearson R G, et al. Ecological niches and geographic distributions[M]. Princeton Uni-versity Press, Princeton, New Jersey, USA, 2011.
[27] 李一琳, 丁长青. 基于GIS和MaxEnt技术对濒危物种褐马鸡的保护空缺分析[J]. 北京林业大学学报, 2016, 38(11): 34-41.
[28] Phillips S J. A Brief Tutorial on MaxEnt[EB/OL]. (2017) [2019-04-28]. http://biodiversityinformatics.amnh.org/open_ source/maxent/.
[29] Radosavljevic A, Anderson R P. Making better MaxEnt models of species distributions: complexity, overfitting and evaluation[J]. Journal of Biogeography, 2014, 41(4): 629- 643.
[30] 刘 博, 覃伟权, 阎 伟. 基于MaxEnt模型的小巢粉虱在中国的潜在地理分布[J]. 环境昆虫学报, 2019, 41(6): 1276-1286.
[31] Oke O A, Thompson K A. Distribution models for moun-tain plant species: the value of elevation[J]. Ecological Modelling, 2015, 301: 72-77.
[32] 林 偉, 徐淼锋, 权永兵, 等. 基于MaxEnt模型的草地贪夜蛾适生性分析[J]. 植物检疫, 2019, 33(4): 69-73.
[33] 刘 博, 郑 奋, 王晔楠, 等. 危险性入侵害虫橘绵粉虱在中国的适生性预测[J]. 植物检疫, 2019, 33(4): 74-79.
[34] 卢 辉, 唐继洪, 吕宝乾, 等. 草地贪夜蛾的生物防治及潜在入侵风险[J]. 热带作物学报, 2019, 40(6): 1237-1244.
[35] 张晓芹. 西北旱区典型生态经济树种地理分布与气候适宜性研究[D]. 北京: 中国科学院大学(中国科学院教育部水土保持与生态环境研究中心), 2018.
[36] 朱耿平, 刘国卿, 卜文俊, 等. 生态位模型的基本原理及其在生物多样性保护中的应用[J]. 生物多样性, 2013, 21(1): 90-98.
[37] 于昕塍, 梁韩枝, 陈双艳, 等. 抗风桐的丛生芽诱导与再生[J]. 广西植物, 2021, 41(6): 890-896.
责任编辑:谢龙莲
