玉米赤霉烯酮降解酶ZHD101 的表达优化及 在玉米浆脱毒中的应用
2021-01-08谷新晰李广靖谷子林陈宝江卢海强
陈 伟,谷新晰,李广靖,徐 冉,谷子林,陈宝江,卢海强
(1.河北农业大学 食品科技学院,河北 保定 071000;2. 河北农业大学 动物科技学院,河北 保定 071000)
玉米浆是湿法工艺生产玉米淀粉的主要副产品,是1 种黏稠酸性的浆体[1]。玉米浆含有大量的氨基酸、维生素和生长因子,因此在氨基酸发酵、植物蛋白调味液和饲料等领域有重要应用价值。然而玉米极易受到玉米赤霉烯酮毒素污染[2-3],过高的玉米赤霉烯酮含量给玉米浆的应用带来了潜在的生物安全问题。玉米赤霉烯酮( Zearalenone,ZEN)是镰刀菌产生的次生代谢产物,具有较强的雌激素效应、致癌性和遗传毒性[4-6],ZEN 不仅可以降低粮食和农副产品的质量,还可以从食物链进入人体,并对人体健康造成严重损伤[7]。
大量研究者尝试物理、化学及生物法进行玉米赤霉烯酮的降解,而其中的生物酶法具有绿色、高效及反应温和等特点。目前已发现三类酶可有效降解玉米赤霉烯酮,分别为漆酶(Laccase,EC 1.10.3.2),内酯水解酶(Lactonohydrolase,EC 3.1.1)和过氧化物酶(Peroxidase,EC 1.11.1.X)[8]。内酯水解酶因降解ZEN 及ZEN 衍生物的彻底性而成为研究热点,其中ZHD101 酶因其在该类酶中的引领地位一直是研究关注的对象。2002 年,Takahashi-Ando 等首次从粉红黏帚霉 IFO7063 中获得ZEN 降解酶基因(ZHD101),随后研究人员利用大肠杆菌、裂殖酵母、酿酒酵母和水稻为宿主对ZHD101基因进行异源表达及性质研究,这为全面表征酶学性质提供了丰富的数据[9-12]。随后,ZHD101 酶的表达效率的提高及应用价值探讨逐渐成为关注重点[13]。
大肠杆菌表达系统在廉价培养基中易快速生长和易于规模化生产,大量研究结果指出外源基因密码子与表达宿主密码子偏好性的差异会降低翻译水平,甚至会阻断翻译过程。因此,通过对外源基因密码子进行优化可提高外源蛋白的表达量。除此之外,适宜的发酵参数是提高工程菌生产能力和降低生产成本的重要手段[14-16]。目前,响应面优化策略是众多优化方法中的首选。
本研究依据大肠杆菌的密码子偏好性对ZHD101基因的密码子进行同义优化,采用响应面优化策略探究工程菌最优的发酵条件,直接分析ZHD101 酶降解玉米浆中ZEN 的效果。本研究不仅可以丰富ZHD101 蛋白高效表达的策略,同时还可探究其在玉米浆ZEN 脱毒中的应用前景,为实际生产的应用提供一定指导。
1 材料与方法
1.1 材料与试剂
玉米浆样品购自河北斐默特生物科技有限公司;赤霉烯酮标准品购自上海源叶生物科技有限公司;玉米赤霉烯酮(ZEN)ELISA 试剂盒,酶免;异丙基-β-D-硫代半乳糖苷(IPTG),卡那霉素(Kan)等分子试剂购自北京索莱宝科技有限公司;菌株E. coliBL21(DE3)为本实验室保存。甲醇、乙腈为色谱纯,购自德国默克公司;其他试剂均为国产分析纯。
1.2 仪器与设备
酶标仪,Bio-RAD;SDS-PAGE 电泳设备与凝胶成像系统,北京君意东方电泳设备有限公司。立式高速低温离心机,HITACHI 公司;Biometra Tprofessional PCR 仪, 德 国Biometra 公 司;TU-1810 紫外分光光度计,北京普析通用仪器有限公司。
1.3 方法
1.3.1 玉米赤霉烯酮酶ZHD101基因的密码子优化与合成 使用数据库(http://www.kazusa.or.jp/codon/) 中的大肠杆菌相关数据将ZHD101基因(ALI16790.1)中使用频率低的密码子进行优化,同时使用RNA 折叠网络服务器和mRNA 翻译折叠优化的密码子以进行蛋白质二级结构预测手动校正。GC含量设定在40%至50%之间,没有局部峰。优化后的基因由金斯瑞生物科技有限公司合成至pET-28a 表达质粒,并转化至大肠杆菌TOP10 中保存。
1.3.2 重组菌株的构建表达及分析 将pET-28a-ZHD101 表达质粒转化至大肠杆菌BL21(DE3)感受态细胞中,涂布于含卡那抗生素的 LB 平板上,37°C 过夜培养。挑取单克隆进行菌落PCR 鉴定,阳性克隆送上海生工生物公司测序验证。将鉴定正确的重组子的菌株接种于含卡那抗生素的LB 液体培养基中,37 ℃振荡培养12 h,以1%的接种量转接于含相应抗生素的 LB 液体培养基中,37 ℃振荡培养3 h 至A600=0.8 时,加入异丙基-β-D 硫代半乳糖苷( IPTG)至终浓度1 mmol/L,在37 ℃振荡培养4 h。离心收集诱导表达后的细胞并使用超声波细胞粉碎机进行破碎。高速离心后,取适量上清进行SDS-PAGE 分析。
1.3.3 工程菌株发酵条件的优化 在单因素实验基础上,利用Minitab 17 软件设计Plackett-Burman 试验对6 个影响工程菌株产酶量的因素进行分析,筛选影响表达量的最主要影响因素。Plackett-Burman设计见表1。

表1 Plackett-Burman 试验因子与水平Table 1 Plackett Burman test factors and levels
根据Plackett-Burman 试验结果找出的3 个最主要因素设计最陡爬坡实验,并通过Plackett-Burman试验回归方程得到各因子系数,确定主要因素的变化方向及步长,得到响应面实验的中心点。在此基础上,运用Design Expert 11 设计Box-Benhnkens试验对工程菌发酵产酶条件进行3 因素3 水平的响应面分析实验,获得最佳发酵工艺条件。
1.3.4 ZHD101蛋白表达量分析 收集取菌体沉淀,液氮冻融后向其加入100 μL 裂解液(0.5 mg/mL 溶菌酶,50 mmol/L 三羟甲基氨基甲烷—盐酸缓冲液(Tris-HCl)pH 8.0,100 mmol/L 氯化钠(NaCl),冰浴 45 min 后离心 15 min,取上清制样,使用12%的聚丙烯酰胺凝胶进行SDS-PAGE(Bio-Rad),考马斯亮蓝染液对凝胶进行染色。
扫描图像并利用Quantity One 软件对每条电泳条带进行图像分析,测得电泳条带的平均灰度值乘以相应条带的面积(总灰度值)分析得到各样品中目的蛋白的表达量。
1.3.5 重组酶ZHD101在玉米浆中的应用 以0.05 mol/L Tris-HCl (pH8.0) 缓 冲 液 配 制20 μg/mL ZEN 标 准品溶液。取1 mL 毒素标准品溶液,加入1 mL 酶液使ZEN 标准品溶液终浓度为10 μg/mL,37 ℃反应2 h,使用酶免玉米赤霉烯酮(ZEN)ELISA 试剂盒MM-32862O1 测定反应体系中ZEN 的含量,由ZEN 的减少量来表征ZHD101 活性。重组酶ZHD101 酶活定义:1 个酶活单位U 为每分钟降解 1 μg ZEN 所需的酶量。
将玉米浆pH 调至8.0,在1 mL 玉米浆中加入1 mL 酶液混匀,37 ℃条件下反应1、2 和3 h,对照加入1 mL 无菌水代替酶液,进行ZEN 含量的测定。以脱毒率来评价重组酶对玉米浆中赤霉烯酮降解能力,脱毒率计算公式:脱毒率(%)=[(A1-A2) /A1]×100,其中A1为对照组玉米浆ZEN 含量,A2为酶处理组玉米浆ZEN 含量。
1.3.6 数据分析及处理 每次试验重复测定3 次,利用 Excel 和SPSS19.0 软件对测定结果进行统计分析,数据结果采用平均值±标准误差表示。
2 结果与分析
2.1 ZHD101 基因的设计和密码子优化
大量研究指出,不同物种或基因存在着同义密码子的使用偏好性,密码子偏好性显著影响外源基因在表达宿主中的转录和翻译速率。按照大肠杆菌密码子偏好性替代野生基因中的部分密码子,合成和构建ZHD101opt基因,开展重组酶的表达研究。ZHD101(ALI16 790.1)CDS 区全长为795 bp,编码1 个264AA 组成的成熟蛋白。重新设计的目标基因ZHD101opt与野生ZHD101wt序列的一致性为 81.1%。设计后的目标基因编码1 条长度为264 个氨基酸残基组成的重组蛋白,与野生ZHD101一致。
在本研究中,对ZHD101wt基因密码子、ZHD101opt基因密码子和大肠杆菌密码子使用频率进行分析,结果见表2。经分析ZHD101wt基因在同义密码子的使用偏好性上与大肠杆菌有较大差异,ZHD101wt基因序列中含有大肠杆菌稀有密码子27 个,并且ZHD101wt基因序列中有4 处出现连续稀有密码子(GGA CCC、CCC GGA、AGG AUA、CGA GGA), 稀有密码子和连续稀有密码子的存在会降低翻译效率,甚至使翻译终止。ZHD101基因在优化119个密码子后,27 个稀有密码子被相应的同义密码子替换,连续稀有密码子消失。其CAI(Codon Adaptation Index)值经OptimumGeneTM 软件在线(http://www.genscript.com.cn/ technology-support/ on-line-tools)计算为0.97;与优化前,为0.64 相比有较大改善,CAI值大于 0.8 则意味着该目标基因可以在大肠杆菌中高效表达。优化了GC含量和不利峰后,ZHD101opt基因中GC平均值由56.35%调整为60.23%,并且延长了mRNA 的半衰期,破坏了影响核糖体结合和mRNA 稳定性的茎环结构。此外,优化过程筛选并成功地修饰了顺式负作用位点。

表2 ZHD101 野生型基因密码子、ZHD101 优化型基因密码子和大肠杆菌密码子使用频率统计表Table 2 Statistics of ZHD101 wild type gene codon, ZHD101 optimized gene codon and E. coli codon usage frequency
2.2 工程菌株ZHD101opt 基因诱导表达及产物的SDS-PAGE 分析
采用热激法进行了ZHD101opt基因的大肠杆菌转化,经测序鉴定筛选出正确的工程菌E. coliBL21 (DE3){pET-28a-ZHD101opt}进行摇瓶诱导表达后,收集菌体并裂解,取上清进行 SDS-PAGE 分析表达情况,结果见图1,经SDS-PAGE 电泳分析得出ZHD101opt表观分子量约为28 ~29 kD,与理论分子量28.8 kD 相符。

图1 玉米赤霉烯酮水解酶ZHD101 的SDS-PAGE 分析Fig. 1 SDS-PAGE analysis of zearalenone hydrolase ZHD101
2.3 玉米赤霉烯酮水解酶ZHD101opt 工程菌发酵条件的优化
使 用Minitab 17 软 件 设 计Plackett-Burman 试验(N=6),每组进行3 次平行试验,以工程菌表达的ZHD101 的SDS-PAGE 总灰度值为响应值。Plackett-Burman 试验结果经分析得出诱导时间、诱导温度和诱导种龄为显著影响因子,进而设计最陡爬坡试验得出响应面试验中心点,最陡爬坡试验实验设计及分析结果见表3。
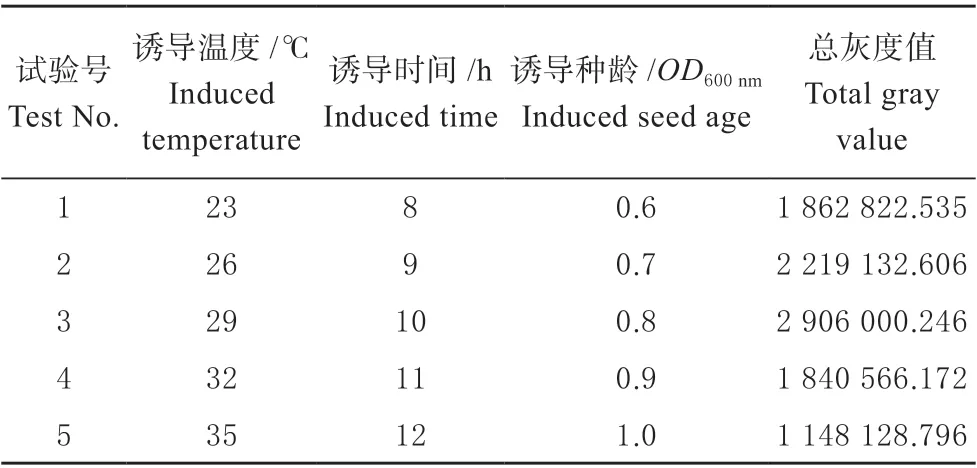
表3 最陡爬坡试验及结果Table 3 Climbing test and results
由表3 可知,试验号为3 的玉米赤霉烯酮降解酶ZHD101 表达总灰度值最大,即诱导温度29 ℃、诱导种龄0.8 和诱导时间10 h,以此为中心点进行响应面试验设计。运用Design-Expert 11 软件设计3因素3 水平响应面试验共17 个实验点的响应面分析实验。其中,5 个零点重复作为误差分析,每个试验进行3 次。响应面设计及结果见表4,回归方程方差分析见表5。
利用 Design Expert 11 软件,对中心组合实验结果进行响应面回归分析,得到工程菌的ZHD101表达量对诱导时间(A)、诱导温度(B)和诱导种龄(C) 的多元二次回归方程:总灰度值=-28 088 300+ 2 235 630A+9 56 535B+9 127 290C-12 651.562 52AB+183 675AC-35 316.667 41BC-95 994.416 80A2-13 675.468 75B2-6 002 300C2。
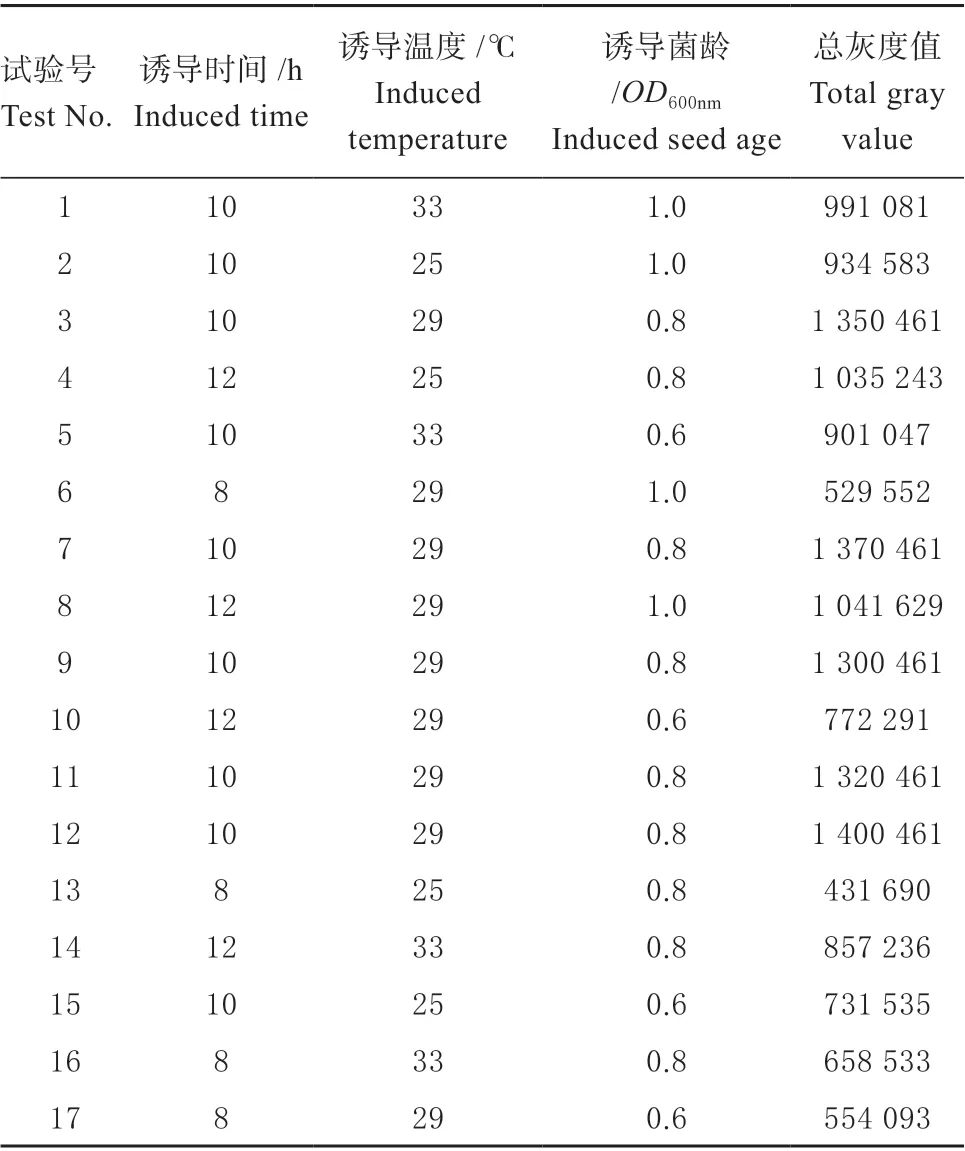
表4 响应面设计及结果Table 4 Response surface design and results
由表5 可知,本实验的回归模型P<0.000 1,说明方程拟合度较好。同时,失拟项的P=0.468 0,说明失拟不显著,残差由随机误差引起,模型选择正确;模型的决定系数R2=0.993 0,调整后调整性决定系数AdjR2=0.984 0,表明方程与实测值拟合度较高,足以反映出ZHD101的表达量与发酵条件诱导时间、诱导温度和诱导种龄的关系,说明方程模型可信度较高。
诱导时间和诱导温度对重组菌表达玉米赤霉烯酮水解酶ZHD101opt影响的交互作用最为显著,Design- Expert 11 软件预测重组菌发酵最佳条件为:诱导时间8.658 h,诱导温度30.109 ℃,诱导种龄0.998,理论总灰度值能达8 335 575.112。在验证预 测值时,将发酵最佳条件稍作调整为:诱导时间8.5 h, 诱导温度30.0 ℃,诱导种龄1.0,共进行3 次平行试验,通过SDS-PAGE 检测,以总灰度值大小作为产酶量的比较,3 次实验结果的总灰度值平均值为 9 334 666,是预测值的1.12 倍,结果表明模型预测是可用的。经与优化前的表达量进行对比发现, 表达量大幅度提升了约3 倍左右。
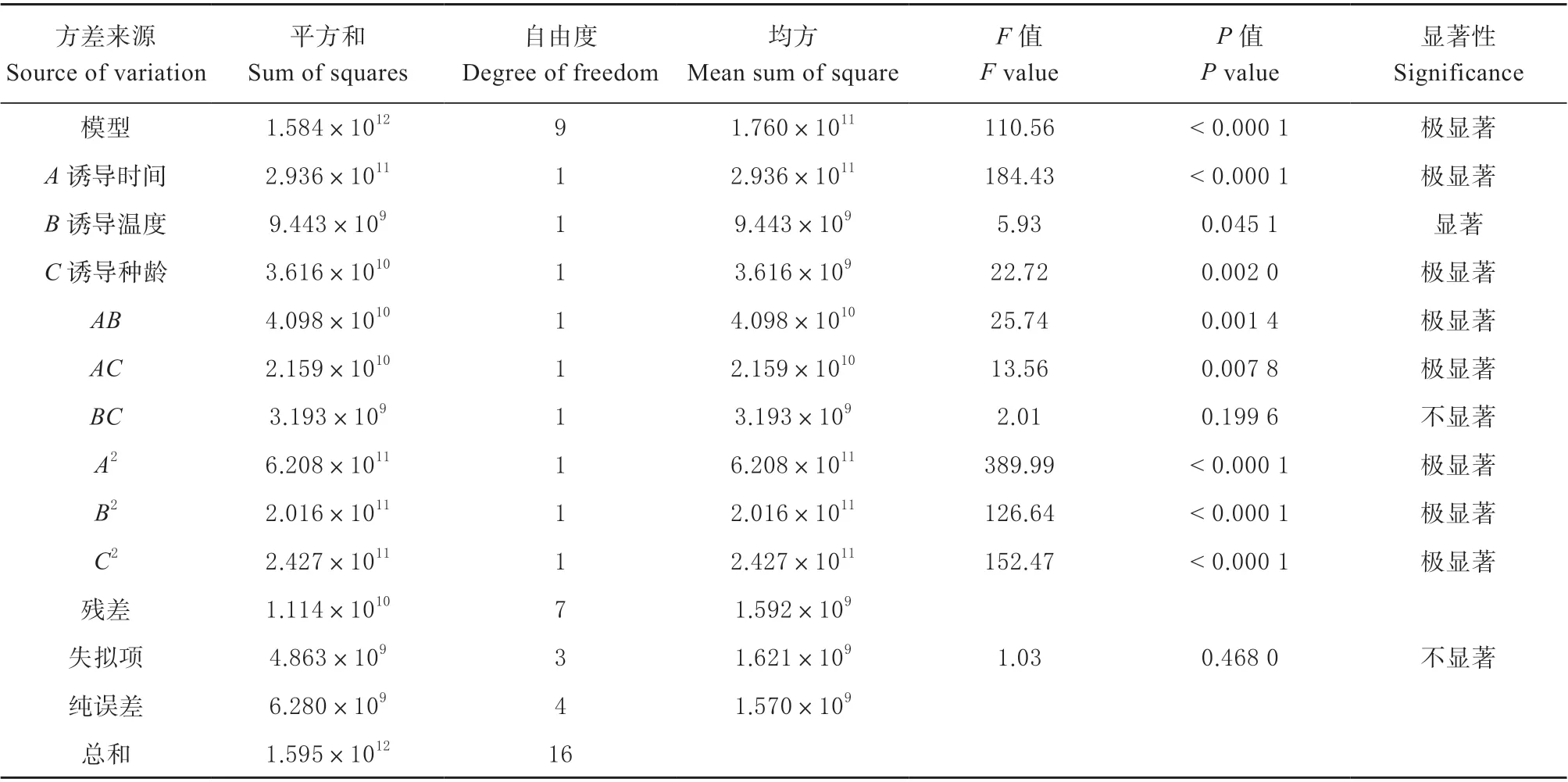
表5 回归方程方差分析 Table 5 Analysis of variance for the regression equation
2.4 玉米赤霉烯酮水解酶ZHD101 活性的测定及在玉米浆中的应用
在玉米深加工过程中,富含水溶性蛋白质的玉米浸泡液等副产物经浓缩后为玉米浆副产物。玉米浆可以作为发酵工业的营养基源;可替代饲料中的部分蛋白质原料;还可作为植物蛋白调味液而被用在食品生产等多种领域。然而由于玉米身受玉米赤霉烯酮毒素的困扰,使得相关产品的安全性受到越来越多关注。
先以ZEN 标准品为底物测定玉米赤霉烯酮水解酶ZHD101 的活性,在37 ℃ 反应2 h 可降解(5.5±1.1)μg/mL ZEN,酶 活 为0.05 U/mL。玉米浆pH 为3.5,而pH 低于4.5 可使玉米赤霉烯酮水解酶ZHD101 不可逆失活[10]。因此进行脱毒应用时,将玉米浆的pH 调至8.0 左右。如图3 所示,脱毒率随着酶解时间的延长而提高,在1、2 和3 h的脱毒率分别为(6.2±1.5)%、(13.4±2.8)%和(17.1±8.0)%,其中处理1 h 和处理2 h 的脱毒率之间差异极显著,处理1 h 和处理3 h 的脱毒率之间差异显著,而处理2 h 和处理3 h 的脱毒率之间不存在差异。该酶对玉米浆中ZEN 的降解效果不及对ZEN 标准品的降解效果,推测很可能由于酶液浓度及玉米浆复杂的成分所致,如大量的硫酸盐存在使得玉米赤霉烯酮水解酶ZHD101 活性受到抑制,相应后续试验还需继续开展。
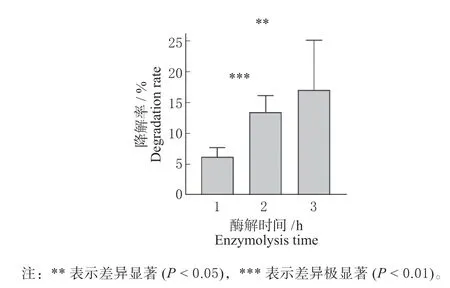
图3 不同酶解时间的玉米浆脱毒率Fig. 3 Detoxification rate of corn steep liquor with different enzymatic hydrolysis time
3 讨论
本研究参照大肠杆菌密码子偏好性对ZHD101酶原有碱基序列进行分析,发现ZHD101基因在同义密码子的使用偏好性上与大肠杆菌有较大差异,ZHD101wt基因序列中含有大肠杆菌稀有密码子27个,并且有4 处出现连续稀有密码子,稀有密码子和连续稀有密码子的存在会降低翻译效率,甚至使翻译终止。ZHD101基因在优化后,稀有密码子尽数被相应的同义密码子替换,连续稀有密码子消失,其CAI值由0.64 提高至0.97。利用宿主的密码子偏好性对目的基因进行优化以提高异源蛋白的表达量是目前得到了人们广泛认可的方法,但也有研究表明,对目的基因密码子优化后,基因表达量没有提高甚至使蛋白的含量降低[16,17]。田健等[18]利用大数据对大肠杆菌中的基因进行了统计分析发现,大肠杆菌中的高表达的基因并不是完全使用高频密码子进行表达,而是以1 种前段序列使用低频密码子,下游使用高频密码子,即外源基因采用高频密码子和低频密码子交错使用的方式来实现高效表达。除此之外,基因表达效率受多种因素的影 响[19]。因此,要在分子改造策略上实现玉米赤酶烯酮ZHD101基因高效表达,后续研究中仍需要进行系统全面的分子优化措施。
重组菌发酵条件也可显著影响重组蛋白的表达产量,如宿主外源基因的表达往往受到诱导温度、诱导时间、诱导剂浓度、pH 值、接菌量和诱导种龄等因素的影响,并且往往是这些因素的叠加。如诱导温度和诱导时间之间存在着交互作用,低温诱导可以避免重组蛋白形成包涵体,诱导温度过低则需要较长的诱导时间,而长时间的诱导可令重组蛋白降解且时间成本较高[20-22]。Garcia-Fraga 等[23]的研究发现较低的IPTG 浓度诱导可以获得最高酶活性,而极低或极高的IPTG 诱导剂浓度可以降低酶的活性。培养基pH 值主要影响宿主大肠杆菌的生长速度,而在诱导过程中,pH 值对转录效率和包涵体的形成有影响[24-25]。总之,多种条件影响着重组蛋白的表达,并且某些条件之间还存在着交互作用。响应面法可以在试验次数较低的情况下确定各独立因子对1 个响应因子的最佳影响,并且可以解释不同因子之间的相互作用,采用响应面方法在优化重要条件提高大肠杆菌中重组蛋白产量已取得较好效果[14-15]。本研究通过Plackett-Burman 设计对影响玉米赤霉烯酮降解酶ZHD101 产量的相关因素进行评价和筛选,发现诱导温度、pH、接种量是影响ZHD101 表达量的3 个显著因素,然后采用响应面分析法确定最优产酶发酵条件:诱导时间8.5 h, 诱导温度30.0 ℃,诱导种龄1.0,表达量提高了300%。
2016 年Xiang 等[13]首 次 定 义 了ZHD101 的酶活力单位,即每分钟降解1 μg/mL ZEN 所需要的酶量作为1 个酶活单位U。在此之前,玉米赤霉烯酮水解酶ZHD101 的酶活性是以对添加在培养基中的某一浓度的ZEN 标准品溶液的降解率来表征。同时Xiang 采用了密码子优化、增加外源基因拷贝数和高密度发酵表达的策略,使得ZHD101 在P.pastorisGS115 中高效表达,在甲醇诱导84 h 时,发酵上清酶活性最高可达150.1 U/mL,虽然,远远高于本研究中的大肠杆菌ZHD101 酶的表达量,但是大肠杆菌表达系统培养条件简单、生产时间短和培养成本低廉等特点,使得ZHD101 酶在大肠杆菌系统中进行表达依然具有强大优势。本课题组会在后续的试验过程中,进一步通过筛选启动子、优化基因结构、表达质粒及宿主等方面研究提高酶的表达量。
玉米赤霉烯酮毒素是1 种影响农业经济的主要毒素,严重污染谷物衍生的食品和饲料。在湿法工艺生产玉米淀粉时,大部分玉米自身毒素滞留在玉米浆中。调查研究表明,玉米及其深加工产品受ZEN 的污染较为严重,如2017 年,周建川等[26]调查发现玉米和玉米副产物中 ZEN 的检出率分别为 80.25%和88. 89%。为了提高玉米相关产品的安全性,我国颁布了相应标准,如《GB 2761—2017食品中真菌毒素限量》要求谷物及其制品中玉米赤霉烯酮含量应低于60 μg/kg,《GB13078—2017饲料卫生标准》要求饲料原料的玉米浆干粉中玉米赤霉烯酮含量不得超过1 500 μg/kg[27-28]。众多的研究只是在产酶微生物的分离及酶学性质挖掘等方面开展,而玉米赤霉烯酮降解酶表达量及应用研究相对不足。Xiang 等[13]在麦芽汁的ZEN 脱毒研究中50 μL 的ZHD101 降解酶,用12 min 可将ZEN浓度为20 μg/mL 的麦芽汁降解90%。Bi 等[29]将ZEN 降解酶ZENC (800 U) 添加到干酒糟、玉米副产品和玉米麸皮(25 g)中,ZEN 的浓度分别降低了70.9%、88.9% 和94.7%。本研究涉及的羧酸酯酶具有较强的pH 值和温度适应性,最适反应 pH 值为8.0,最适反应温度为30 ℃。酶解过程具有时间依赖性,本研究将ZEN 降解酶ZHD101(粗酶液)应用到玉米浆的ZEN 脱毒研究中,反应1、2 和 3 h 对玉米浆中的ZEN(约25 μg/mL)分别降解了(6.2±1.5)%、(13.4±2.8)%和(17.1±8.0)%,这一应用效果不及Xiang 等在麦芽汁应用效果,可能是玉米浆富含多种物质,如氨基酸、矿物质(微量元素)、有机酸、还原糖和维生素等[30],可能抑制ZEN 降解酶ZHD101 的活性。
本研究通过目的基因的密码子优化和基于Plackett-Burman 设计和响应面分析法优化发酵条件的策略,以最大程度提高大肠杆菌中玉米赤霉烯酮降解酶表达量,使酶的产量提高了3 倍左右,并且该酶在玉米浆中玉米赤霉烯酮降解应用中有着巨大前景。
