母体VD3 水平对仔猪背最长肌和背脂LPLmRNA 表达的研究
2021-01-11侯文博焦慧慧李亚欣刘永亮苗志国张金洲
侯文博,焦慧慧,李亚欣,刘永亮,苗志国,张金洲
(河南科技学院动物科技学院,河南新乡453003)
LPL(脂蛋白脂肪酶,Lipoprotein lipase)在调控脂蛋白运输和脂肪沉积方面发挥重要作用,具有将毛细血管内皮细胞表面血液循环乳糜微粒和极低密度脂蛋白所含的甘油三酯(Triglyceride,TG)水解为游离脂肪酸和作为脂蛋白摄取介导受体的双重功能.从Hocquette 等[1]研究可知:LPL mRNA 水平与其活性正相关.LPL 的活性决定脂肪沉积的程度,主要是通过控制肌肉和脂肪中TG 分配的数量来实现,特别是在肌内脂肪(Intramuscular fat,IMF)的合成和代谢进程中.王晶等[2]研究发现,鸡LPL 基因表达量和IMF 含量的发育性变化呈一致的正相关.另有研究表明,猪14 号染色体上LPL 基因突变对背部脂肪沉积、背膘厚有显著影响[3-5].脂肪沉积是影响肉质重要的因素之一.皮下脂肪沉积主要影响胴体瘦肉率,IMF 含量与猪肉的嫩度、风味、多汁性呈正相关[6-8].
VD3(维生素D3,Vitamin D3)属于类固醇脂溶性维生素,在生物学方面具有促进细胞生长分化、机体繁殖免疫和调节矿物质(钙磷)吸收代谢的功能.仔猪VD 缺乏时,钙磷吸收紊乱,进而引起骨骼钙化.Norman 等[9]研究发现,VDR 存在于许多和钙磷稳态无关的组织中,说明VD 可能有其他重要的生物学功能.目前大多数报道主要集中在日粮VD3水平对于断奶仔猪的生长功能、免疫功能、抗炎症功能的影响研究[10-12].但通过母体遗传效应改变仔猪脂肪代谢基因调控机制的研究鲜有报道.因此,通过开展母体不同VD3 水平对仔猪脂肪酸转运基因LPL 表达量的影响研究, 从而探求母体VD3水平影响子代LPL 基因表达规律和机制,以期在以后的生产生活中为通过改善母体VD3水平调控仔猪脂肪沉积、满足优良猪肉品质需求提供参考.
1 材料与方法
1.1 试验动物分组
选取由洛阳市伊川县新大牧业有限公司饲养的24 头体质量接近(144.6±2.3 kg)、健康的同期配种妊娠长白母猪,根据妊娠期间基础日粮VD3水平随机分为4 组,即对照组VD3-200 IU/kg、低水平VD3组-800 IU/kg、中等水平VD3组-3 200 IU/kg、高水平VD3组-6 400 IU/kg.泌乳期所有母猪基础日粮VD3水平保持相同,断奶后仔猪基础日粮营养成分保持一致.试验期保证相同的猪舍环境,包括温度、湿度、光照等.妊娠期、泌乳期母猪和仔猪各生长阶段基础饲粮配方参照2012 年NRC 配制.
1.2 采样
待仔猪六月龄时,从各组中随机选择3 个个体,共12 个个体按照《生猪屠宰操作规程》(1998)屠宰.采集背最长肌、背脂样品,一部分于-20 ℃保管,用于IMF 含量测定,剩余部分置于液氮中冷冻,然后移至-80 ℃储存,用于总RNA 的提取研究.
1.3 IMF 含量测定
采用索氏浸提法测定IMF 含量.利用石油醚提取单个背最长肌样品中的脂肪,每个样品测定3 次,以3 次测定的平均值代表样品的IMF 含量.
1.4 RT-PCR 相对定量测定
1.4.1 总RNA 的提取及cDNA 的合成 分别取黄豆粒大小背最长肌、背脂样品置于研钵中,倒入适量液氮,依照RNAAiso Plus 说明书使用TRIzol 提取总RNA.利用超微分光光度计测定纯度(D260nm/D280nm=1.8~2.0).使用RT-PCR 反转录试剂构建20 μL 反转录反应体系.具体步骤:加入RNA 模板、RNase Free dH2O、5×gDNA Eraser Buffer(2.0 μL)、gDNA Eraser 42℃(1.0 μL)共10.0 μL后于PCR 仪42 ℃反应2 min,再加入剩余试剂(10.0 μL)37 ℃反应15 min.
1.4.2 引物设计与合成 依据GenBank 发布的LPL 基因序列,以猪的β-actin 基因作为内参基因.利用Primer Premier 5.0 软件设计引物,由上海生工生物公司合成,引物序列信息见表1.

表1 引物序列信息Tab. 1 Primer sequence information
1.4.3 PCR 反应与电泳分析 以β-actin 作为内参基因,测定LPL 基因在背最长肌、背脂中相对表达量.RT-PCR 反应体系(25 μL):cDNA 模板1 μL、上下游引物各0.5 μL、TaqDNA 聚合酶12.5 μL、ddH2O 10.5 μL.PCR 仪反应程序:95 ℃预变性5 min;95 ℃变性30 s,57.5 ℃(表1)退火30 s,72℃延伸30 s,共32 个循环;最后72℃延伸5 min.每个样品设置3 个重复.取21 μLPCR 产物,以EB 染色的琼脂糖凝胶为载体在TAE 缓冲液中进行电泳.以目的基因LPL 与内参基因β-actin PCR 产物灰度值之比表示LPL基因的相对表达量.
1.5 数据处理与分析
运用SPSS 22.0 统计软件中的One-way ANOVA 对各组背最长肌肌内脂肪含量、LPL 基因表达和背脂LPL 基因表达差异进行单因素分析,结果采用平均值±标准差表示;对于不同VD3水平下背肌LPL基因表达量与IMF 含量双变量,使用SPSS 22.0 统计软件中的Bivariate Correlation 进行相关性分析.
2 结果与分析
2.1 背最长肌IMF 含量分析
由表2 可知,随母体VD3水平的增加,背最长肌IMF 含量总体呈下降趋势.对照组200 IU/kg IMF 含量显著高于试验组各水平(P<0.05);低水平试验组800 IU/kgIMF 含量显著高于高水平试验组6 400 IU/kg(P<0.05).

表2 各组背最长肌IMF 含量Tab. 2 IMF content of the longissimus dorsi in each group
2.2 背最长肌LPL 基因mRNA 的表达差异及其与IMF 含量的相关性分析
由图1 可知,仔猪背最长肌中LPL 基因mRNA 的表达随母体VD3水平的增加,总体呈先略微升高再逐渐下降的趋势.在低水平试验组(800 IU/kg)时LPL 表达量达到峰值,显著高于高水平试验组(6 400 IU/kg)(P<0.05);对照组(200 IU/kg)LPL 表达量显著高于高水平试验组(6 400 IU/kg)(P<0.05).其余各组之间差异不显著(P>0.05).
长白仔猪背最长肌LPL 基因表达量与IMF 含量的相关性分析表明:长白仔猪背最长肌LPL 基因mRNA 表达量与肌内脂肪含量总体上呈显著正相关,相关系数为R=0.679(P<0.05).见表3.

图1 母体不同VD3水平下仔猪背肌中LPL 基因的差异表达Fig. 1 Differential expression of LPL gene in the back muscle of piglets at different VD3 levels.

表3 背肌LPL 基因的表达与IMF 含量的相关性分析Tab. 3 Correlation Analysis of LPL gene expression and IMF content in dorsal muscle
2.3 背部脂肪LPL 基因mRNA 的表达差异分析
由图2 可知,随母体VD3水平的增加,仔猪背脂中LPL 基因mRNA 的表达呈先下降再逐渐上升的趋势.LPL 基因在低水平试验组(800 IU/kg)时表达量最低;在高水平试验组(6 400 IU/kg)时LPL 表达量达到峰值,明显高于低水平试验组(800 IU/kg)(P<0.05);其余各组之间差异不显著(P>0.05).
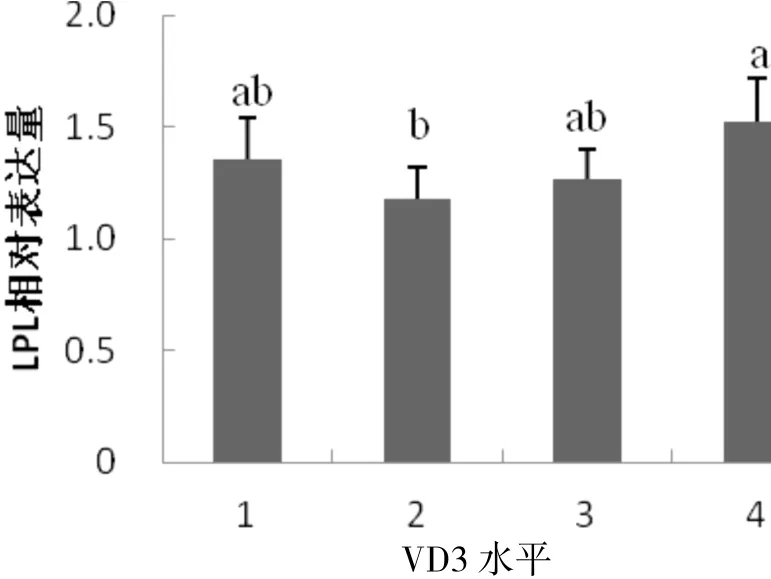
图2 母体不同VD3水平下仔猪背脂中LPL 基因的差异表达Fig. 2 Differential expression of LPL gene in piglet back fat at different vitamin D3 levels
3 讨论
早期研究表明,VD 在钙稳态和骨质代谢方面起着不可替代的作用.近年来,也有研究发现,VD 与肥胖、脂肪质量和组织功能、糖尿病有关[13-15].VD3 化学名称为胆钙化醇,是迄今为止在动物体内发现的VD的唯一天然成分.VD3 主要来源是阳光照射下皮肤合成,也可通过日常饮食摄取,与受体蛋白结合后,依次在肝脏、肾脏内经过羟基化,最终形成具有生物活性的1,25(OH)2D3[13,16].有关研究发现,VD 的分解代谢主要是在肝脏,所生成的25(OH)D3 是血液循环中VD 的主要存在形式,因此在临床上以血清中25(OH)D3 浓度来评估VD 水平[17-18].关于血清25(OH)D3 研究表明,其浓度与体质量指数(BMI)、脂肪质量呈负相关[19-21].
目前大多体外研究确定,VD3抑制3T3-L1 前脂肪细胞分化为成脂细胞,明显抑制3T3-L1 成脂细胞沉积脂肪[22-23].相关研究表明,1,25(OH)2D3 可诱导3T3-L1 脂肪细胞LPL 脂解活性,使其mRNA 表达水平明显提高,并且能够促进脂肪动员,增强脂肪分解,减少脂肪积累[24-25].另有研究发现,1,25(OH)2D3 通过剂量依赖抑制脂肪转录因子和脂肪生成晚期标志物(LPL、FAS 和脂质结合蛋白2)表达,抑制3T3-L1 前脂肪细胞分化和脂肪细胞生成脂肪[13,26].妊娠大鼠体内研究表明,VD3抑制了脂肪生成相关基因的表达,脂肪组织的脂肪沉积率显著降低[27].此外,VDR 作为VD 的受体同样参与3T3-L1 前脂肪细胞分化过程,而且高度表达时也会抑制前脂肪细胞分化.但有体内研究表明,VDR 可抑制脂肪分解基因(如激素敏感脂肪酶HSL),抑制脂质动员[28].由此可见,在脂肪沉积过程中,VD3及其受体对LPL 表达活性的影响可能与体内外研究结构层次、1,25(OH)2D3 剂量有关.
脂肪积累过程主要包括前脂肪细胞分化、脂肪细胞增殖分裂、脂肪细胞体积增大.LPL 作为分化初期阶段的标志基因之一,其降解转运外源性脂肪酸对促进脂肪细胞分化成熟有着重要的作用,最终直接影响脂肪沉积.张雄[29]对江香猪肌内脂肪构建预测模型研究表明:LPLmRNA 表达量与IMF 含量呈显著正相关(P<0.05).本研究通过控制长白妊娠母猪日粮VD3添加水平,发现六月龄仔猪背最长肌IMF 含量随母体VD3水平增加呈下降趋势,LPL mRNA 表达水平先略微上升后一直保持下降趋势,二者呈显著正相关(P<0.05).因此,推测VD3极有可能通过抑制肌肉中LPL 基因活性,减少了由脂解血液循环中乳糜微粒和极低密度脂蛋白供应肌肉的游离脂肪酸量,从而影响IMF 含量.另外,本研究还发现仔猪背脂LPLmRNA 随母体VD3水平先下降后逐渐升高.LPL 基因测序研究表明,猪和鼠LPL 基因同源性高达88.7%.LPL 基因在脂肪和肌肉中的表达差异极大,以鼠为研究对象表达甚至是相悖的[30].由此推断,本研究中LPL 基因在背最长肌与背脂中表达趋势差异大,这可能与LPL 基因本身在肌肉和脂肪组织中作用机制不同有关,也可能是VD3水平使肌肉和脂肪组织内LPL 基因表达活性发生变化的阈值不同.本研究未测定仔猪其余部位肌肉、脂肪中LPL 基因mRNA 变化趋势,因此探究母体VD3水平使仔猪肌肉和脂肪内LPL 基因表达变化规律还需要进一步讨论和总结.
4 结论
长白仔猪背最长肌和背脂中LPL 基因表达趋势不同,背最长肌中随母体VD3水平的增加,LPL 基因mRNA 表达水平先上升后逐渐下降,在VD3水平为800IU/kg 时达到峰值,且显著高于VD3水平为6400IU/kg表达水平(P<0.05);背脂中LPL 基因mRNA 表达水平先下降后上升,在VD3 水平6 400 IU/kg 时最高,显著高于800 IU/kg 表达水平(P<0.05).背最长肌LPLmRNA 表达量与肌内脂肪含量呈显著正相关(R=0.679,P<0.05).本研究表明,通过母体遗传效应,VD3水平升高显著抑制了背最长肌中LPL 基因表达,进而IMF含量降低,脂肪沉积减少.
