黔北麻羊FABP3基因多态性及其与生长性状的相关性研究
2021-01-08艾锦新龙安炬罗卫星蔡惠芬
艾锦新,龙安炬,罗卫星,蔡惠芬*
(1. 贵州大学动物科学学院/贵州省动物遗传育种与繁殖重点实验室,贵州 贵阳 550025;2. 贵州大学高原山地动物遗传育种与繁殖教育部重点实验室,贵州 贵阳 550025)
心脏型脂肪酸结合蛋白(heart fatty acid binding protein, H-FABP)是脂肪酸结合蛋白(FABP)家族中重要的一员,又名FABP3,是一种低分子量胞浆蛋白,主要在心肌、骨骼肌和脂肪细胞中表达[1-3]。FABP3基因主要通过参与胞内长链脂肪酸的摄取、转运和利用,调节脂肪代谢;在脂肪生成过程中,FABP3基因通过在心肌和脂肪细胞中沉积甘油三酯,从而促进肌内脂肪(intramuscular fat, IMF)沉积,增加IMF含量[4-5]。山羊的FABP3基因定位于2号染色体上,由5个外显子和4个内含子组成[6]。在基因多态性上,余刚等[7]研究发现,FABP3基因第2外显子的SNP与陕北绒山羊的生长性状(胸围、管围)、胴体性状(眼肌面积、背膘厚度)显著相关。罗燕等[8]研究发现,FABP3基因外显子2存在多态位点且具有3种基因型AA、AB和BB,其中BB基因型是影响中国美利奴羊IMF的优势基因型。曲亮等[9]研究发现,FABP3基因多态性与苏淮猪的胴体性状极显著相关。王珊等[10]研究发现,FABP3基因HaeⅢ位点存在多态性,各基因型秦川牛及其杂种牛的体质量、胸围存在显著差异。在基因表达水平上,Huang等[11]研究发现,30~90日龄的公哈萨克绵羊肌肉组织FABP3基因的mRNA表达水平与肌间脂肪含量呈极显著的强正相关。岳彩娟等[12]研究发现,FABP3基因的表达量与滩羊背最长肌、股二头肌及腰大肌的IMF均呈正相关。沙尔山别克·阿不地力大等[13]研究发现,不同发育阶段拜城油鸡的肌肉组织中FABP3基因mRNA表达量与其IMF含量呈一定的相关性。这些研究结果表明,FABP3基因可能是影响动物生长发育的重要基因。
黔北麻羊是贵州省三大优良地方山羊品种之一,其具有适应性强、耐粗饲、膻味轻、肉质鲜美、皮张品质好、生产性能优等特点,深受广大消费者和养殖户的青睐[14-15]。本研究以黔北麻羊为研究对象,对影响山羊肌肉发育和肉质相关性状的FABP3基因进行研究,在FABP3基因中检测影响黔北麻羊重要经济表型的基因突变,寻找与生长发育性状具有关联性的遗传标记,为后续的分子育种提供理论依据。
1 材料与方法
1.1 试验动物
随机选取贵州省习水县富兴牧业有限公司饲养管理条件相同、健康无病的6月龄黔北麻羊120只(公羊67只、母羊53只),颈静脉釆血8 mL,用肝素钠抗凝,置于冰盒带回实验室,-20 ℃保存备用。
1.2 主要试剂
血液基因组提取试剂盒购自北京天根生化科技有限公司;核酸染料、DL2000 Maker、2×EsTaqMasterMix均购自北京康为世纪生物科技有限公司;双蒸水、TAE缓冲液、琼脂糖凝胶均为实验室自备。
1.3 主要仪器
梯度PCR仪(Bio-rad公司);高速离心机(无锡瑞江分析仪器有限公司);超微量分光光度计(NANODROP 2000,Thermo公司);电泳槽和电泳仪(DYY-6C型,北京六一仪器厂);凝胶成像成像仪(Bio-rad公司)。
1.4 基因组DNA提取
试验采用血液基因组DNA提取试剂盒,分别对120头黔北麻羊血液中的基因组DNA进行抽提,并用超微量紫外分光光度计检测其浓度和纯度,1.2%琼脂糖凝胶电泳检测其完整性,取检测结果完好的DNA样品,-20 ℃保存备用。
1.5 引物设计及PCR扩增测序
根据GenBank发布山羊FABP3基因序列(NC_030809.1),利用Primer5.0引物设计软件设计5对特异性引物用于该基因全部外显子的扩增,引物详细信息见表1。PCR扩增反应体系为30 μL:2×EsTaqMasterMix 13 μL,DNA模板3 μL,上、下游引物各2 μL,双蒸水10 μL;反应程序为:95 ℃预变性5 min,95 ℃变性30 s,退火30 s(退火温度见表1),72 ℃延伸40 s,35个循环;72 ℃终延伸10 min,4 ℃保存。PCR产物用1.0%琼脂糖凝胶电泳检测,凝胶成像系统观察电泳结果,鉴定正确的PCR产物送往上海生物工程有限公司进行测序。

表1 引物信息
1.6 生长性状指标测定
按照文献[16-17]测定各试验羊的生长性状指标(包括体重、体高、体斜长、胸围、胸深、胸宽和管围),并做好数据记录。
1.7 SNP筛查与数据统计分析
测序结果用DNAStar软件进行序列分析和峰图比对,筛选出SNP位点;然后利用Microsoft Excel 2010软件统计各基因型在黔北麻羊群体中的分布情况,计算基因型频率、等位基因频率、纯合度(Ho)、杂合度(He)、有效等位基因数(Ne)、多态信息含量(PIC)及卡方值(χ2)。采用SPSS 18.0统计软件一般线性模型(GLM)对各基因型与生长性状指标进行差异显著性检验,以P<0.05为差异显著水平,结果均以“最小二乘均值±标准误”表示。一般线性模型如下:
Yij=μ+&ij+bij+&ij×bij+eij,
式中,Yij表示性状表型值,μ表示总体均值,&ij表示基因型效应,bij表示品种效应,eij为随机差。
2 结果与分析
2.1 FABP3基因PCR扩增
以黔北麻羊FABP3基因组DNA为模板,分别扩增FABP3基因全部外显子区,1.0%琼脂糖凝胶电泳进行检测,结果见图1。由图1可知,扩增产物与目的片段大小一致,条带清晰明亮,无非特异性扩增,无明显拖尾现象,说明引物特异性良好,PCR产物可直接用于测序。
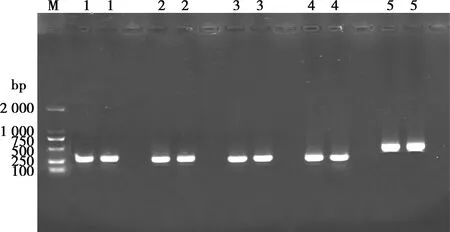
M.DL2000 DNA Marker;1~5.引物分别为P1、P2、P3、P4、P5,每对引物2个重复图1 黔北麻羊FABP3基因产物电泳
2.2 PCR产物测序及序列分析
PCR产物进行直接测序,测序结果经序列比对分析,在黔北麻羊FABP3基因第5外显子区域筛选出1个SNP位点(C/T),命名为g.13665051C>T,该突变未引起编码氨基酸发生改变,属同义突变,产生3种基因型:CC、CT和TT;同时在黔北麻羊FABP3基因3′非翻译区(3′UTR)也筛选出1个SNP位点(G/A),命名为g.13664737G>A,该突变产生3种基因型:GG、AG和AA,测序峰如图2所示。
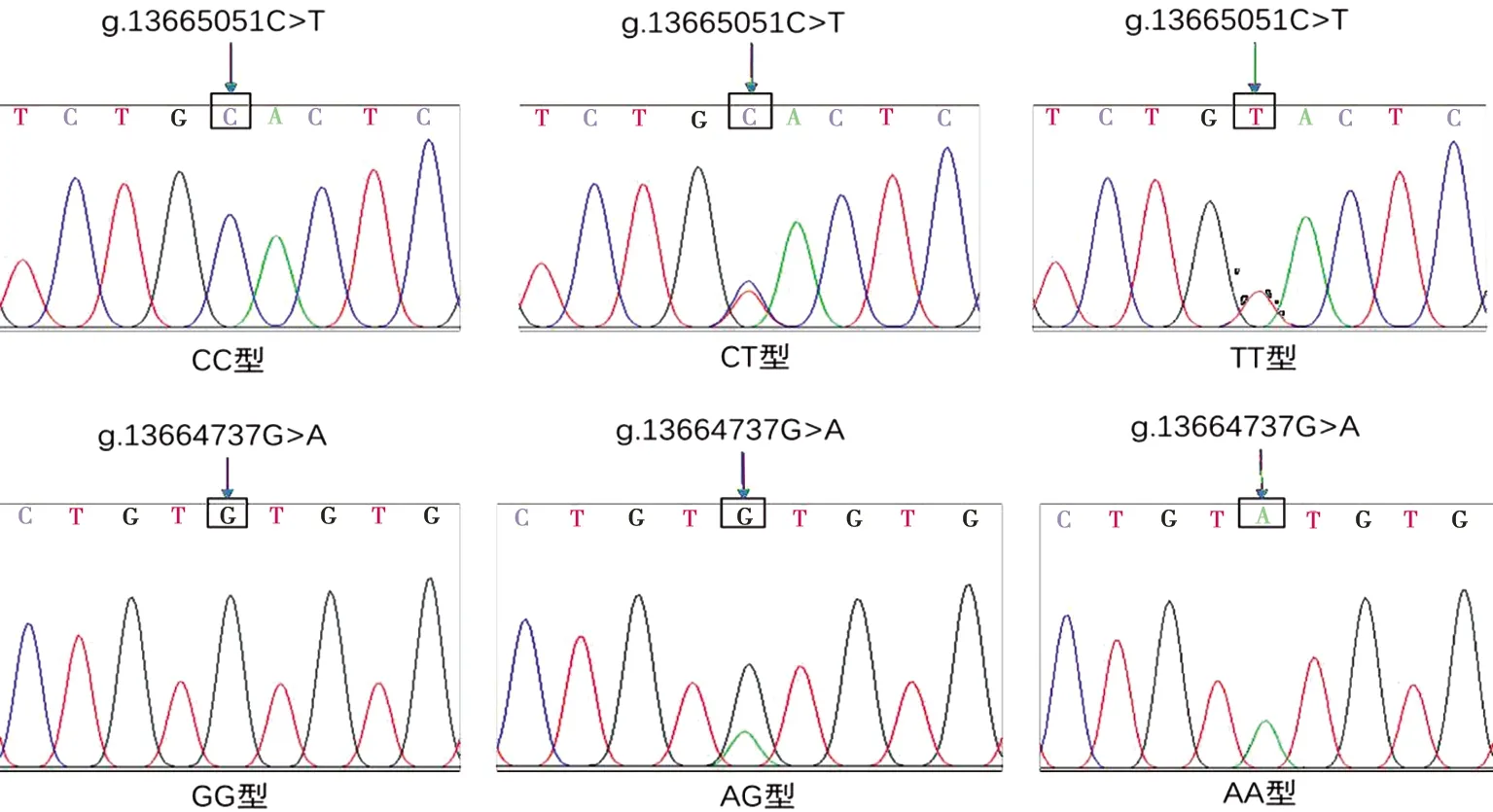
图2 黔北麻羊FABP3基因单个样本测序峰
2.3 FABP3基因遗传学分析
对黔北麻羊FABP3基因2个SNPs位点进行群体遗传特性分析,结果见表2。由表2可知,黔北麻羊FABP3基因g.13665051C>T位点中CC为优势基因型,等位基因C为优势等位基因,该基因座基因纯合度为0.562,杂合度为0.438,有效等位基因为1.779,多态信息含量为0.341,属中度多态位点;黔北麻羊FABP3基因g.13664737G>A位点中GG为优势基因型,等位基因G为优势等位基因,该基因座基因纯合度为0.531,杂合度为0.469,有效等位基因为1.883,多态信息含量为0.359,属中度多态位点。卡方(χ2)检验结果显示,g.13665051C>T和g.13664737G>A突变位点的基因型分布均未偏离Hardy-Weinberg平衡(P>0.05)。

表2 SNPs位点在黔北麻羊中的群体遗传信息
2.4 FABP3基因不同基因型与生长性状的关联分析
对黔北麻羊FABP3基因SNPs位点与生长性状进行关联分析,结果见表3。由表3可知,在黔北麻羊群体中,FABP3基因g.13665051C>T位点中CC、CT和TT基因型个体间的体重差异不显著(P>0.05),CT基因型个体的体高、体斜长、胸宽和管围显著高于CC和TT基因型个体(P<0.05),CC和CT基因型个体的胸围、胸深显著高于TT基因型个体(P<0.05),其中虽然CC和CT基因型个体的胸围、胸深无显著差异(P>0.05),但是CT基因型个体的胸围、胸深高于CC基因型个体;在FABP3基因g.13664737G>A位点中GG、AG和AA不同基因型的生长性状指标均差异不显著(P>0.05)。
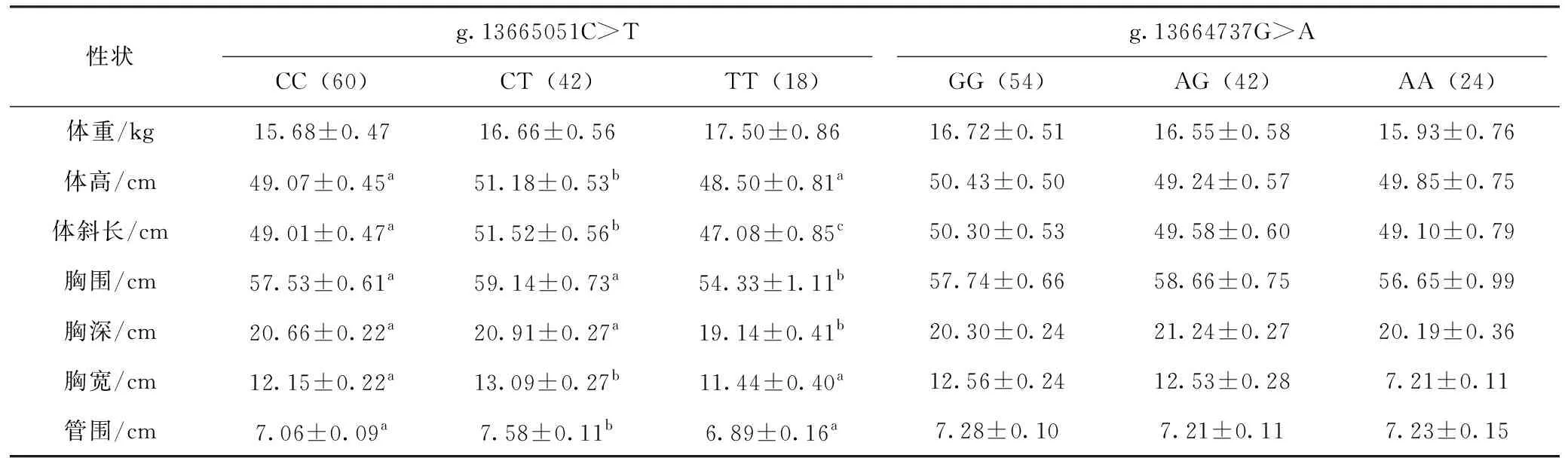
表3 黔北麻羊FABP3基因SNPs位点与生长性状的关联性分析
3 讨论
FABP3基因具有特异结合脂肪酸及调节脂肪酸代谢的重要功能,对畜禽的肉质性状、生长性状兼有影响[18]。本研究通过PCR产物直接测序技术对黔北麻羊FABP3基因的SNP进行检测,筛选出2个SNPs位点g.13665051C>T和g.13664737G>A,其中g.13665051C>T位于外显子5,产生3种基因型:CC、CT和TT;g.13664737G>A位于3′非翻译区,产生3种基因型:GG、AG和AA。其中g.13665051C>T突变是位于编码区的同义突变,编码区的同义突变虽不改变编码的氨基酸,但可能引起外显子拼接增强,从而影响蛋白质的表达量[19-20]。有研究表明,同义突变可以通过改变mRNA的二级结构[21]或改变核糖体通过mRNA特定区域时的速度[22]来改变蛋白质的结构和功能。因此,g.13665051C>T突变仍可能是会改变蛋白质空间结构,进而影响蛋白质正常功能的重要突变。
多态信息含量、杂合度与群体遗传多样性密切相关,其值的高低反映了群体内个体的均质度,群体遗传变异程度越大,说明该群体的遗传多样性就越丰富,选择潜力就越大,对育种改良就越有利[1]。本研究从纯合度、杂合度、有效等位基因数、多态信息含量等指标多角度对黔北麻羊群体的遗传变异程度进行分析,发现g.13665051C>T位点基因座的纯合度为0.562,杂合度为0.438,有效等位基因为1.779,多态信息含量为0.341,为中度多态位点;g.13664737G>A位点基因座的纯合度为0.531,杂合度为0.469,有效等位基因为1.883,多态信息含量为0.359,为中度多态位点,说明2个SNPs位点的群体遗传变异程度较大,遗传多样性较为丰富,具有一定的选择潜力。卡方(χ2)检验结果显示,黔北麻羊群体FABP3基因的基因型分布并未受到遗传漂变及人工选育的影响[23],符合Hardy-Weinberg平衡。
近年来,随着分子生物技术的发展,生长性状相关的分子标记已广泛应用于畜禽的育种改良中,大大提高了育种效率,缩短了育种进程[24]。本研究除在黔北麻羊FABP3基因外显子5和3′非翻译区各发现1个SNP位点外,并未发现其他突变位点,这与黄李勇等[1]在新疆巴什拜羊FABP3基因外显子2发现1个SNP位点和赵雪[25]在东北半细毛羊、小尾寒羊、杜泊和道赛特羊4个群体中发现FABP3基因外显子2存在4个突变位点,外显子3存在3个突变位点的结果存在差异,可能是由于品种、遗传基础和样本数量不同所致。本研究采集了黔北麻羊的体重、体高、体斜长、胸深、胸宽、胸围及管围等生长性状数据,并与在FABP3基因上筛选出的2个SNPs位点进行关联分析,结果显示,g.13665051C>T突变位点除了对黔北麻羊体重无显著影响外,对其体高、体斜长、胸围、胸深、胸宽和管围均有显著影响;但g.13664737G>A突变位点对黔北麻羊体重、体高、体斜长、胸围、胸深、胸宽和管围均无显著影响。由此推测,g.13665051C>T突变位点在对黔北麻羊生长性状指标上的影响程度大于g.13664737G>A突变位点。余刚等[26]研究发现周岁陕北绒山羊FABP3基因内含子3存在SNP位点,产生2种基因型:AA和AB,其AB基因型的体高、胸围显著高于AA基因型;曾鸿普等[27]研究发现五指山猪FABP3基因外显子2存在多态性,产生3种基因型:AA、AB和BB,其8月龄AB基因型五指山猪的体质量、体长和10月龄AB基因型五指山猪的胸围显著高于AA、AB基因型。上述研究结果与本试验在g.13665051C>T位点上的研究结果存在一定的相似性,证实了FABP3基因上的SNP位点与部分生长性状存在关联性,且会对部分生长性状产生影响。而柴志欣等[4]研究发现麦洼牦牛FABP3基因外显子4存在T7339C突变,但该突变对麦洼牦牛的生长性状均无显著影响,这与本试验g.13664737G>A位点的关联性分析结果一致。综合分析2种不同结果,可能是由于检测的位点、品种、采集生长性状数据的阶段及统计分析方法不同引起。在本次关联性分析中,g.13665051C>T位点中CT基因型的体高、体斜长、胸宽和管围均显著高于CC、TT基因型,CT基因型的胸深、胸围显著高于TT基因型,这表明g.13665051C>T突变位点显著影响黔北麻羊的诸多生长性状指标,初步推测FABP3基因外显子5的g.13665051C>T突变有可能作为一种遗传标记,通过对优势基因型的选择,从而加快对黔北麻羊的育种改良。
4 结论
本研究在黔北麻羊FABP3基因外显子5和3′非翻译区各检测出1个SNP位点,分别为g.13665051C>T、g.13664737G>A。其中g.13665051C>T为同义突变,产生3种基因型:CC、CT和TT;g.13664737G>A产生3种基因型:GG、AG和AA。通过遗传学分析,黔北麻羊FABP3基因存在较为丰富的遗传多态性。经关联性分析显示,g.13665051C>T突变位点对黔北麻羊体高、体斜长、胸围、胸深、胸宽和管围均有显著影响,可作为一种遗传标记用于辅助性选择。
